2.1. Peptide library screening
During development, hematopoietic and vascular endothelial progenitors originate from a common precursor cell, the hemangioblast, and share several phenotypic characteristics [
15]. Here we exploit this shared phenotype and use human blood outgrowth endothelial cells (HBOEC) cultured from peripheral blood mononuclear cells to enrich the phage library before panning on BM cells. This will allow for the discovery of peptides that specifically recognize molecular markers expressed both by endothelial precursors and distinct BM sub-population(s). To distinguish the cell population(s) of BM origin capable of homing to tumors, we introduced a functional feature in our screen: only phage carried by BM cells to angiogenic tumors were propagated and used for subsequent enrichment.
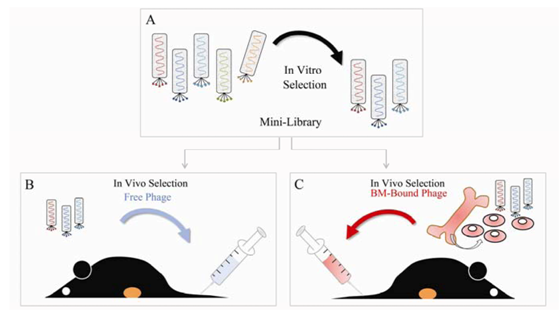
Figure 1 depicts a schematic of our selection procedures.
Figure 1.
Selection of tumor-associated phage via a combination of in vitro and in vivo biopanning protocols. (A) A random 12-mer peptide library was pre-selected in vitro on HUVEC (one round of negative selection) and on HBOEC (three rounds of positive selection) which resulted in an enriched phage pool, designated as mini-library. The mini-library was used for in vivo panning of tumor-associated phage employing two distinct selection schemes. (B) The mini-library was injected i.v. in a LLC tumor bearing mouse. Phage were allowed to circulate for 2 hours and the mouse was perfused with PBS. Then the tumor was excised, the tumor-bound phage pool was amplified in E. Coli, and used for another round of biopanning. Total of three in vivo selection cycles were conducted utilizing free phage. (C) Freshly isolated bone marrow (BM) cells were labeled with the mini-library. BM-bound phage were injected i.v. into the tail vein of a LLC tumor bearing mouse. After 2 hours of circulation the mouse was perfused with PBS. The phage were rescued from the tumor, amplified, and used to label BM cells for subsequent enrichment cycles. Three rounds of in vivo functional selection were performed using BM-bound phage.
Figure 1.
Selection of tumor-associated phage via a combination of in vitro and in vivo biopanning protocols. (A) A random 12-mer peptide library was pre-selected in vitro on HUVEC (one round of negative selection) and on HBOEC (three rounds of positive selection) which resulted in an enriched phage pool, designated as mini-library. The mini-library was used for in vivo panning of tumor-associated phage employing two distinct selection schemes. (B) The mini-library was injected i.v. in a LLC tumor bearing mouse. Phage were allowed to circulate for 2 hours and the mouse was perfused with PBS. Then the tumor was excised, the tumor-bound phage pool was amplified in E. Coli, and used for another round of biopanning. Total of three in vivo selection cycles were conducted utilizing free phage. (C) Freshly isolated bone marrow (BM) cells were labeled with the mini-library. BM-bound phage were injected i.v. into the tail vein of a LLC tumor bearing mouse. After 2 hours of circulation the mouse was perfused with PBS. The phage were rescued from the tumor, amplified, and used to label BM cells for subsequent enrichment cycles. Three rounds of in vivo functional selection were performed using BM-bound phage.
![Molecules 16 00900 g001]()
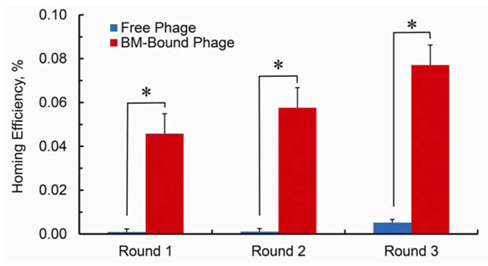
Figure 2.
Homing efficiency of the phage pool from each selection cycle, calculated as the ratio of output phage titer to input phage titer multiplied by 100. Using BM cells to deliver phage to tumors improves the efficiency of the selection procedure in the range of 50-fold. The bars show standard error of the mean (s.e.m.) from plating quadruplicates. P values were calculated by Student’s t-test and were considered statistically significant at P < 0.05.
Figure 2.
Homing efficiency of the phage pool from each selection cycle, calculated as the ratio of output phage titer to input phage titer multiplied by 100. Using BM cells to deliver phage to tumors improves the efficiency of the selection procedure in the range of 50-fold. The bars show standard error of the mean (s.e.m.) from plating quadruplicates. P values were calculated by Student’s t-test and were considered statistically significant at P < 0.05.
After one round of negative selection on human umbilical vein endothelial cells (HUVEC) to deplete the library of clones that bind to common cell surface receptors and three rounds of positive selection on HBOEC
in vitro, we generated a mini-library of enriched phage pool for further downstream biopanning (
Figure 1A). We utilized two distinct
in vivo selection protocols in parallel and compared the results: (i) The mini-library was either directly injected intravenously into a mouse bearing subcutaneously implanted LLC tumor (
Figure 1B) or (ii) A functional step was employed that consisted of administering BM-bound phage into a mouse bearing subcutaneously implanted LLC tumor (
Figure 1C). To enable the latter protocol we isolated murine BM cells from femoral and tibial bones and labeled the cell suspension with the mini-library. Free phage or BM-bound phage were allowed to circulate for 2 hours. Then the mice were perfused through the heart with PBS and the tumors were harvested. Tumor-associated phage pool from each protocol was propagated separately in
E. Coli, and used in subsequent enrichment cycles. The efficiency of each cycle was quantified as the ratio of the output phage titer to input phage titer multiplied by 100. As seen from
Figure 2 using BM cells to deliver phage to tumors improves the efficiency of the selection procedure dramatically.
To rule out library phage binding to the Matrigel matrix which is a component of the implanted tumors, we used a Matrigel plug without LLC cells as a control. Matrigel was injected symmetrically to the LLC tumor in the same animal used for the last round of biopanning. After tumors and the control plugs were excised and bound phage recovered, fifty randomly chosen phage clones resulting from each protocol both for the tumors and the control plugs (200 phage clones in total) were plaque purified and sequenced.
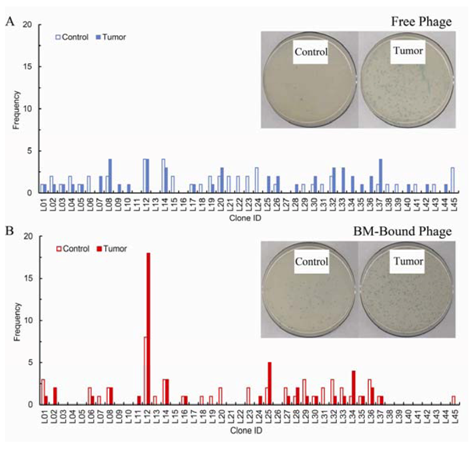
Figure 3 presents the phage inserts sequencing distribution profiles for both selection procedures for the tumors and the control plugs. The biopanning protocol with free phage does not produce a noticeable enrichment after three rounds of
in vivo selection. In contrast to what is seen with free phage, when BM cells are utilized in the panning procedure a dominant phage clone, L12, emerges. The sequencing results in
Figure 3 are in agreement with the homing efficiency findings in
Figures 2, supporting the observation that the selection procedure based on specific functional attributes,
i.e., the target is unique to cancer as these BM-derived circulating cells have the natural ability to localize to tumors, has notable efficiency and produces a phage clone that displays a preferentially enriched amino acid sequence. Regarding specificity, the dominant clone, L12, is recovered from the Matrigel plug with equal frequency to the tumor in the free phage screen, and with highest frequency (8 out of 50 sequenced clones) for the plug in the BM-bound phage screen. However, as shown in
Figure 3, we observed that the overall amount of phage homing to tumors in both screens is much higher compared to the amount of phage localizing to the Matrigel controls,
i.e., in order to obtain 50 phage clones for sequencing we processed larger amounts of control plugs compared to tumors. Thus we conclude that the selection protocols utilized here are remarkably specific as minimal background phage accumulation to the control plugs has been detected.
Figure 3.
Frequency of sequenced peptide ligands selected either by panning with free phage (A) or by panning using bone marrow-bound phage (B). After three rounds of in vivo selection, fifty phage clones homing to tumors and to Matrigel controls from each protocol were randomly chosen for sequencing. Frequency refers to the number of times each phage was isolated out of the total number of phage sequenced. In vivo selection with free phage produces no apparent enrichment of particular clones. By stark contrast, three rounds of in vivo functional selection, using BM cells to direct the phage pool to the tumor site, are sufficient to identify a dominant amino acid sequence. The inserts in both panels represent typical plates from which phage clones were selected and processed for sequencing.
Figure 3.
Frequency of sequenced peptide ligands selected either by panning with free phage (A) or by panning using bone marrow-bound phage (B). After three rounds of in vivo selection, fifty phage clones homing to tumors and to Matrigel controls from each protocol were randomly chosen for sequencing. Frequency refers to the number of times each phage was isolated out of the total number of phage sequenced. In vivo selection with free phage produces no apparent enrichment of particular clones. By stark contrast, three rounds of in vivo functional selection, using BM cells to direct the phage pool to the tumor site, are sufficient to identify a dominant amino acid sequence. The inserts in both panels represent typical plates from which phage clones were selected and processed for sequencing.
2.2. Peptide sequence analysis
The peptide sequences that appeared with the highest frequency in the sequenced phage pool are listed in
Table 1. Two of the peptides, L12 and L34, were encountered in a previous study that utilized
in vitro selection protocol for identification of HBOEC-specific peptides [
16]. Although these findings confirm our initial hypothesis of shared phenotypic expression between outgrowth endothelial precursors and circulating bone marrow derived cells, neither L12 nor L34 peptide was enriched in the HBOEC-specific phage pool (1 occurrence for each peptide in a pool of 40 sequenced peptides). Furthermore, we searched MimoDB, the newly developed repository of peptides derived by screening of phage display libraries [
17], and found that none of the peptides listed in
Table 1 has been identified to bind to other targets. We selected the phage clone with the highest frequency of occurrence for further characterization. This phage expresses the peptide sequence QFPPKLTNNSML, denoted also as QFP-peptide.
Table 1.
Peptides identified with the highest frequency from the sequenced phage pool.
Table 1.
Peptides identified with the highest frequency from the sequenced phage pool.
| Code | Peptide sequence (three letter code) | Frequency of occurrence |
| L12 | GLN-PHE-PRO-PRO-LYS-LEU-THR-ASN-ASN-SER-MET-LUE | 18 |
| L25 | SER-TRY-ASP-ILE-LEU-LYS-PRO-ASN-PRO-GLN-ARG-LEU | 5 |
| L34 | SER-HIS-GLY-LYS-PRO-PRO-SER-PHE-SER-PRO-TRY-THR | 4 |
Prompted by observations from other studies that peptides isolated by phage display are often functionally active and bind to sites of protein-protein interactions [
18,
19], we searched the non-redundant protein database (
nr) at NCBI using the BLASTp tool to find potential homologies to the QFP-peptide [
20]. BLASTp alignments with algorithm parameters adjusted to identify short and nearly exact matches, revealed a number of similarities to proteins with cell membrane or extracellular function (see
Table 2). Human immunodeficiency virus type 1 enhancer binding protein also recognized as major histocompatibility complex binding protein 1 is known to mediate immune evasion and promote viral persistence by down regulating major histocompatibility complex in peripheral blood mononuclear cells [
21]. Serine protease 55 (PRSS55) is a member of a group of membrane anchored chemotrypsin (S1)-like serine proteases that have a role in normal homeostasis as well as in pathology of diseases such as cancer. Homology with the QFP-peptide occurs at the hydrophobic part of the C-terminus of PRSS55 which provides basis for cell membrane anchoring [
22]. Cubilin (3623 aa protein) acts as a receptor for intrinsic factor-vitamin B12 (cobaltamin) complexes. Limited tissue expression of cubilin and its putative importance for cell cancer growth have motivated studies to explore cubilin as a new target for the delivery of organometallic B12-based conjugates for cancer diagnosis and treatment [
23]. Finally, we have found through analysis for conserved domains that the KLTNNS region of the integrin β2 aligns with the QFP-peptide [
24]. Βeta-2 integrins are known to play an important role in the leukocyte-endothelial interactions [
25]. Recent studies indicate that circulating tumor cells bind in increased numbers to cytokine activated endothelium in a process reminiscent with of leukocyte-endothelial adhesion [
26]. Of note, the extracellular matrix components of Matrigel in the LLC tumors may provide ligands for β2 integrin mediated interactions thus potentially causing off-specificity binding. Whether the QFP-peptide actually participates in the functional interactions identified through bioinformatics analysis and the specific molecular mechanisms involved remains to be further investigated.
Table 2.
Identified sequence homologies to the QFPPKLTNNSML peptide obtained by searching the non-redundant (nr) NCBI protein database performing local sequence alignment. The underlined amino acids correspond to exact alignment of the protein sequences with the new peptide. The numbering on both ends of the sequences indicates the region where homology occurs.
Table 2.
Identified sequence homologies to the QFPPKLTNNSML peptide obtained by searching the non-redundant (nr) NCBI protein database performing local sequence alignment. The underlined amino acids correspond to exact alignment of the protein sequences with the new peptide. The numbering on both ends of the sequences indicates the region where homology occurs.
| Protein Sequence | Protein Name | Accession |
| 233—PPKLKNSSM –241 | Human Immunodeficiency Virus
Type 1 Enhancer Binding Protein | AAI 40817.1 |
| 227—PKLTKN-ML –234 | Serine Protease 55 (PRSS55) | NP_001183949 |
| 3118—PPNVKSSNNSML – 3129 | Cubilin
(intrinsic factor cobalamin receptor) | EAN 86221.1 |
| 187—KLTNNS –192 | Integrin 2 | EAX 09383 |
2.3. Characterization of QFP-phage clone binding to LLC tumors by plaque assay
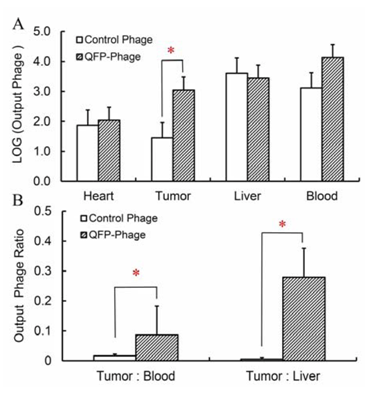
QFP-phage was grown to a high titer and used for labeling of freshly isolated BM cells. Phage-BM cell complexes were re-suspended in PBS and injected intravenously into LLC tumor bearing mice (n = 3). Experiments with control non-targeted phage were performed in parallel. After 2 hours of circulation blood was collected, the mouse perfused with PBS, and heart, tumor and liver harvested. The amount of phage in blood and each tissue was determined by plaque assay by counting the number of infectious phage particles. As shown in
Figure 4, liver and blood retained the most phage and heart retained the least at 2 hours post-injection. The amount of QFP-phage in the tumor was statistically higher than the amount of non-targeted phage. Furthermore, tumor-to-liver and tumor-to-blood ratio were significantly higher for QFP-phage compared to the control.
Figure 4.
(A) Localization of QFP- and control phage to LLC tumors, heart, liver and blood at 2 hours post-injection of BM cells labeled with phage. Statistically higher amount of QFP-phage was detected in LLC compared to a control insertless phage. (B) Tumor-to-blood and tumor-to-liver ratio for QFP- and control phage. Both ratios are statistically higher for QFP-phage compared to the control (P < 0.05).
Figure 4.
(A) Localization of QFP- and control phage to LLC tumors, heart, liver and blood at 2 hours post-injection of BM cells labeled with phage. Statistically higher amount of QFP-phage was detected in LLC compared to a control insertless phage. (B) Tumor-to-blood and tumor-to-liver ratio for QFP- and control phage. Both ratios are statistically higher for QFP-phage compared to the control (P < 0.05).
2.4. Noninvasive quantitation of in vivo peptide binding to LLC tumors by positron emission tomography
Next, we explored the ability of the QFPPKLTNNSML peptide to deliver molecular cargo to sites of ongoing angiogenesis by noninvasive positron emission tomography (PET) imaging. For this purpose we developed a labeling platform employing the phage that displays the QFPPKLTNNSML peptide as a molecularly targeted imaging agent. Using phage as imaging probes and biological nanoparticles in targeting tumors offers an immediate advantage: phage can be covalently attached to numerous labels while simultaneously expressing multiple copies of the tumor-avid peptide [
27]. In this study we labeled QFP-phage with
64Cu radionuclide via the macrocyclic chelator 1,4,7,10-tetraazacyclododecane-1,4,7,10-tetra acetic acid (DOTA). First, we functionalized the phage particle conducting a coupling chemical reaction between amino groups exposed on the phage surface and the bifunctional p-SCN-Bn-DOTA derivative as illustrated in
Figure 5. In a second step we performed a labeling reaction to attach the
64Cu
2+ radiometal to the DOTA-phage-QFP construct.
64Cu radiolabeled insertless phage conjugates and
64Cu-DOTA complexes were prepared to serve as controls. The chemical modification of the phage surface did not affect cellular binding or phage infectivity (results not shown). Thus phage retained target affinity and biological activity on labeling.
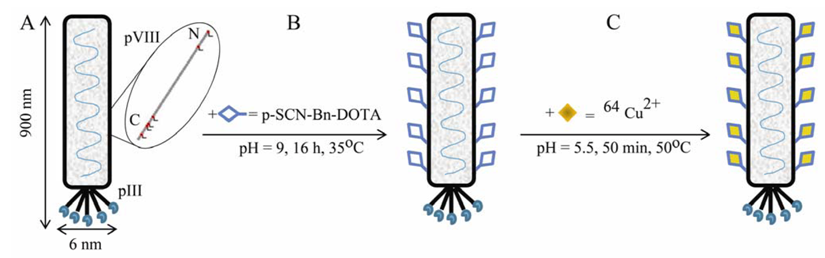
Figure 5.
Design and synthesis of M13 phage-based molecularly targeted imaging platform for noninvasive assessment and visualization of
in vivo peptide binding by positron emission tomography (PET). (A) M13 bacteriophage is a long filamentous particle approx. 6 nm in diameter and 900 nm in length. The viral genome is encapsulated in approx. 2700 copies of the major coat protein pVIII. The minor coat protein pIII that caps the particle is engineered to display five copies of a tumor-avid QFPPKLTNNSML peptide. Five lysine groups (Lys 8, 40, 43, 44, 48) and one N-terminal amino group (Ala 1) on each pVIII subunit are available for chemical modification [
28]. (B) Bifunctional p-SCN-Bn-DOTA derivative is added to a 1×10
11 pfu phage in conjugation buffer at pH = 9. The chemical reaction for covalent attachment of DOTA to the phage surface is carried out for 16 hours at 35°C. (C) The
64Cu
2+ radiometal is attached through coordinative binding to the DOTA-phage construct for 50 min at 50°C.
Figure 5.
Design and synthesis of M13 phage-based molecularly targeted imaging platform for noninvasive assessment and visualization of
in vivo peptide binding by positron emission tomography (PET). (A) M13 bacteriophage is a long filamentous particle approx. 6 nm in diameter and 900 nm in length. The viral genome is encapsulated in approx. 2700 copies of the major coat protein pVIII. The minor coat protein pIII that caps the particle is engineered to display five copies of a tumor-avid QFPPKLTNNSML peptide. Five lysine groups (Lys 8, 40, 43, 44, 48) and one N-terminal amino group (Ala 1) on each pVIII subunit are available for chemical modification [
28]. (B) Bifunctional p-SCN-Bn-DOTA derivative is added to a 1×10
11 pfu phage in conjugation buffer at pH = 9. The chemical reaction for covalent attachment of DOTA to the phage surface is carried out for 16 hours at 35°C. (C) The
64Cu
2+ radiometal is attached through coordinative binding to the DOTA-phage construct for 50 min at 50°C.
We utilized PET imaging to quantitate noninvasively the ability of the
64Cu labeled QFP-phage to bind
in vivo to its target in angiogenic highly vascularized subcutaneously implanted LLC tumors (n = 3). The labeled phage were injected i.v. into a tumor bearing mouse and the localization of
64Cu-DOTA-phage -QFP radiotracer visualized. The distribution of two control radiotracers,
64Cu-DOTA and
64Cu-DOTA- phage with no peptide insert, was examined in parallel. Representative decay corrected coronal PET images at 18 hours post-injection are shown in
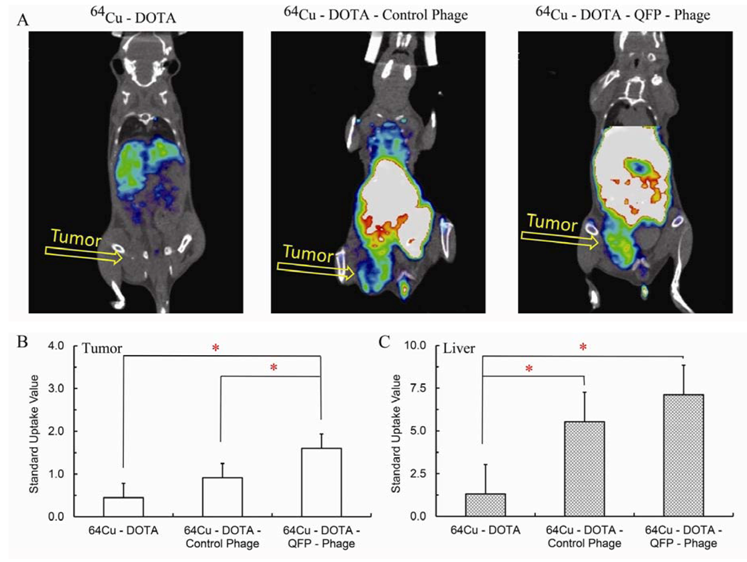
Figure 6A.
64Cu-DOTA-phage -QFP was able to bind its target
in vivo thus producing an excellent tumor uptake and contrast in the tumor tissue while the control phage showed little to no accumulation in the tumor.
64Cu-DOTA complex revealed a completely different pattern of
in vivo distribution confirming that the signal from the
64Cu-DOTA-phage -QFP is due to the
64Cu-DOTA covalently bound in a stable manner to the QFP-phage vector.
Figure 6B and C quantify the standard uptake values (SUV) for the tumor and liver, respectively.
64Cu-DOTA-phage –QFP has a statistically higher standard tumor uptake value compared to both controls.
Figure 6.
(A) Representative coronal microPET images of mice bearing LLC tumors. Images were acquired 18 hours post-injection of 800 μCi of 64Cu-DOTA, 64Cu-DOTA-control non-targeted phage, or 64Cu-DOTA-QFP-targeted phage. (B) Standard uptake values calculated from the coronal PET images in tumor (mean ± s.e.m., n = 3). (C) Standard uptake values calculated from the coronal PET images in liver (mean ± s.e.m., n=3). P values were determined by Student’s t-test and were considered statistically significant at P < 0.05.
Figure 6.
(A) Representative coronal microPET images of mice bearing LLC tumors. Images were acquired 18 hours post-injection of 800 μCi of 64Cu-DOTA, 64Cu-DOTA-control non-targeted phage, or 64Cu-DOTA-QFP-targeted phage. (B) Standard uptake values calculated from the coronal PET images in tumor (mean ± s.e.m., n = 3). (C) Standard uptake values calculated from the coronal PET images in liver (mean ± s.e.m., n=3). P values were determined by Student’s t-test and were considered statistically significant at P < 0.05.
As seen in
Figure 6 there is a capture of radiolabeled phage by the liver. These observations are consistent with the findings from the phage distribution experiments presented in
Figure 4. Both QFP-phage and the insertless control phage are taken up and catabolized by the liver. The fact that liver localization is independent of the presence of a peptide on the surface of the phage clearly indicates that liver uptake is a property of the phage, and not the peptide carried by the phage. Taken together our imaging results reveal that the QFPPKLTNNSML peptide identified by
in vivo functional selection from a phage display peptide library can deliver molecular cargo to angiogenic tumors.
Possible limitations may be associated with the synthetic approach employed in this study, i.e., the chemical modification of primary amino groups exposed on the surface of the phage particle. The isothiocyanate of the DOTA reagent can presumably form a thiourea with the side chain primary amine of the lysine residue of the QFP-peptide thus potentially reducing the effective binding affinity of the targeting moiety. This in turn may affect the biodistribution profile of the radioconjugate. Detailed MALDI-TOF MS analysis is currently underway to further characterize the reaction products and identify modification sites. Alternative bioconjugation methods such as covalent attachment of DOTA to the carboxylic reactive groups of the aspartic and glutamic acid residues or to the phenol groups of the tyrosine on the phage surface are warranted in future studies.
The ability to target vasculature for imaging or drug delivery purposes has been an elusive goal. Peptide sequences harnessing endogenous cellular machinery to accomplish this goal represent a new targeting approach that is likely to escape immune surveillance. The studies reported here provide proof-of-principle for this approach. Subsequent refinements using modified peptide sequences coupled to imaging agents and/or biologically active molecules may have clinical utility in a variety of physiologic and pathologic processes where neovascular growth is a critical component.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}


