
Extract from Maize (Zea mays L.): Antibacterial Activity of DIMBOA and Its Derivatives against Ralstonia solanacearum


Abstract
:1. Introduction
2. Results
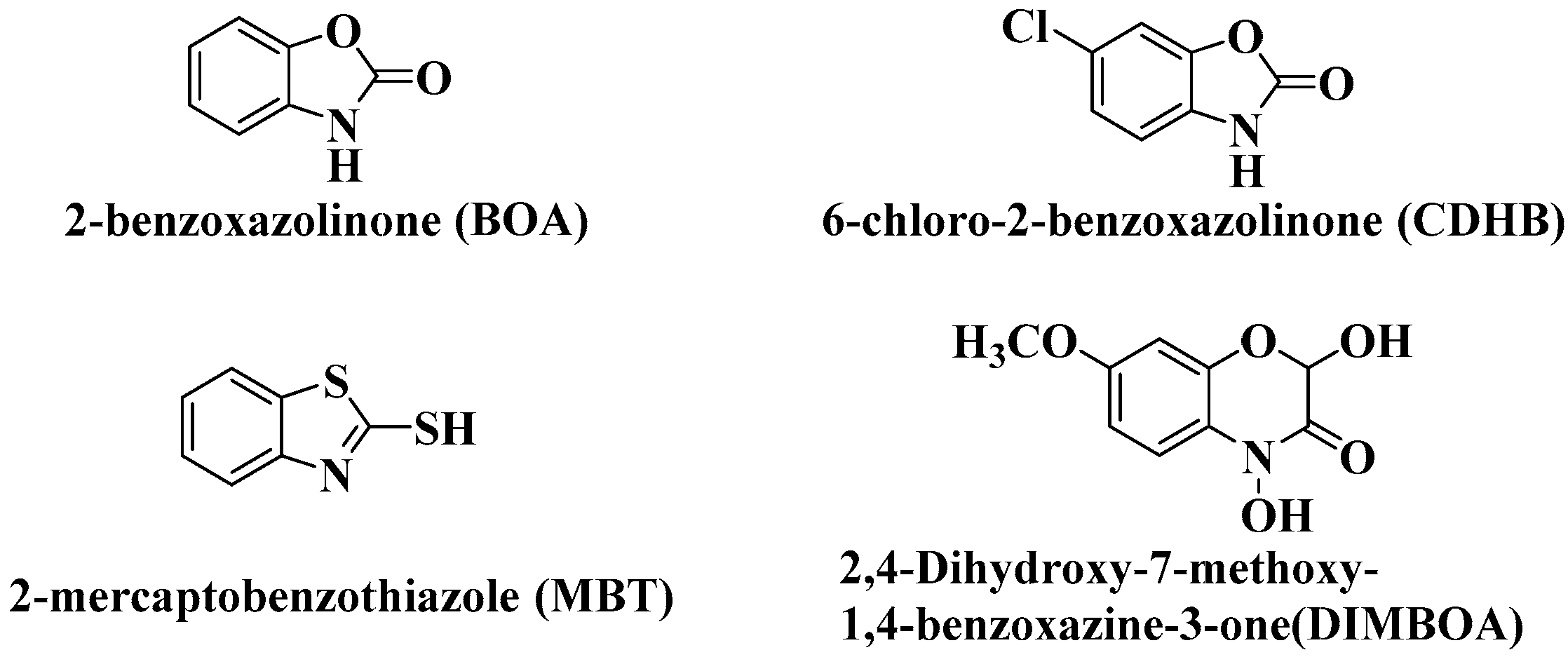
2.1. Determination of DIMBOA Antibacterial Activity
2.2. MIC and MBC of DIMBOA and Its Derivatives against R. solanacearum
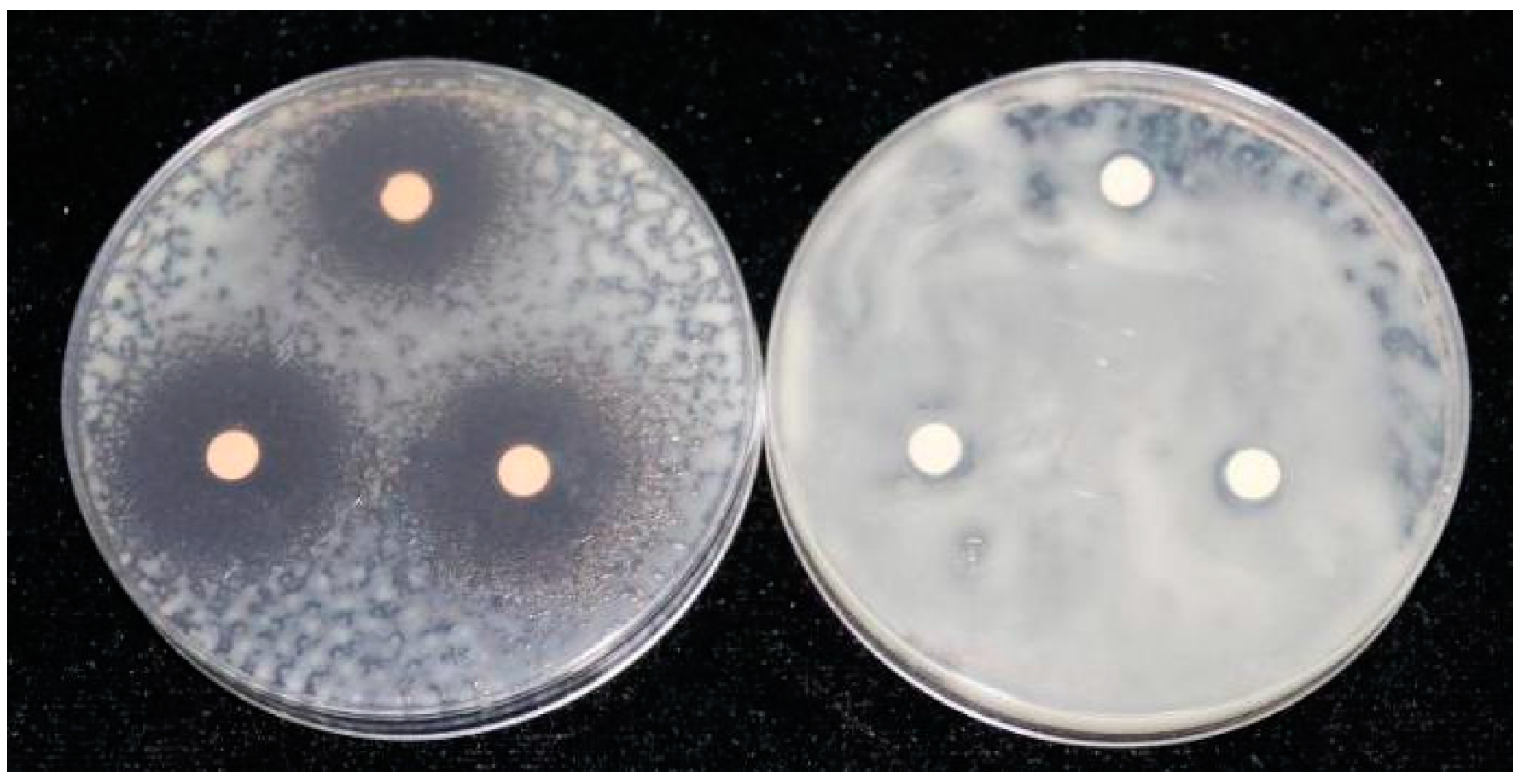
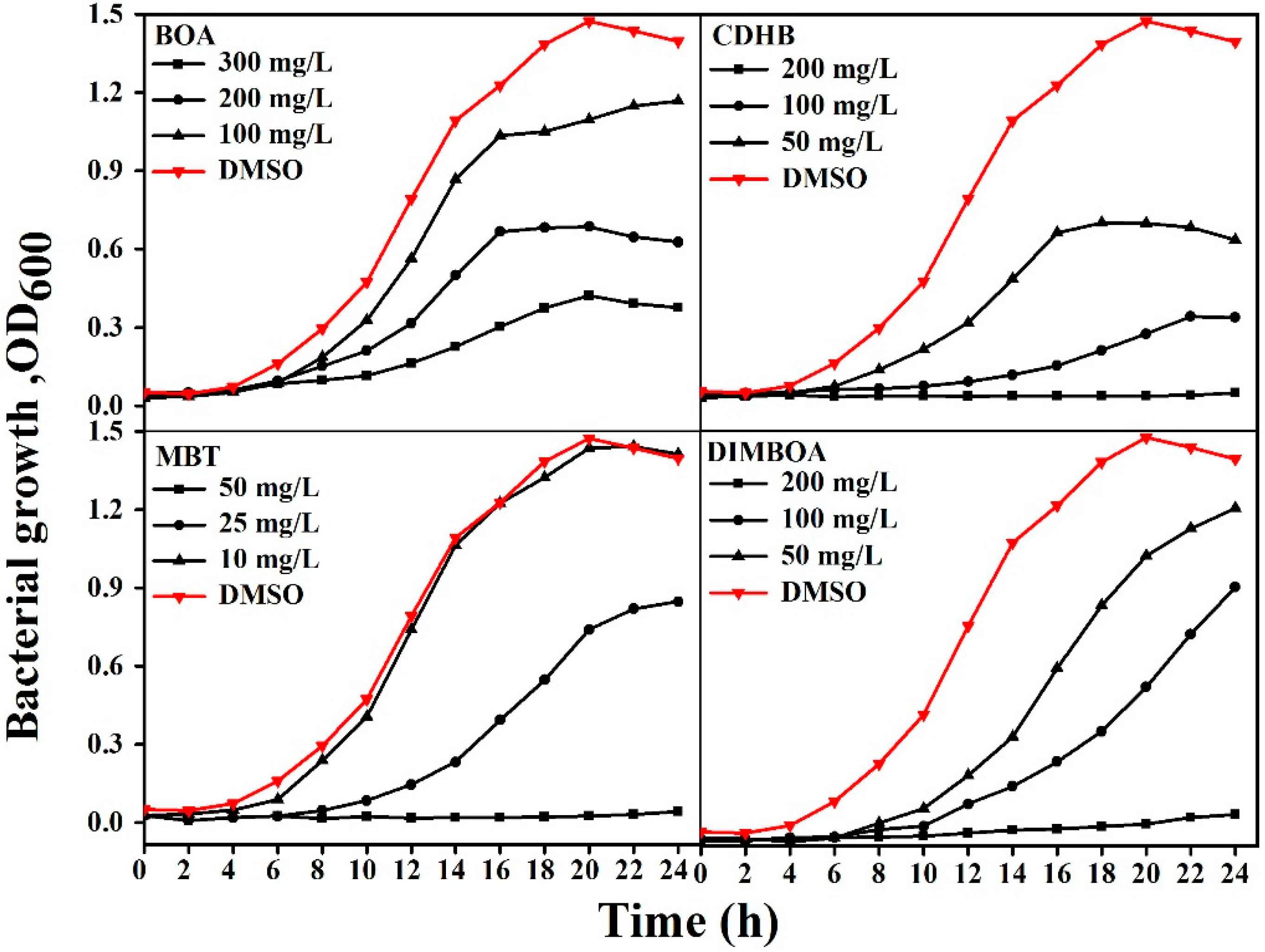
2.3. DIMBOA and Its Derivatives Inhibit the Growth of R. solanacearum
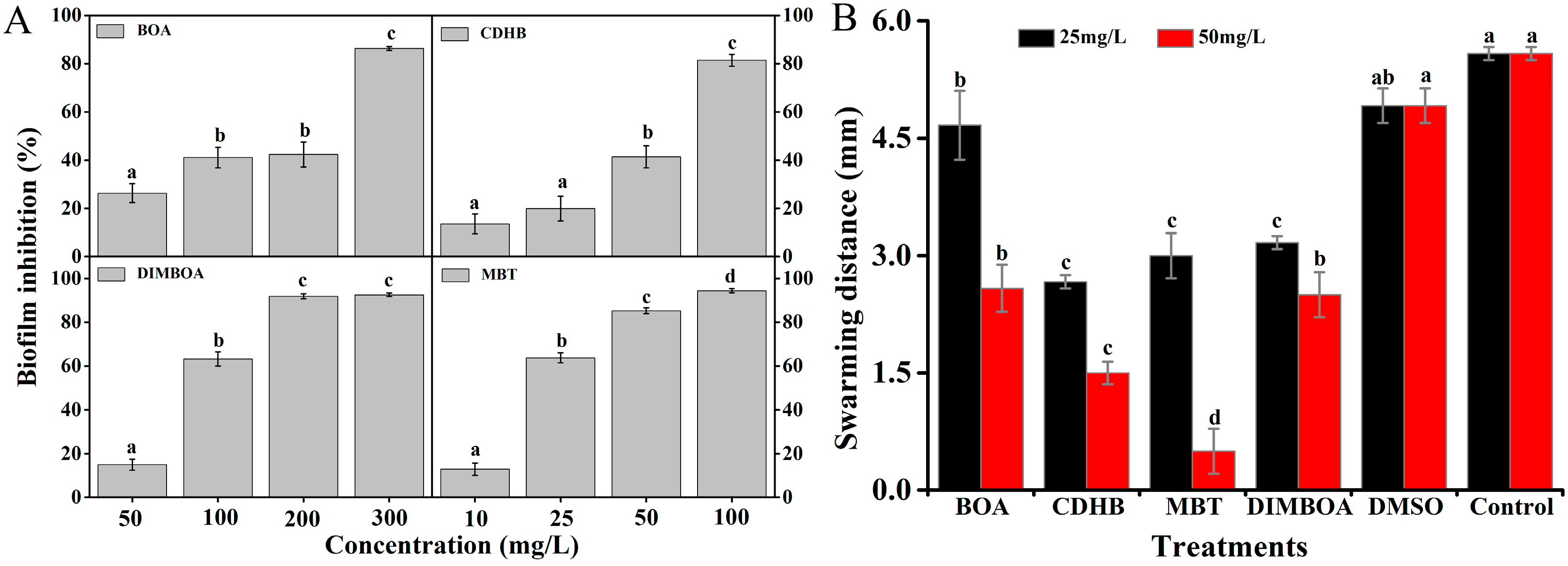
2.4. DIMBOA and Its Derivatives Reduce Biofilm Formation of R. solanacearum
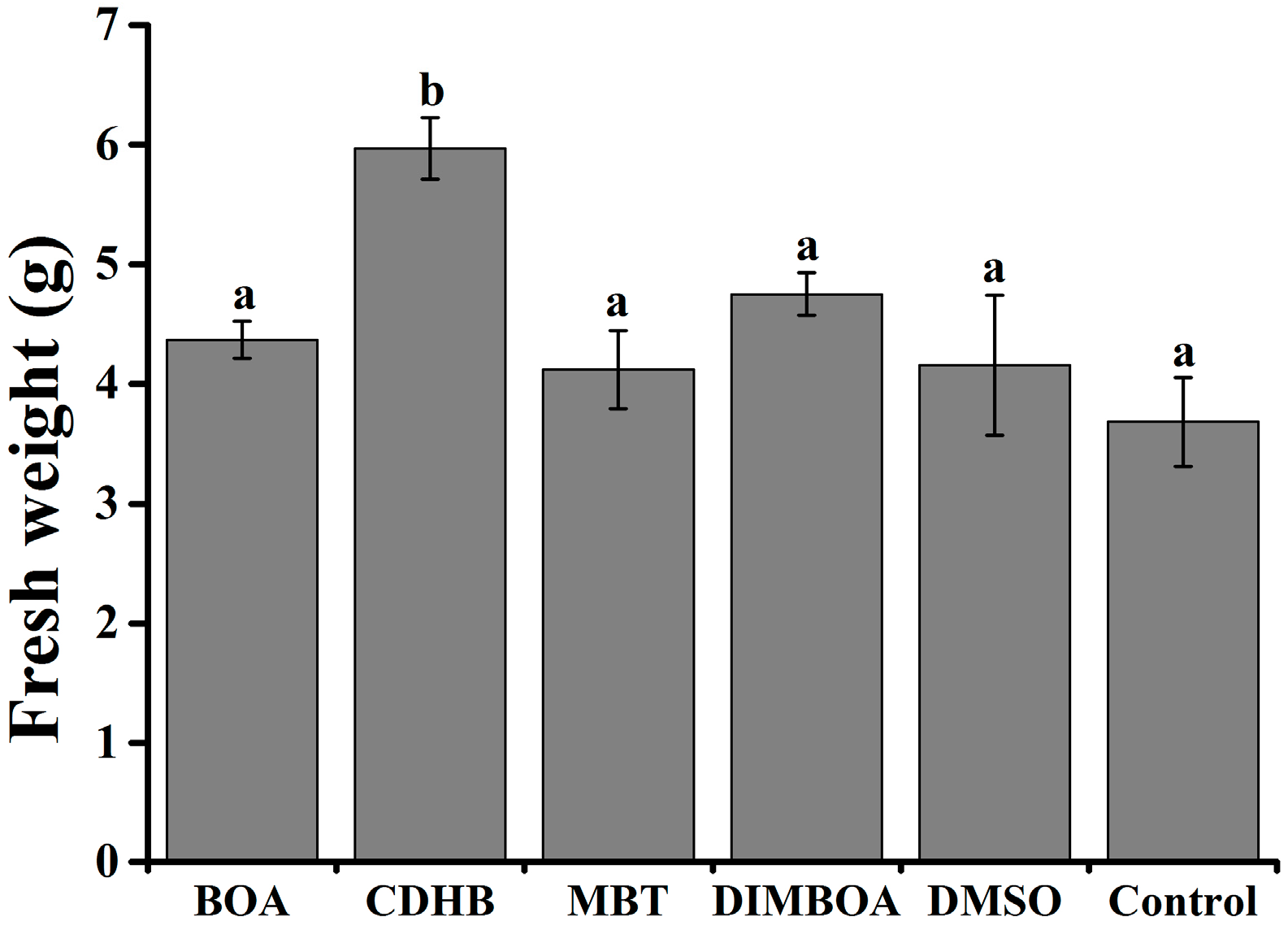
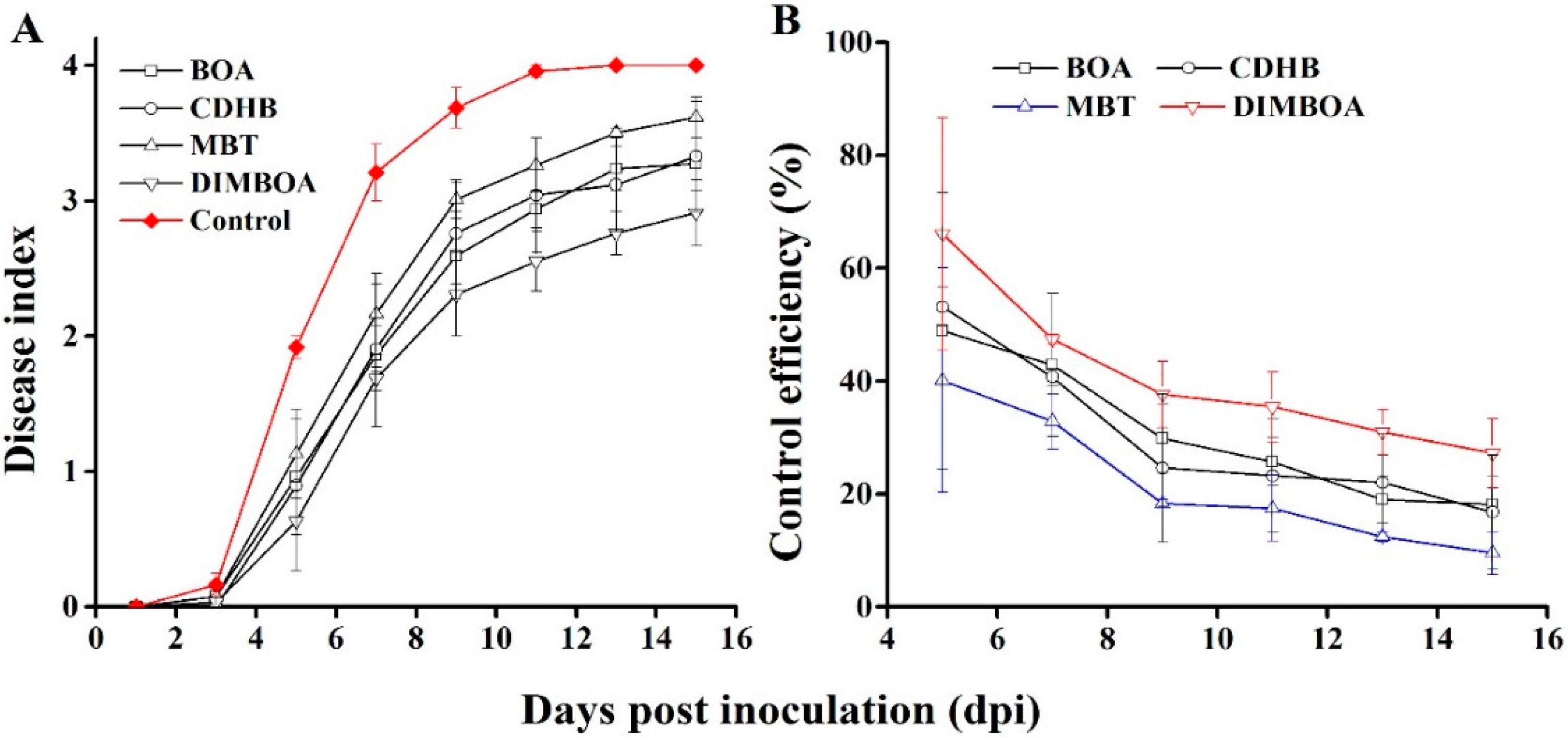
2.5. DIMBOA and Its Derivatives Enhanced the Resistance of Tobacco Seedlings to R. solanacearum
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Inoculum Preparation
4.2. Preparation of DIMBOA and Its Derivatives
4.3. Determination of DIMBOA Antibacterial Activity
4.4. MIC and MBC Assay
4.5. Bacterial Growth Curve Assay
4.6. Biofilm Formation Assay
4.7. Swarming Motility Assay
4.8. Greenhouse Experiments
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, L.; Feng, X.; Tang, M.; Hao, W.; Han, Y.; Zhang, G.; Wan, S. Antibacterial activity of Lansiumamide B to tobacco bacterial wilt (Ralstonia solanacearum). Microbiol. Res. 2014, 169, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Akiew, E.; Trevorrow, P.R.; Tonello, P.E. Management of bacterial wilt of tobacco. In Proceedings of the Bacterial Wilt Proceedings of an International Conference, Taiwan, 28–31 October 1992; Watson Ferguson and Company: Brisbane, Australia, 1993; pp. 270–275. [Google Scholar]
- Bailey, K.L.; Lazarovits, G. Suppressing soil-borne diseases with residue management and organic amendments. Soil. Till. Res. 2003, 72, 169–180. [Google Scholar] [CrossRef]
- Michel, V.V.; Hartman, G.L.; Midmore, D.J. Effect of previous crop on soil populations of Burkholderia solanacearum, bacterial wilt, and yield of tomatoes in Taiwan. Plant. Dis. 1996, 80, 1367–1372. [Google Scholar] [CrossRef]
- Niemeyer, H.M. Hydroxamic acids derived from 2-hydroxy-2H-1,4-benzoxazin-3(4H)-one: Key defense chemicals of cereals. J. Agric. Food Chem. 2009, 57, 1677–1696. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, K.B.; Laursen, B.B.; Gregersen, P.L.; Schnoor, H.J.; Witten, M.; Poulsen, L.K.; Jensen, B.M.; Fomsgaard, I.S. Absorption and metabolic fate of bioactive dietary benzoxazinoids in humans. Mol. Nutr. Food Res. 2013, 57, 1847–1858. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, H.M. Hydroxamic acids (4-hydroxy-1,4-benzoxazin-3-ones), defence chemicals in the Gramineae. Phytochemistry 1988, 27, 3349–3358. [Google Scholar] [CrossRef]
- Frey, M.; Chomet, P.; Glawischnig, E.; Stettner, C.; Grün, S.; Winklmair, A.; Eisenreich, W.; Bacher, A.; Meeley, R.B.; Briggs, S.P.; et al. Analysis of a chemical plant defense mechanism in grasses. Science 1997, 277, 696–699. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Bacon, C.W.; Richardson, M.D. Biotransformation of BOA and 6-methoxy-benzoxazolinone by Fusarium moniliforme. Phytochemistry 1998, 48, 451–454. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, Y.; Qi, L.; Mei, X.; Liao, J.; Ding, X.; Deng, W.; Fan, L.; He, X.; Vivanco, J.M.; et al. Plant-plant-microbe mechanisms involved in soil-borne disease suppression on a maize and pepper intercropping system. PLoS ONE 2014, 9, e115052. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, X.; Dong, F.; Xu, J.; Guo, L.Q.; Kong, Z.Q.; Tian, Y.Y.; Wu, Y.B.; Zheng, Y.Q. A simple method for the isolation and purification of 2,4-dihydroxy-7-methoxy-2H-1,4-benzoxazin-3(4H)-one (DIMBOA) from maize (Zea mays L.) seedlings. J. Integr. Agr. 2013, 12, 95–102. [Google Scholar] [CrossRef]
- Harput, U.S.; Arihan, O.; Iskit, A.B.; Nagatsu, A.; Saracoglu, I. Antinociceptive, free radical-scavenging, and cytotoxic activities of Acanthus hirsutus Boiss. J. Med. Food 2011, 14, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Peng, Q.; Selimi, D.; Wang, Q.; Charkowski, A.O.; Chen, X.; Yang, C.H. The plant phenolic compound p-coumaric acid represses gene expression in the Dickeya dadantii type III secretion system. Appl. Environ. Microbiol. 2009, 75, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hutchins, W.; Wu, X.; Liang, C.; Zhang, C.; Yuan, X.; Khokhani, D.; Chen, X.; Che, Y.; Wang, Q.; Yang, C.H. Derivative of plant phenolic compound inhibits the type III secretion system of Dickeya dadantii via HrpX/HrpY two-component signal transduction and Rsm systems. Mol. Plant Pathol. 2015, 16, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Khokhani, D.; Zhang, C.; Li, Y.; Wang, Q.; Zeng, Q.; Yamazakia, A.; Hutchins, W.; Zhou, S.S.; Chen, X.; Yang, C.H. Discovery of plant phenolic compounds that act as type III secretion system inhibitors or inducers of the fire blight pathogen, Erwinia amylovora. Appl. Environ. Microbiol. 2013, 79, 5424–5436. [Google Scholar] [CrossRef] [PubMed]
- Tans-Kersten, J.; Huang, H.; Allen, C. Ralstonia solanacearum needs motility for invasive virulence on tomato. J. Bacteriol. 2001, 183, 3597–3605. [Google Scholar] [CrossRef] [PubMed]
- Buchmann, C.A.; Nersesyan, A.; Kopp, B.; Schauberger, D.; Darroudi, F.; Grummt, T.; Krupitza, G.; Kundi, M.; Hermann, R.S.; Knasmueller, S. Dihydroxy-7-methoxy-1,4-benzoxazin-3-one (DIMBOA)and 2,4-dihydroxy-1,4-benzoxazin-3-one (DIBOA), two naturally occurring benzoxazinones contained in sprouts of Gramineae are potent aneugens in human-derived liver cells (HepG2). Cancer Lett. 2007, 246, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Corcuera, L.J.; Woodward, M.D.; Helgeson, J.P.; Kelman, A.; Upper, C.D. 2,4-Dihydroxy-7-methoxy-2H-1, 4-benzoxazin-3(4H)-one, an inhibitor from Zea mays with differential activity against soft rotting Erwinia species. Plant Physiol. 1978, 61, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.Y.; Cao, M.; Xie, L.J.; Liang, X.T.; Zeng, R.S.; Su, Y.J.; Huang, J.H.; Wang, R.L.; Luo, S.M. Induction of DIMBOA accumulation and systemic defense responses as a mechanism of enhanced resistance of mycorrhizal corn (Zea mays L.) to sheath blight. Mycorrhiza 2011, 21, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Havlin, J.L.; Kissel, D.E.; Maddux, L.D.; Claassen, M.M.; Long, J.H. Crop rotation and tillage effects on soil organic carbon and nitrogen. Soil Sci. Soc. Am. J. 1990, 54, 448–452. [Google Scholar] [CrossRef]
- Balota, E.L.; Colozzi-Filho, A.; Andrade, D.S.; Dick, R.P. Microbial biomass in soils under different tillage and crop rotation systems. Biol. Fertil. Soils 2003, 38, 15–20. [Google Scholar] [CrossRef]
- Cuevas, L.; Niemeyer, H.M.; Jonsson, L.M.V. Partial purification and characterization of a hydroxamic acid glucoside β-d-glucosidase from maize. Phytochemistry 1992, 31, 2609–2612. [Google Scholar] [CrossRef]
- Friebe, A.; Vilich, V.; Hennig, L.; Kluge, M.; Sicker, D. Detoxification of benzoxazolinone allelochemicals from wheat by Gaeumannomyces graminis var. tritici, G. graminis var. graminis, G. graminis var. avenae, and Fusarium culmorum. Appl. Environ. Microbiol. 1998, 64, 2386–2391. [Google Scholar] [PubMed]
- Cambier, V.; Hance, T.; de-Hoffmann, E. Non-injured maize contains several 1,4-benzoxazin-3-one related compounds but only as glucoconjugates. Phytochem. Anal. 1999, 10, 119–126. [Google Scholar] [CrossRef]
- Cambier, V.; Hance, T.; de-Hoffmann, E. Variations of DIMBOA and related compounds content in relation to the age and plant organ in maize. Phytochemistry 2000, 53, 223–229. [Google Scholar] [CrossRef]
- Gleńsk, M.; Gajda, B.; Franiczek, R.; Krzyżanowskab, B.; Biskupa, I.; Włodarczyka, M. In vitro evaluation of the antioxidant and antimicrobial activity of DIMBOA [2,4-dihydroxy-7-methoxy-2H-1, 4-benzoxazin-3 (4H)-one]. Nat. Prod. Res. 2016, 30, 1305–1308. [Google Scholar] [CrossRef] [PubMed]
- Cambier, V.; Hance, T.; de-Hoffmann, E. Effects of 1,4-benzoxazin-3-one derivatives from maize on survival and fecundity of Metopolophium dirhodum (Walker) on artificial diet. J. Chem. Ecol. 2001, 27, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Macías, F.A.; Chinchilla, N.; Varela, R.M.; Oliverors-Bastidas, A.; Marín, D.; Molinillo, J.M.G. Structure-activity relationship studies of benzoxazinones and related compounds. Phytotoxicity on Echinochloa crus-galli (L.) P. Beauv. J. Agric. Food Chem. 2005, 53, 4373–4380. [Google Scholar] [CrossRef] [PubMed]
- Macias, F.A.; Chinchilla, N.; Arroyo, E.; Molinillo, J.M.G.; Marin, D.; Varela, R.M. Combined strategy for phytotoxicity enhancement of benzoxazinones. J. Agric. Food Chem. 2010, 58, 2047–2053. [Google Scholar] [CrossRef] [PubMed]
- Neal, A.L.; Ahmad, S.; Gordon-Weeks, R.; Ton, J. Benzoxazinoids in root exudates of maize attract Pseudomonas putida to the rhizosphere. PLoS ONE 2012, 7, e35498. [Google Scholar] [CrossRef] [PubMed]
- Park, W.J.; Hochholdinger, F.; Gierl, A. Release of the benzoxazinoids defense molecules during lateral-and crown root emergence in Zea mays. J. Plant Physiol. 2004, 161, 981–985. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Boone, L.; Kocz, R.; Zhang, C.; Binns, A.N.; Lynn, D.G. At the maize/Agrobacterium interface: Natural factors limiting host transformation. Chem. Biol. 2000, 7, 611–621. [Google Scholar] [CrossRef]
- Macías, F.A.; Oliveros-Bastidas, A.; Marín, D.; Castellano, D.; Simonet, A.M.; Molinillo, J.M.G. Degradation studies on benzoxazinoids. Soil degradation dynamics of 2,4-dihydroxy-7-methoxy-(2H)-1, 4-benzoxazin-3 (4H)-one (DIMBOA) and its degradation products, phytotoxic allelochemicals from Gramineae. J. Agric. Food Chem. 2004, 52, 6402–6413. [Google Scholar] [CrossRef] [PubMed]
- Fomsgaard, I.S.; Mortensen, A.G.; Carlsen, S.C.K. Microbial transformation products of benzoxazolinone and benzoxazinone allelochemicals––A review. Chemosphere 2004, 54, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.J.; Zheng, Y.Q.; Kong, C.H.; Zhang, S.Z.; Li, J.; Liu, X.G. 2, 4-Dihydroxy-7-methoxy-1,4-benzoxazin-3-one (DIMBOA) and 6-methoxy-benzoxazolin-2-one (MBOA) levels in the wheat rhizosphere and their effect on the soil microbial community structure. J. Agric. Food Chem. 2010, 58, 12710–12716. [Google Scholar] [CrossRef] [PubMed]
- Linderman, R.G. Mycorrhizal interactions with the rhizosphere microflora: the mycorrhizosphere effect. Phytopathology 1988, 78, 366–371. [Google Scholar]
- Wu, D.; Ding, W.; Zhang, Y.; Liu, X.; Yang, L. Oleanolic acid induces the type III secretion system of Ralstonia solanacearum. Front. Microbiol. 2015, 6, 1466. [Google Scholar] [CrossRef] [PubMed]
- Yoshimochi, T.; Zhang, Y.; Kiba, A.; Hikichi, Y.; Ohnishi, K. Expression of hrpG and activation of response regulator HrpG are controlled by distinct signal cascades in Ralstonia solanacearum. J. Gen. Plant Pathol. 2009, 75, 196–204. [Google Scholar] [CrossRef]
- Larsen, E.; Christensen, L.P. Simple method for large scale isolation of the cyclic arylhydroxamic acid DIMBOA from maize (Zea mays L.). J. Agr. Food Chem. 2000, 48, 2556–2558. [Google Scholar] [CrossRef]
- Woodward, M.D.; Corcuera, L.J.; Helgeson, J.P.; Upper, C.D. Decomposition of 2,4-dihydroxy-7-methoxy-2H-1,4-benzoxazin-3(4H)-one in aqueous solutions. Plant Physiol. 1978, 61, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Lyons, P.C.; Hipskind, J.D.; Wood, K.V.; Nicholson, R.L. Separation and quantification of cyclic hydroxamic acids and related compounds by high-pressure liquid chromatography. J. Agric. Food Chem. 1988, 36, 57–60. [Google Scholar] [CrossRef]
- Atkinson, J.; Morand, P.; Arnason, J.T.; Niemeyer, H.M.; Bravo, H.R. Analogs of the cyclic hydroxamic acid 2,4-dihydroxy-7-methoxy-2H-1,4-benzoxazin-3-one (DIMBOA): Decomposition to benzoxazolinones and reaction with. β-mercaptoethanol. J. Org. Chem. 1991, 56, 1788–1800. [Google Scholar] [CrossRef]
- Knölker, H.J.; Braxmeier, T. Isocyanates, part 5 Synthesis of chiral oxazolidin-2-ones and imidazolidin-2-ones via DMAP-catalyzed isocyanation of amines with di-tert-butyl dicarbonate. Tetrahedron Lett. 1998, 39, 9407–9410. [Google Scholar] [CrossRef]
- Yang, L.; Ding, W.; Xu, Y.; Wu, D.; Li, S.; Chen, J.; Guo, B. New Insights into the Antibacterial Activity of Hydroxycoumarins against Ralstonia solanacearum. Molecules 2016, 21, 468. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Serio, A.W.; Pechter, K.B.; Sonenshein, A.L. Bacillus subtilis aconitase is required for efficient late sporulation gene expression. J. Bacteriol. 2006, 188, 6396–6405. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yu, Y.; Chen, J.; Guo, B.; Yang, L.; Ding, W. Evaluation of the antibacterial effects and mechanism of action of protocatechualdehyde against Ralstonia solanacearum. Molecules 2016, 21, 754. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Yang, C.; Mei, X.; Shen, S.; Raza, W.; Shen, Q.; Xu, Y. The effect of organic acids from tomato root exudates on rhizosphere colonization of Bacillus amyloliquefaciens T-5. Appl. Soil Ecol. 2013, 64, 15–22. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the DIMBOA and its derivatives are available from the authors.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (μg/disc) | Inhibition Zone (mm) | |||
|---|---|---|---|---|
| BOA | CDHB | MBT | DIMBOA | |
| 0 | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 | 0.0 ± 0.00 |
| 10 | 0.0 ± 0.00 a | 2.4 ± 0.22 a | 3.5 ± 0.29 a | 1.5 ± 0.29 a |
| 50 | 1.2 ± 0.17 a | 4.3 ± 0.33 a | 7.0 ± 0.58 ab | 4.5 ± 0.29 b |
| 100 | 5.0 ± 0.58 b | 10.5 ± 0.50 b | 8.0 ± 0.58 b | 4.5 ± 0.29 b |
| 200 | 9.67 ± 0.83 c | 17.7 ± 0.44 c | 11.0 ± 0.50 b | 13.7 ± 0.33 c |
| 300 | 10.83 ± 0.93 c | 22.0 ± 1.26 d | 17.7 ± 2.73 c | 19.3 ± 0.33 d |
| DIMBOA and Its Derivatives | MIC (mg/L) | MBC (mg/L) |
|---|---|---|
| BOA | 300 | >1000 |
| CDHB | 100 | 500 |
| MBT | 50 | 400 |
| DIMBOA | 200 | 500 |
| DIMBOA and Its Derivatives | Regression Equations | IC50 (mg/ L) | IC90 (mg/L) | R Value | |
|---|---|---|---|---|---|
| BOA | Y = 2.0144x + 0.3265 | 208.92 | 904.05 | 0.9938 | |
| CDHB | Y = 1.0285x + 3.4861 | 29.65 | 522.50 | 0.9733 | |
| MBT | Y = 0.9786x + 4.1034 | 8.25 | 168.18 | 0.8930 | |
| DIMBOA | Y = 1.6101x + 2.1541 | 58.55 | 366.03 | 0.9015 | |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, B.; Zhang, Y.; Li, S.; Lai, T.; Yang, L.; Chen, J.; Ding, W. Extract from Maize (Zea mays L.): Antibacterial Activity of DIMBOA and Its Derivatives against Ralstonia solanacearum. Molecules 2016, 21, 1397. https://doi.org/10.3390/molecules21101397
Guo B, Zhang Y, Li S, Lai T, Yang L, Chen J, Ding W. Extract from Maize (Zea mays L.): Antibacterial Activity of DIMBOA and Its Derivatives against Ralstonia solanacearum. Molecules. 2016; 21(10):1397. https://doi.org/10.3390/molecules21101397
Chicago/Turabian StyleGuo, Bing, Yongqiang Zhang, Shili Li, Ting Lai, Liang Yang, Juanni Chen, and Wei Ding. 2016. "Extract from Maize (Zea mays L.): Antibacterial Activity of DIMBOA and Its Derivatives against Ralstonia solanacearum" Molecules 21, no. 10: 1397. https://doi.org/10.3390/molecules21101397
APA StyleGuo, B., Zhang, Y., Li, S., Lai, T., Yang, L., Chen, J., & Ding, W. (2016). Extract from Maize (Zea mays L.): Antibacterial Activity of DIMBOA and Its Derivatives against Ralstonia solanacearum. Molecules, 21(10), 1397. https://doi.org/10.3390/molecules21101397