Prebiotic Potential of Agave angustifolia Haw Fructans with Different Degrees of Polymerization

Abstract
:1. Introduction
2. Results and Discussion
2.1. Agave Fructan Extraction and Purification

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification | Fractionation | ||||||
|---|---|---|---|---|---|---|---|
| Crude Extract | Deproteinized UF 10 KDa | Ion-exchange Chromatography | AFF1 > 3 KDa | AFF2 3 a 1 KDa | AFF3 < 1 KDa | ||
| Conductivity μs | 1403 | 1365 | 18.5 | 22 | 22 | 21 | |
| pH | 4.9 | 5.02 | 6.09 | 6.2 | 6.1 | 6.2 | |
| Dissolved solids °Brix | 15.1 | 15 | 11.4 | 19 | 21 | 22 | |
| Protein % | ~0.2 | - | - | - | - | - | |
| Vol L | 17 | 16 | 19.5 | 5.13 | 3.23 | 2.12 | |
| Total solids g | 2567 | 2400 | 2223 | 974.7 | 678.3 | 466.4 | |
| % | 46 | 32 | 22 | ||||
2.2. Agave Fructans Fractionation
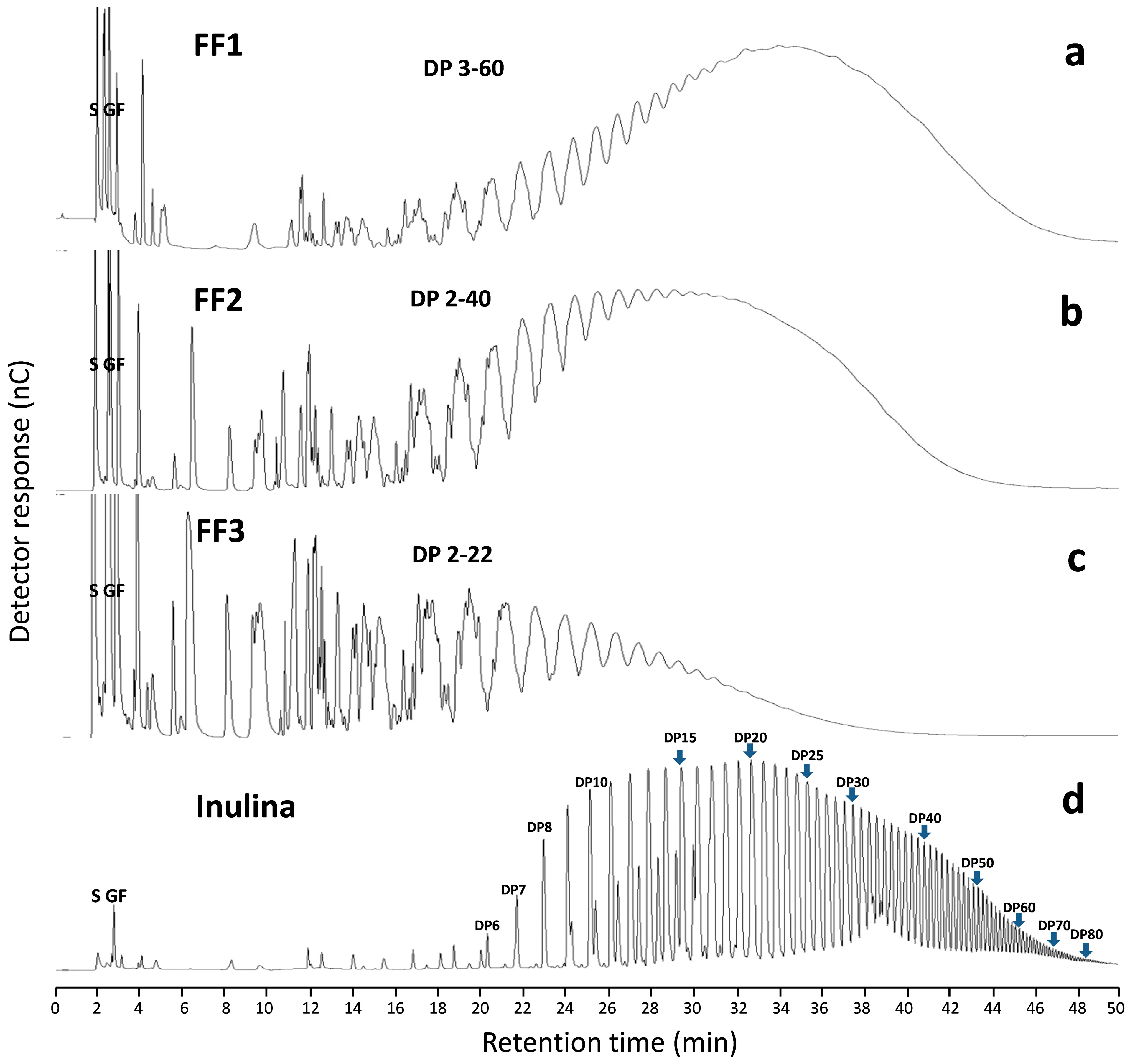
2.3. AFF Fructan Profile (DP)

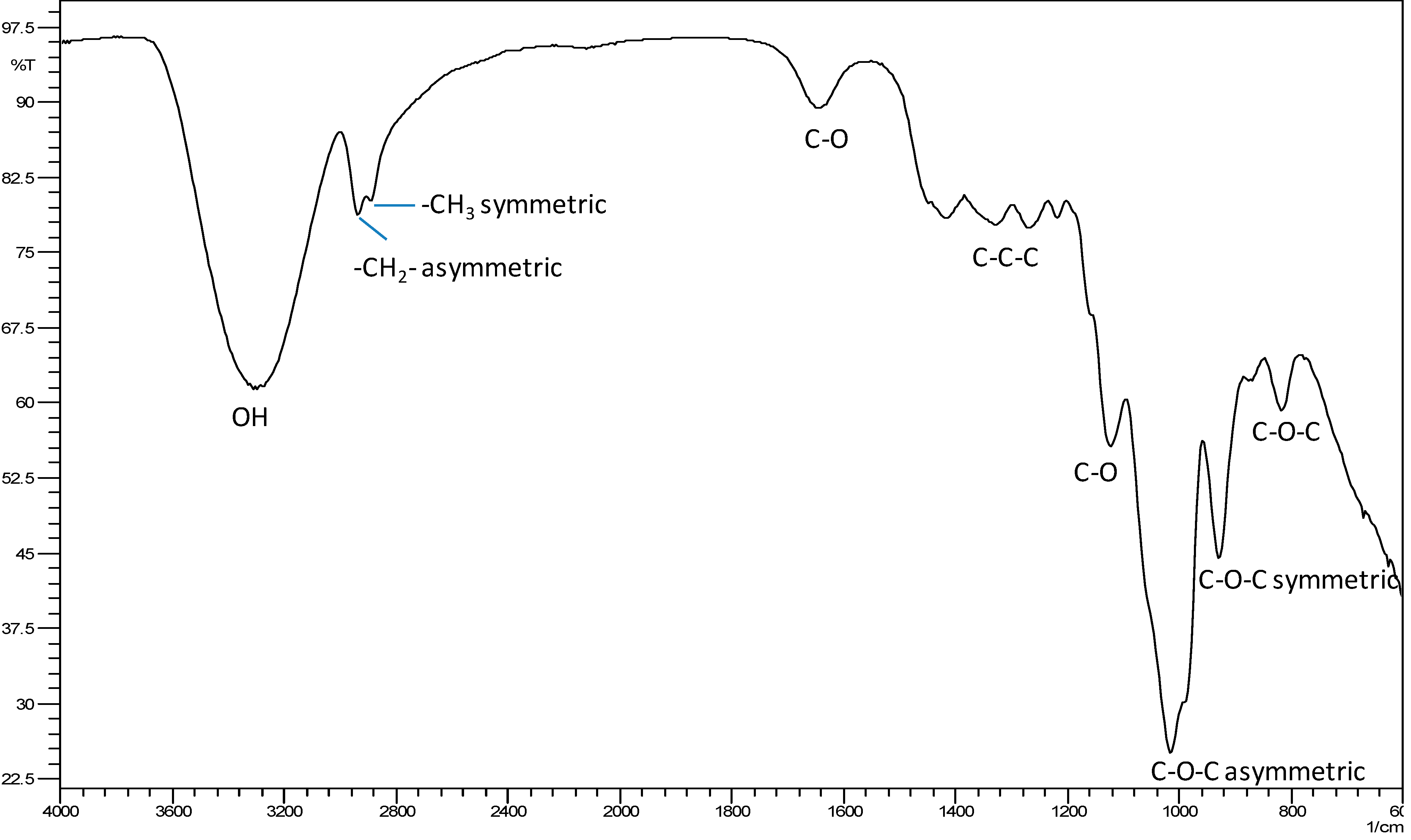
2.4. Molecular Recognition of Fructans Using Fourier Transform Infra-Red (FTIR) Spectroscopy


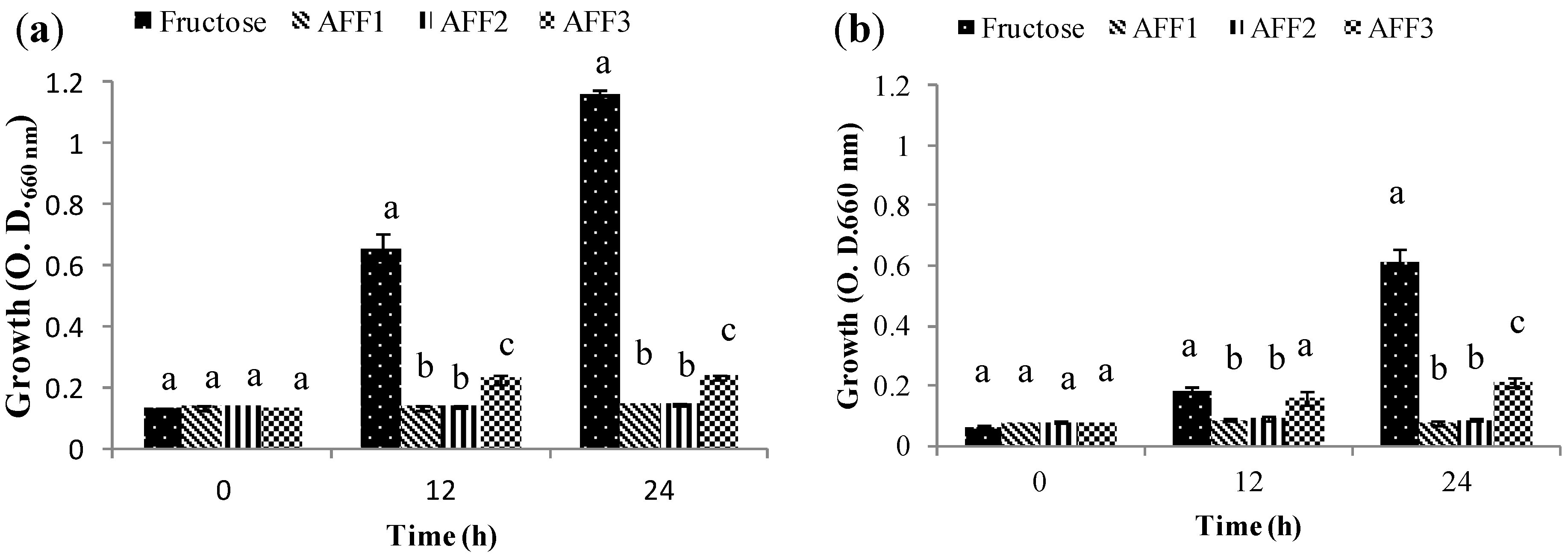
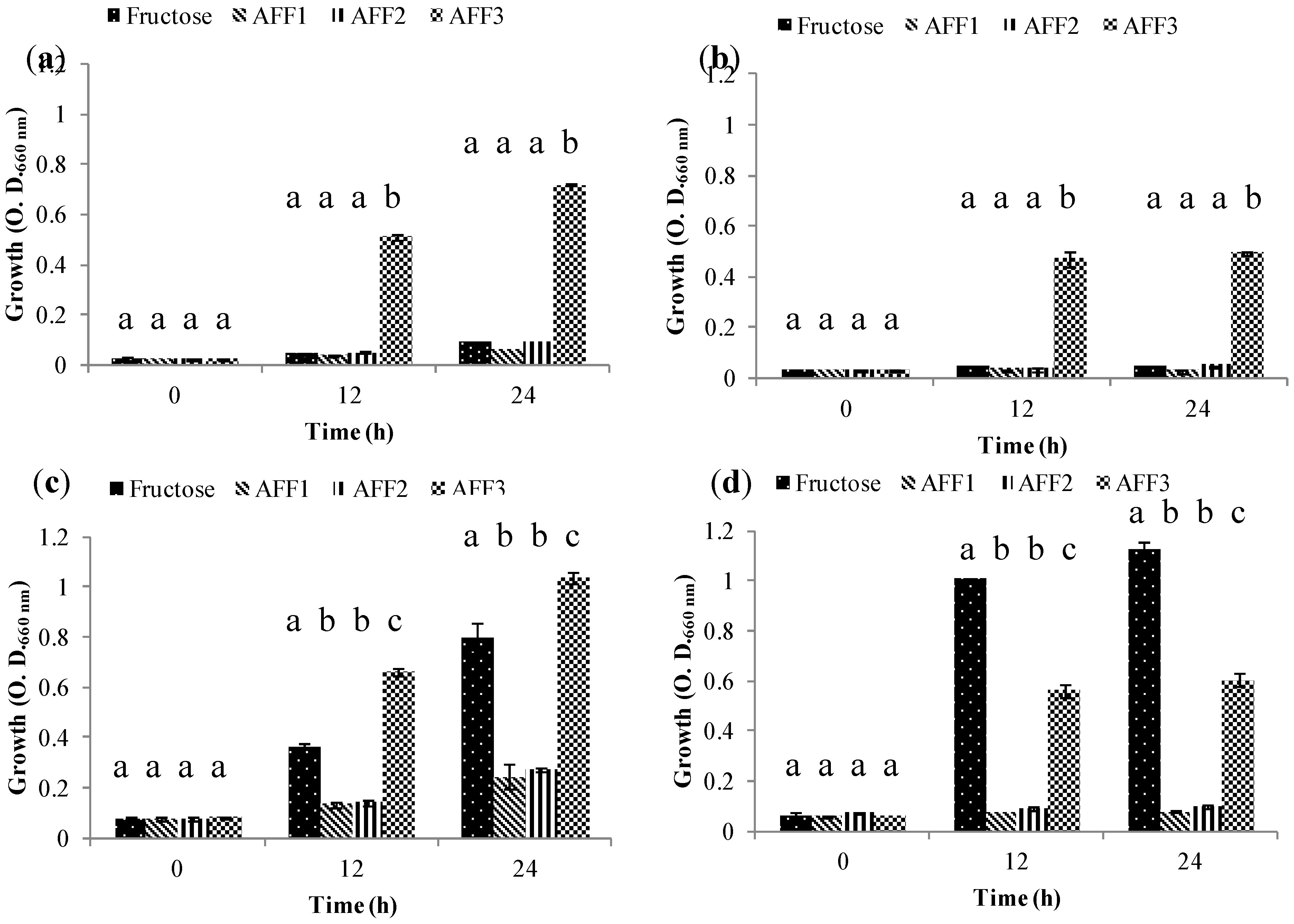
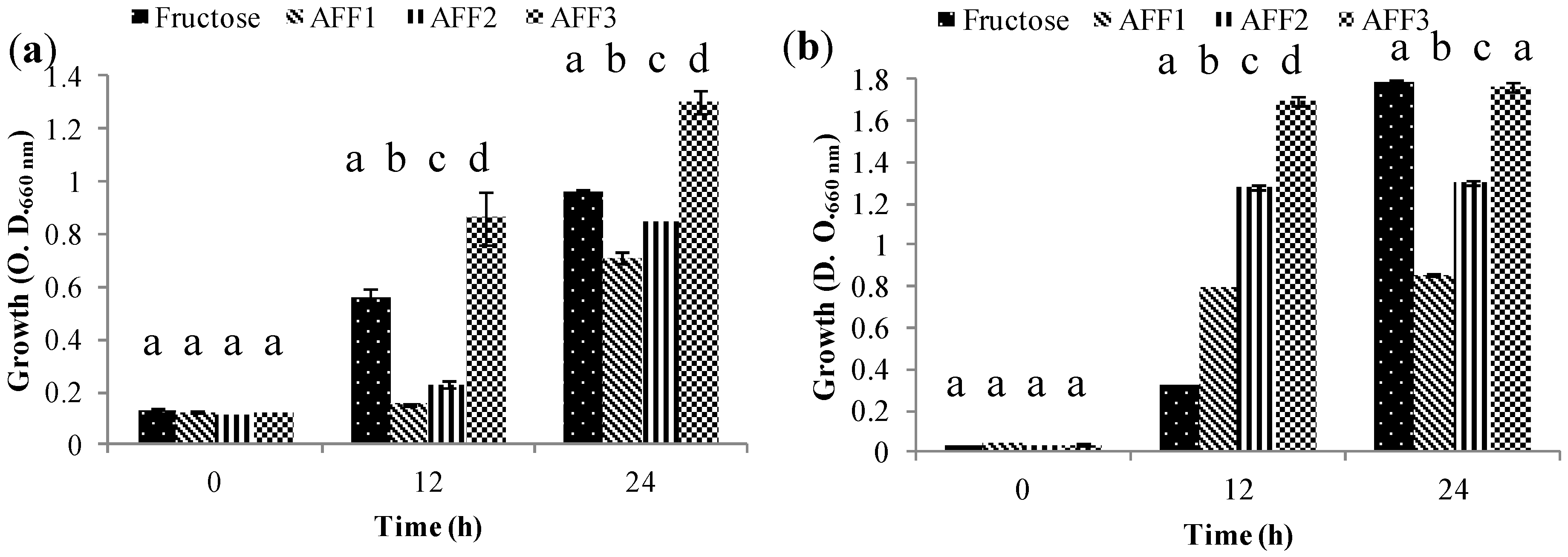
2.5. Prebiotic Effect “In Vitro”
| Strain | G1 | G2 | G3 | G4 |
|---|---|---|---|---|
| B. adolecentis ATCC 15703 | X | |||
| B. animalis ATCC 25527 | X | |||
| B. bifidum ATCC 29521 | X | |||
| B. breve ATCC 15700 | X | |||
| B. infantis ATCC 17930 | X | |||
| B. longum ATCC 15707 | X | |||
| B. lactis DSM 10140 | X | |||
| L. casei subsp. rhamnosus ATCC 9595 | X | |||
| L. paracasei subsp. paracasei | X | |||
| L. plantarum 299V | X |




3. Experimental Section
3.1. Chemical Reagents
3.2. Extraction of Agave angustifolia Haw Fructans
3.2.1. Agave Stems
3.2.2. Extraction
3.3. Purification of Agave angustifolia Haw fructans
3.4. Agave Fructans Fractionation
Thin Layer Chromatography (TLC)
3.5. AFF Fructan Profile (DP)
| Time (min) | Flow (mL/min) | Step |
|---|---|---|
| 0–5 | 0.42 | 100% A |
| 5–44 | 0.42 | Gradient to 50% A/50% B |
| 44–50 | 0.42 | Isocratic at 50% A/50% B |
| 50–53 | 0.35 | Gradient 50% C/50% D |
| 53–60 | 0.35 | Isocratic at 50% C/50% D |
| 60–70 | 0.35 | Gradient to 100% A |
| 70–80 | 0.35 | 100% A |
| 80–90 | 0.42 | 100% A |
3.6. Structure of Fructan from A. angustifolia Haw
3.7. Prebiotic Effect “In Vitro”
3.7.1. Media
3.7.2. Microorganisms
3.7.3. Fermentation from Fructans of Agave angustifolia Haw
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- García, A. Los agaves de México. Ciencias 2007, 87, 14–23. [Google Scholar]
- Trueba, L.A.C. Los destilados de agave en méxico y su denominación de origen. Ciencias 2007, 40–49. [Google Scholar]
- Wang, N.; Nobel, P.S. Phloem transport of fructans in the crassulacean acid metabolism species agave deserti. Plant Physiol. 1998, 116, 709–714. [Google Scholar] [CrossRef]
- Vijn, I.; Smeekens, S. Fructan: More than a reserve carbohydrate? Plant Physiol. 1999, 120, 351–360. [Google Scholar] [CrossRef]
- Ritsema, T.; Smeekens, S. Fructans: Beneficial for plants and humans. Curr. Opin. Plant Biol. 2003, 6, 223–230. [Google Scholar] [CrossRef]
- Waleckx, E.; Gschaedler, A.; Colonna-Ceccaldi, B.; Monsan, P. Hydrolysis of fructans from Agave tequilana weber var. Azul during the cooking step in a traditional tequila elaboration process. Food Chem. 2008, 108, 40–48. [Google Scholar]
- Mancilla-Margalli, N.A.; López, M.G. Water-soluble carbohydrates and fructan structure patterns from agave and dasylirion species. J. Agric. Food Chem. 2006, 54, 7832–7839. [Google Scholar]
- Mellado-Mojica, E.; López, M.G. Fructan metabolism in a. Tequilana weber blue variety along its developmental cycle in the field. J. Agric. Food Chem. 2012, 60, 11704–11713. [Google Scholar]
- Kolida, S.; Gibson, G.R. Prebiotic capacity of inulin-type fructans. J. Nutr. 2007, 137, 2503S–2506S. [Google Scholar]
- Biedrzycka, E.; Bielecka, M. Prebiotic effectiveness of fructans of different degrees of polymerization. Trends Food Sci. Tech. 2004, 15, 170–175. [Google Scholar] [CrossRef]
- Roberfroid, M. Prebiotics: The concept revisited. J. Nutr. 2007, 137, 830S–837S. [Google Scholar]
- Saulnier, D.M.A.; Spinler, J.K.; Gibson, G.R.; Versalovic, J. Mechanisms of probiosis and prebiosis: Considerations for enhanced functional foods. Curr. Opin. Biotech. 2009, 20, 135–141. [Google Scholar] [CrossRef]
- Van de Wiele, T.; Boon, N.; Possemiers, S.; Jacobs, H.; Verstraete, W. Inulin-type fructans of longer degree of polymerization exert more pronounced in vitro prebiotic effects. J. Appl. Microbiol. 2007, 102, 452–460. [Google Scholar]
- Pompei, A.; Cordisco, L.; Raimondi, S.; Amaretti, A.; Pagnoni, U.M.; Matteuzzi, D.; Rossi, M. In vitro comparison of the prebiotic effects of two inulin-type fructans. Anaerobe 2008, 14, 280–286. [Google Scholar] [CrossRef]
- Márquez-Aguirre, A.L.; Camacho-Ruiz, R.M.; Arriaga-Alba, M.; Padilla-Camberos, E.; Kirchmayr, M.R.; Blasco, J.L.; González-Avila, M. Effects of agave tequilana fructans with different degree of polymerization profiles on the body weight, blood lipids and count of fecal lactobacilli/bifidobacteria in obese mice. Food Function 2013, 4, 1237–1244. [Google Scholar]
- López-Molina, D.; Navarro-Martínez, M.D.; Rojas-Melgarejo, F.; Hiner, A.N.P.; Chazarra, S.; Rodríguez-López, J.N. Molecular properties and prebiotic effect of inulin obtained from artichoke (Cynara scolymus L.). Phytochemistry 2005, 66, 1476–1484. [Google Scholar]
- Yang, Z.; Hu, J.; Zhao, M. Isolation and quantitative determination of inulin-type oligosaccharides in roots of morinda officinalis. Carbohydr. Polym. 2011, 83, 1997–2004. [Google Scholar] [CrossRef]
- Ávila-Fernández, Á; Galicia-Lagunas, N.; Rodríguez-Alegría, M.E.; Olvera, C.; López-Munguía, A. Production of functional oligosaccharides through limited acid hydrolysis of agave fructans. Food Chem. 2011, 129, 380–386. [Google Scholar]
- López, M.G.; Urıas-Silvas, J. Agave Fructans as Prebiotics. In Advances in Fructooligosaccharides Research; Shiami, N., Benkeblia, N., Ondera, S., Eds.; Research Signpost: Kerala, India, 2007. [Google Scholar]
- Jenkins, C.L.D.; Lewis, D.; Bushell, R.; Belobrajdic, D.P.; Bird, A.R. Chain length of cereal fructans isolated from wheat stem and barley grain modulates in vitro fermentation. J. Cereal Sci. 2011, 53, 188–191. [Google Scholar] [CrossRef]
- Pavis, N.; Chatterton, N.; Harrison, P.; Baumgartner, S.; Praznik, W.; Boucaud, J.; Prud’Homme, M. Structure of fructans in roots and leaf tissues of lolium perenne. New Phytol. 2001, 150, 83–95. [Google Scholar] [CrossRef]
- Ravenscroft, N.; Cescutti, P.; Hearshaw, M.A.; Ramsout, R.; Rizzo, R.; Timme, E.M. Structural analysis of fructans from agave americana grown in south africa for spirit production. J. Agric. Food Chem. 2009, 57, 3995–4003. [Google Scholar] [CrossRef]
- Grube, M.; Bekers, M.; Upite, D.; Kaminska, E. Infrared spectra of some fructans. J. Spectroscopy 2002, 16, 289–296. [Google Scholar] [CrossRef]
- Fariña, J.; Viñarta, S.; Cattaneo, M.; Figueroa, L. Structural stability of sclerotium rolfsii atcc 201126 β-glucan with fermentation time: A chemical, infrared spectroscopic and enzymatic approach. J. Appl. Microbiol. 2009, 106, 221–232. [Google Scholar]
- Nikonenko, N.; Buslov, D.; Sushko, N.; Zhbankov, R. Spectroscopic manifestation of stretching vibrations of glycosidic linkage in polysaccharides. J. Mol. Struct. 2005, 752, 20–24. [Google Scholar] [CrossRef]
- Wang, J.; Yuan, X.; Sun, B.; Cao, Y.; Tian, Y.; Wang, C. On-line separation and structural characterisation of feruloylated oligosaccharides from wheat bran using hplc-esi-msn. Food Chem. 2009, 115, 1529–1541. [Google Scholar] [CrossRef]
- Azmi, A.F.M.N.; Mustafa, S.; Hashim, D.M.; Manap, Y.A. Prebiotic activity of polysaccharides extracted from gigantochloa levis (buluh beting) shoots. Molecules 2012, 17, 1635–1651. [Google Scholar] [CrossRef]
- Wu, X.Y.; Lee, P.I. Preparation and characterization of inulin ester microspheres as drug carriers. J. Appl. Polym. Sci. 2000, 77, 833–840. [Google Scholar] [CrossRef]
- Bolado, M.; Acedo, F. Sugar catabolism in bifidobacteria. Salus 2006, 7. [Google Scholar]
- Kaplan, H.; Hutkins, R.W. Metabolism of fructooligosaccharides by lactobacillus paracasei 1195. Appl. Environ. Microbiol. 2003, 69, 2217–2222. [Google Scholar] [CrossRef] [PubMed]
- Falony, G.; Lazidou, K.; Verschaeren, A.; Weckx, S.; Maes, D.; De Vuyst, L. In vitro kinetic analysis of fermentation of prebiotic inulin-type fructans by bifidobacterium species reveals four different phenotypes. Appl. Environ. Microbiol. 2009, 75, 454–461. [Google Scholar] [CrossRef]
- Van der Meulen, R.; Makras, L.; Verbrugghe, K.; Adriany, T.; De Vuyst, L. In vitro kinetic analysis of oligofructose consumption by bacteroides and bifidobacterium spp. indicates different degradation mechanisms. Appl. Environ. Microbiol. 2006, 72, 1006–1012. [Google Scholar]
- Amaretti, A.; Bernardi, T.; Tamburini, E.; Zanoni, S.; Lomma, M.; Matteuzzi, D.; Rossi, M. Kinetics and metabolism of bifidobacterium adolescentis mb 239 growing on glucose, galactose, lactose, and galactooligosaccharides. Appl. Environ. Microbiol. 2007, 73, 3637–3644. [Google Scholar] [CrossRef] [Green Version]
- Van der Meulen, R.; Avonts, L.; De Vuyst, L. Short fractions of oligofructose are preferentially metabolized by bifidobacterium animalis dn-173 010. Appl. Environ. Microbiol. 2004, 70, 1923–1930. [Google Scholar]
- Makras, L.; van Acker, G.; De Vuyst, L. Lactobacillus paracasei subsp. Paracasei 8700:2 degrades inulin-type fructans exhibiting different degrees of polymerization. Appl. Environ. Microbiol. 2005, 71, 6531–6537. [Google Scholar]
- Lingyun, W.; Jianhua, W.; Xiaodong, Z.; Da, T.; Yalin, Y.; Chenggang, C.; Tianhua, F.; Fan, Z. Studies on the extracting technical conditions of inulin from jerusalem artichoke tubers. J. Food Engin. 2007, 79, 1087–1093. [Google Scholar] [CrossRef]
- Hincha, D.K.; Livingston Iii, D.P.; Premakumar, R.; Zuther, E.; Obel, N.; Cacela, C.; Heyer, A.G. Fructans from oat and rye: Composition and effects on membrane stability during drying. Biochimica et Biophysica Acta (BBA)–Biomembranes 2007, 1768, 1611–1619. [Google Scholar]
- Yamazaki, H.; Matsumoto, K. Purification of jerusalem artichoke fructans and their utilisation by bifidobacteria. J. Sci. Food Agric. 1994, 64, 461–465. [Google Scholar] [CrossRef]
- Michel-Cuello, C.; Ortiz-Cerda, I.; Moreno-Vilet, L.; Grajales-Lagunes, A.; Moscosa-Santillan, M.; Bonnin, J.; González-Chávez, M.M; Ruiz-Cabrera, M. Study of enzymatic hydrolysis of fructans from agave salmiana characterization and kinetic assessment. Sci. World J. 2012, 2012, 10. [Google Scholar]
- Chandrashekar, P.M.; Prashanth, K.V.H.; Venkatesh, Y.P. Isolation, structural elucidation and immunomodulatory activity of fructans from aged garlic extract. Phytochemistry 2011, 72, 255–264. [Google Scholar] [CrossRef]
- Corradini, C.; Cavazza, A.; Bignardi, C. High-performance anion-exchange chromatography coupled with pulsed electrochemical detection as a powerful tool to evaluate carbohydrates of food interest: Principles and applications. Inter. J. Carbohydr. Chem. 2012, 2012. [Google Scholar]
- Kagan, I.A.; Kirch, B.H.; Thatcher, C.D.; Strickland, J.R.; Teutsch, C.D.; Elvinger, F.; Pleasant, R.S. Seasonal and diurnal variation in simple sugar and fructan composition of orchardgrass pasture and hay in the piedmont region of the united states. J. Equine Vet. Sci. 2011, 31, 488–497. [Google Scholar] [CrossRef]
- Willems, J.L.; Low, N.H. Major carbohydrate, polyol, and oligosaccharide profiles of agave syrup. Application of this data to authenticity analysis. J. Agric. Food Chem. 2012, 60, 8745–8754. [Google Scholar]
- González, R.; Klaassens, E.S.; Malinen, E.; De Vos, W.M.; Vaughan, E.E. Differential transcriptional response of bifidobacterium longum to human milk, formula milk, and galactooligosaccharide. Appl. Environ. Microbiol. 2008, 74, 4686–4694. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Velázquez-Martínez, J.R.; González-Cervantes, R.M.; Hernández-Gallegos, M.A.; Mendiola, R.C.; Aparicio, A.R.J.; Ocampo, M.L.A. Prebiotic Potential of Agave angustifolia Haw Fructans with Different Degrees of Polymerization. Molecules 2014, 19, 12660-12675. https://doi.org/10.3390/molecules190812660
Velázquez-Martínez JR, González-Cervantes RM, Hernández-Gallegos MA, Mendiola RC, Aparicio ARJ, Ocampo MLA. Prebiotic Potential of Agave angustifolia Haw Fructans with Different Degrees of Polymerization. Molecules. 2014; 19(8):12660-12675. https://doi.org/10.3390/molecules190812660
Chicago/Turabian StyleVelázquez-Martínez, José Rodolfo, Rina M. González-Cervantes, Minerva Aurora Hernández-Gallegos, Roberto Campos Mendiola, Antonio R. Jiménez Aparicio, and Martha L. Arenas Ocampo. 2014. "Prebiotic Potential of Agave angustifolia Haw Fructans with Different Degrees of Polymerization" Molecules 19, no. 8: 12660-12675. https://doi.org/10.3390/molecules190812660
APA StyleVelázquez-Martínez, J. R., González-Cervantes, R. M., Hernández-Gallegos, M. A., Mendiola, R. C., Aparicio, A. R. J., & Ocampo, M. L. A. (2014). Prebiotic Potential of Agave angustifolia Haw Fructans with Different Degrees of Polymerization. Molecules, 19(8), 12660-12675. https://doi.org/10.3390/molecules190812660