
Laurus nobilis, Zingiber officinale and Anethum graveolens Essential Oils: Composition, Antioxidant and Antibacterial Activities against Bacteria Isolated from Fish and Shellfish

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
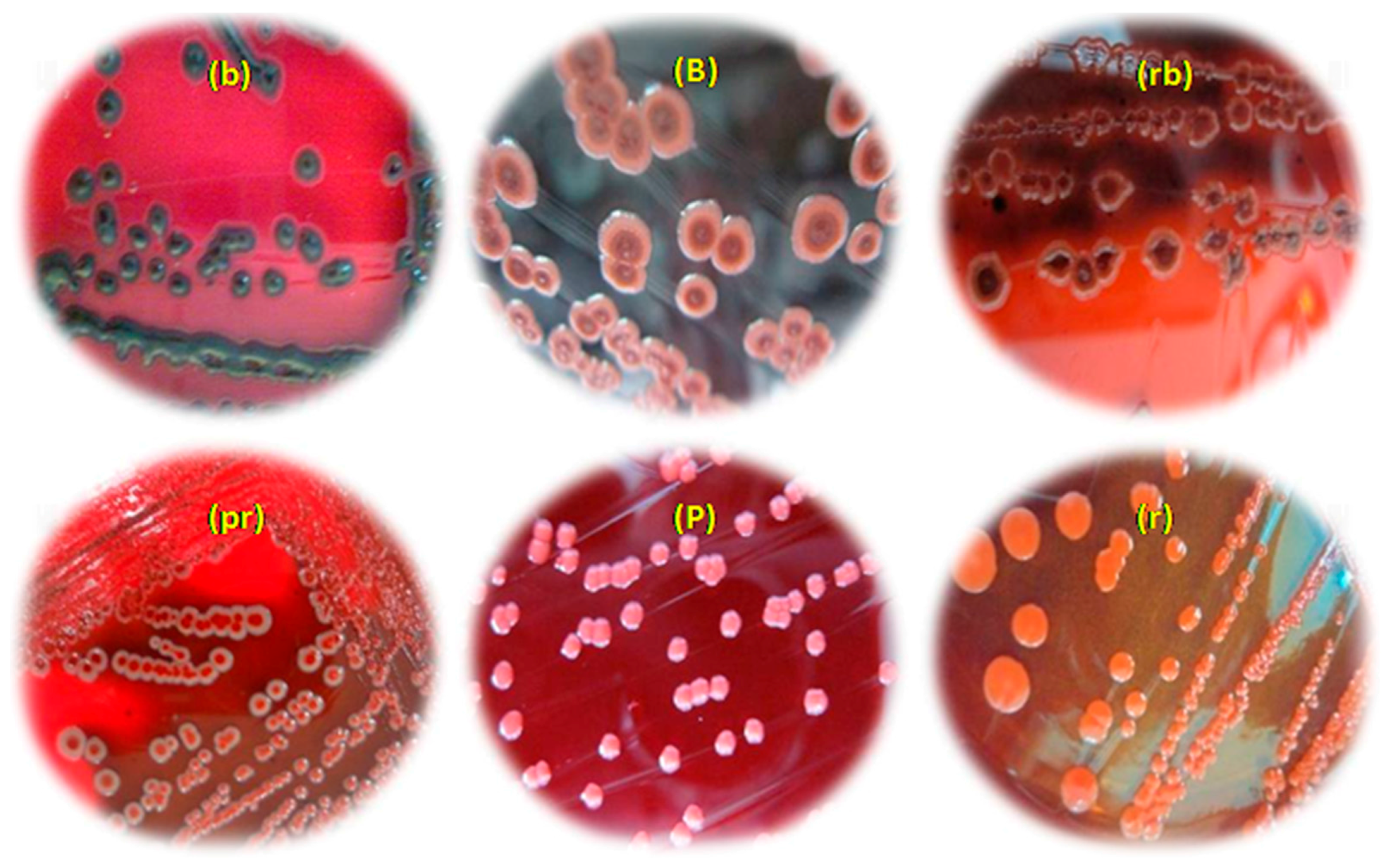
2.1. Identification and Characterization of Isolates
2.2. Chemical Composition of the Essential Oils
2.3. Antioxidant Activities
2.4. Antibacterial Activities
3. Discussion
3.1. Identification and Characterization of Isolates
3.2. Chemical Composition of the Essential Oils
3.3. Antioxidant Activities
3.4. Antibacterial Activities
4. Materials and Methods
4.1. Sampling and Strains Isolation
4.2. Identification and Biochemical Characterization of Bacterial Strains
4.3. Adhesive Properties of the Selected Strains
4.4. Plant Material, Extraction and Chemical Characterization of the Essential Oil
4.5. Antioxidant Properties
4.5.1. DPPH Radical-Scavenging Activity
4.5.2. Superoxide Anion Scavenging Activity
4.5.3. Determination of Reducing Power
4.5.4. β-Carotene-linoleic Acid Model System
4.6. Antimicrobial Activities
4.6.1. Microorganisms
4.6.2. Disk-Diffusion Assay
4.6.3. Microdilution Method for the Determination of the MICs and MBCs
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Jiang, M.; Wang, Y.; Van Santen, E.; Chappell, J.A. Evaluation of textural properties of channel catfish (Ictalurus punctatus Rafinesque) fillet with the natural contour method. LWT-Food Sci. Technol. 2008, 41, 1548–1554. [Google Scholar] [CrossRef]
- Boularès, M.; Mejri, L.; Hassouna, M. Study of the microbial ecology of wild and aquacultured Tunisian fresh fish. J. Food Prot. 2011, 74, 1762–1768. [Google Scholar] [CrossRef] [PubMed]
- Al-Hajj, N.Q.M.; Rashid, H.; Al-Zaemy, A.B.; Alzoreky, N.; Wang, H.; Thabit, R.; Siddeeq, A. Food Additives and their use in Destruction of Some Seafood Poisoning Bacteria. EAR 2014, 2, 8749–8774. [Google Scholar]
- Al-Hajj, N.Q.M.; Alzoreky, N.; Al-Zaimy, A.; Wang, H.; Asamet, S.; Noor, Y.; Thabit, R. Destruction of some food poisoning bacteria and shelf-life extension of seafood. JAIR 2014, 3, 10–15. [Google Scholar]
- Poli, M.B.; Parisi, G.; Zambacavallo, G.; Mecatti, M.; Lupi, P.; Gualtieri, M.; Franci, O. Quality outline of European sea bass (Dicentrarchus labrax) reared in Italy: Shelf life, edible yield, nutritional and dietetic traits. Aquaculture 2001, 202, 303–313. [Google Scholar] [CrossRef]
- Gram, L.; Dalgaard, P. Fish spoilage bacteria—Problems and solutions. Curr. Opin. Biotechnol. 2002, 13, 262–266. [Google Scholar] [CrossRef]
- Huss, H.H.; Reilly, A.; Ben Embarek, P.K. Prevention and control of hazards in seafood. Food Control 2000, 11, 149–156. [Google Scholar] [CrossRef]
- Carraro, V.; Sanna, C.; Brandas, V.; Sanna, A.; Pinna, A.; Coroneo, V. Hygiene and health risks associated with the consumption of edible lamellibranch molluscs. Int. J. Food Microbiol. 2015, 201, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Muli, R.; Giljanovi, S.; Ropac, D.; Katalini, V. Some epidemiologic characteristics of foodborne intoxications in Croatia during the 1992–2001 period. Acta Med. Croat. 2004, 58, 421–427. [Google Scholar]
- Boulares, M.; Mankai, M.; Aouadhi, C.; Ben Moussa, O.; Hassouna, M. Characterisation and identification of spoilage psychotrophic Gram-negative bacteria originating from Tunisian fresh fish. Ann. Microbiol. 2013, 63, 733–744. [Google Scholar] [CrossRef]
- Serio, A.; Fusella, G.C.; Chaves-Lopez, C.; Sacchetti, G.; Paparella, A. A survey on bacteria isolated as hydrogen sulfide-producers from marine fish. Food Control 2014, 39, 111–118. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food. Microbiol. 2004, 4, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. App. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef]
- Snoussi, M.; Noumi, E.; Trabelsi, N.; Flamini, G.; Papetti, A.; de Feo, V. Mentha spicata essential oil: Chemical composition, antioxidant and antibacterial activities against planktonic and biofilm cultures of Vibrio spp. strains. Molecules 2015, 20, 14402–14424. [Google Scholar] [CrossRef] [PubMed]
- Seden, M.E.A.; Abbass, F.E.; Ahmed, M.H. Effect of Origanum vulgare as a feed additive on growth performance, feed utilization and whole body composition of Nile tilapia (Oreochromis niloticus) fingerlings challenged with pathogenic Aeromonas hydrophila. J. Agric. Sci. Mansoura Univ. 2009, 34, 1683–1695. [Google Scholar]
- Shehata, S.A.; Mohamed, M.S.; Abd El-Shafi, S. Antibacterial Activity of Essential Oils and their Effects on Nile Tilapia Fingerlings Performance. J. Med. Sci. 2013, 13, 367–372. [Google Scholar] [CrossRef]
- Hassaan, M.S.; Soltan, M.A. Evaluation of Essential Oil of Fennel and Garlic Separately or Combined with Bacillus licheniformis on the Growth, Feeding Behaviour, Hemato-biochemical Indices of Oreochromis niloticus (L.) Fry. J. Aquac. Res. Dev. 2016, 7, 422–429. [Google Scholar] [CrossRef]
- FDA. Food and Drug Administration of the US, 21 CFR 184; Department Of Health And Human Services: Silver Spring, MD, USA, 2016.
- Sharopov, F.S.; Wink, M.; Gulmurodov, I.S.; Isupov, S.J.; Zhang, H.; Setzer, W.N. Composition and bioactivity of the essential oil of Anethum graveolens L. from Tajikistan. Int. J. Med. Arom. Plants 2013, 3, 125–130. [Google Scholar]
- Baytop, T. Therapy with Medicinal Plants in Turkey (Past and Present), 2nd ed.; Nobel Tıp Press: Istanbul, Turkey, 1999; pp. 13–31. [Google Scholar]
- Merghni, A.; Marzouki, H.; Hentati, H.; Aouni, M.; Mastouri, M. Antibacterial and antibiofilm activities of Laurus nobilis L. essential oil against Staphylococcus aureus strains associated with oral infections. Pathol. Biol. 2016, 64, 29–34. [Google Scholar]
- Chmit, M.; Kanaan, H.; Habib, J.; Abbass, M.; Mcheik, A.; Chokr, A. Antibacterial and antibiofilm activities of polysaccharides, essential oil, and fatty oil extracted from Laurus nobilis growing in Lebanon. Asian Pac. J. Trop. Med. 2014, 7, 546–552. [Google Scholar] [CrossRef]
- Jana, S.; Shekhawat, G.S. Anethum graveolens: An Indian traditional medicinal herb and spice. Pharmacogn. Rev. 2010, 4, 179–184. [Google Scholar] [PubMed]
- Chen, Y.; Zeng, H.; Tian, J.; Ban, X.; Ma, B.; Wang, Y. Dill (Anethum graveolens L.) seed essential oil induces Candida albicans apoptosis in a metacaspase-dependent manner. Fungal Biol. 2014, 118, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, M. Phenolic profile, antioxidant capacity and anti-inflammatory activity of Anethum graveolens L. essential oil. Nat. Prod. Res. 2015, 29, 551–553. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Kapoor, I.P.S.; Singh, P.; de Heluani, C.S.; de Lampasona, M.P.; Catalan, C.A.N. Chemistry, antioxidant and antimicrobial investigations on essential oil and oleoresins of Zingiber officinale. Food. Chem. Toxicol. 2008, 46, 3295–3302. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Karthik, L.; Rao, K.V.B. A Review on Pharmacological and Phytochemical Properties of Zingiber officinale Roscoe (Zingiberaceae). I. Pharm. Res. 2011, 4, 2963–2966. [Google Scholar]
- Croci, L.; Serratore, P.; Cozzi, L.; Stacchini, A.; Milandri, S.; Suffredini, E.; Toti, L. Detection of Vibrionaceae in mussels and in their seawater growing area. Lett. Appl. Microbiol. 2001, 32, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Saidi, N.; Snoussi, M.; Usai, D.; Zanetti, S.; Bakhrouf, A. Adhesive properties of Aeromonas hydrophila strains isolated from Tunisian aquatic biotopes. Afr. J. Microbiol. Res. 2011, 5, 5644–5655. [Google Scholar]
- Khouadja, S.; Snoussi, M.; Saidi, N.; Bakhrouf, A. Phenotypic characterization and RAPD fingerprinting of Vibrio parahaemolyticus and Vibrio alginolyticus isolated during Tunisian fish farm outbreaks. Folia Microbiol. 2013, 58, 17–26. [Google Scholar]
- Hussein Ali, H. Isolation and identification of Staphylococcus bacteria from fish of fresh water and its antibiotics sensitivity in Mosul city. Bas. J. Vet. Res. 2014, 1, 33–42. [Google Scholar]
- Snoussi, M.; Noumi, E.; Hajlaoui, H.; Usai, D.; Sechi, L.A.; Zanetti, S.; Bakhrouf, A. High potential of adhesion to abiotic and biotic materials in fish aquaculture facility by Vibrio alginolyticus strains. J. Appl. Microbiol. 2009, 106, 1591–1599. [Google Scholar] [CrossRef] [PubMed]
- Snoussi, M.; Noumi, E.; Cheriaa, J.; Usai, D.; Sechi, L.A.; Zanetti, S.; Bakhrouf, A. Adhesive properties of environmental Vibrio alginolyticus strains to biotic and abiotic surfaces. New Microbiol. 2008, 31, 489–500. [Google Scholar] [PubMed]
- Sechi, L.A.; Deriu, A.; Falchi, M.P.; Fadda, G.; Zanetti, S. Distribution of virulence genes in Aeromonas spp. Isolated from Sardinian waters and from patients with diarrhoea. J. Appl. Microbiol. 2002, 92, 221–227. [Google Scholar]
- Thenmozhi, S.; Vijayalakshmi, P.; Moorthy, K.; Sureshkumar, B.T. Virulence and molecular characterization of Aeromonas spp. from fish samples using ARDRA technique. Int. J. Pharm. Biol. Sci. 2015, 6, 593–603. [Google Scholar]
- Ben Abdallah, F.; Chaieb, K.; Zmantar, T.; Kallel, H.; Bakhrouf, A. Adherence assays and Slime production of Vibrio alginolyticus and Vibrio parahaemolyticus. Braz. J. Microbiol. 2009, 40, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Sáncheza, D.; Habimanab, O.; Holck, A. Impact of food-related environmental factors on the adherence and biofilm formation of natural Staphylococcus aureus isolates. Curr. Microbiol. 2013, 66, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Ben Jemâa, J.M.; Tersim, N.; Toudert, K.T.; Khouja, M.L. Insecticidal activities of essential oils from leaves of Laurus nobilis L. from Tunisia, Algeria and Morocco, and comparative chemical composition. J. Stored Prod. Res. 2012, 48, 97–104. [Google Scholar] [CrossRef]
- Marzouki, H.; Khaldi, A.; Elaisse, A.; Bouzid, S.; Piras, A.; Porcedda, S.; Tuveri, E.; Marongiu, B. Isolation of the oils from Laurus nobilis of Tunisia and Algeria by supercritical carbon dioxide extraction. In Proceedings of the 1st International Congress on the Chemistry of Natural Products (ICNP-2002), Trabzon, Turkey, 16–19 October 2002; pp. 172–179.
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Peerakam, N.; Wattanathorn, J.; Punjaisee, S.; Buamongkol, S.; Sirisa-ard, P.; Chansakaow, S. Chemical Profiling of Essential Oil Composition and Biological Evaluation of Anethum graveolens L. (Seed) Grown in Thailand. J. Nat. Sci. Res. 2014, 4, 34–41. [Google Scholar]
- Radulescu, V.; Popescu, M.L.; Ilieş, D.C. Chemical composition of the volatile oil from different plant parts of Anethum graveolens L. (Umbelliferae) cultivated in Romania. Farmacia 2010, 58, 594–600. [Google Scholar]
- Jirovetz, L.; Buchbauer, G.; Stoyanova, A.S.; Georgiev, E.V.; Damianova, S.T. Composition, quality control, and antimicrobial activity of the essential oil of long-time stored dill (Anethum graveolens L.) seeds from Bulgaria. J. Agric. Food Chem. 2003, 51, 3854–3857. [Google Scholar] [CrossRef] [PubMed]
- Bailer, J.; Aichinger, T.; Hackl, G.; de Hueber, K.; Dachler, M. Essential oil content and composition in commercially available dill cultivars in comparison to caraway. Ind. Crop. Prod. 2001, 14, 229–239. [Google Scholar] [CrossRef]
- Sintim, H.Y.; Burkhardt, A.; Gawde, A.; Cantrell, C.L.; Astatkie, T.; Obour, A.E.; Zheljazkov, V.D.; Schlegel, V. Hydrodistillation time affects dill seed essential oil yield, composition, and bioactivity. Ind. Crop. Prod. 2015, 63, 190–196. [Google Scholar] [CrossRef]
- Agrawal, M.; Walia, S.; Dhingra, S.; Khambay, B.P.S. Insect growth inhibition, antifeedant and antifungal activity of compounds isolated/derived from Zingiber officinale Roscoe (ginger) rhizomes. Pest. Manag. Sci. 2001, 57, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Menut, C.; Lanaty, G.; Bessiere, J.M.; Koudou, J. Aromatic Plants of Tropical Central Africa. XIII. Rhizomes Volatile Components of Two Zingiberales from the Central African Republic. J. Essent. Oil. Res. 1994, 6, 101–108. [Google Scholar] [CrossRef]
- Singh, S.; Das, S.S.; Singh, G.; Schuff, C.; de Lampasona, M.P.; Catalán, C.A.N. Composition, In Vitro Antioxidant and Antimicrobial Activities of Essential Oil and Oleoresins Obtained from Black Cumin Seeds (Nigella sativa L.). Bio. Med. Res. Int. 2014, 2014, 1–10. [Google Scholar]
- Singh, G.; Maurya, S.; Catalan, C.; Lampasona, M.P. Studies on essential oils, Part 42: Chemical, antifungal, antimicrobial and sprout suppressant studies on ginger essential oil and its oleoresin. Flavour. Frag. J. 2005, 20, 1–6. [Google Scholar] [CrossRef]
- Sivasothy, Y.; Chong, W.K.; Hamid, A.; Eldeen, I.M.; Sulaiman, S.F.; Awang, K. Essential oils of Zingiber officinale var rubrum Theilade and their antibacterial activities. Food Chem. 2011, 124, 514–517. [Google Scholar] [CrossRef]
- Yeh, H.U.; Chuang, C.H.; Chen, H.C.; Wan, C.J.; Chen, T.L.; Lin, L.Y. Bioactive components analysis of two various gengers (Zingiber officinale Roscoe) and antioxidant effect of ginger extracts. LWT-Food Sci. Technol. 2014, 55, 329–334. [Google Scholar] [CrossRef]
- Cherrat, L.; Espina, L.; Garcia-Gonzalo, D.; Bakkali, M.; Pagan, R.; Laglaoui, A. Chemical composition and antioxidant properties of Laurus nobilis L. and Myrtus communis L. essential oils from Morocco and evaluation of their antimicrobial activity acting alone or in combined processes for food preservation. J. Sci. Food. Agric. 2014, 94, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Basak, S.S.; Candan, F. Effect of Laurus nobilis L. Essential Oil and its Main Components on a-glucosidase and Reactive Oxygen Species Scavenging Activity. Iran J. Pharm. Res. 2013, 12, 367–379. [Google Scholar]
- Cao, L.; Si, J.Y.; Liu, Y.; Sun, H.; Jin, W.; Li, Z.; Zhao, X.H.; Pan, R.L. Essential oil composition, antimicrobial and antioxidant properties of Mosla chinensis Maxim. Food Chem. 2009, 115, 801–805. [Google Scholar] [CrossRef]
- Ramadan, M.M.; Yehia, H.A.; Shaheen, M.S.; Abed EL-FattahAm-Euras, M.Sh. Aroma Volatiles, Antibacterial, Antifungal and Antioxidant Properties of Essential Oils Obtained from Some Spices Widely Consumed in Egypt. American-Eurasian. J. Agric. Environ. Sci. 2014, 14, 486–494. [Google Scholar]
- Przygodzka, M.; Zielińska, D.; Ciesarová, Z.; Kukurová, K.; Zielińska, H. Comparison of methods for evaluation of the antioxidant capacity and phenolic compounds in common spices. LWT-Food Sci. Technol. 2013, 33, 1–6. [Google Scholar] [CrossRef]
- Bellik, Y.; Dahmani, Z.; Benabdesselam, F.; Boukraa, L.; Iguer-Ouada, M.; Ayad, A.; Nemmar, A. Antioxidant activity of the essential oil and oleoresin of Zingiber officinale Roscoe as affected by chemical environment. Int. J. Food Prop. 2013, 16, 1304–1313. [Google Scholar] [CrossRef]
- Rehman, Z.; Salariya, A.M.; Farzana, H. Antioxidant activity of ginger extract in sunflower oil. J. Sci. Agric. 2003, 83, 624–629. [Google Scholar] [CrossRef]
- Embuscado, M.E. Spices and herbs: Natural sources of antioxidants—A mini review. J. Funct. Foods 2015, 18, 811–819. [Google Scholar] [CrossRef]
- Sharififar, F.; Moshafi, M.H.; Mansouri, S.H.; Khodashenas, M.; Khoshnoodi, M. In vitro evaluation of antibacterial and antioxidant activities of the essential oil and methanol extract of endemic Zataria multiflora Boiss. Food Control 2007, 18, 800–805. [Google Scholar] [CrossRef]
- Dadalioglu, I.; Evrendilek, G.A. Chemical compositions and antibacterial effects of essential oils of Turkish oregano (Origanum minutiflorum), bay laurel (Laurus nobilis), Spanish lavender (Lavandula stoechas L.), and fennel (Foeniculum vulgare) on common foodborne pathogens. J. Agric. Food. Chem. 2004, 52, 8255–8260. [Google Scholar] [CrossRef] [PubMed]
- Lopez, P.; Sánchez, C.; Battle, R.; Neriän, C. Solid- and vapor-phase antimicrobial activities of six essential oils: susceptibility of selected foodborne bacterial and fungal strains. J. Agric. Food. Chem. 2005, 53, 6939–6946. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Maurya, S.; de Lampasona, M.P.; Catalan, C. Chemical constituents, antimicrobial investigations, and antioxidative potentials of Anethum graveolens L. essential oil and acetone extract: Part 52. J. Food. Sci. 2006, 70, 208–215. [Google Scholar] [CrossRef]
- Karuppiah, P.; Rajaram, S. Antibacterial effect of Allium sativum cloves and Zingiber officinale rhizomes against multiple-drug resistant clinical pathogens. Asian Pac. J. Trop. Biomed. 2012, 2, 597–601. [Google Scholar] [CrossRef]
- AnbuJebaSunilson, J.; Suraj, R.; Rejitha, G.; Anandarajagopa, K. In vitro antibacterial evaluation of Zingiber officinale, Curcuma longa and Alpinia galangal extracts as natural foods preservatives. Am. J. Food. Technol. 2009, 4, 192–200. [Google Scholar]
- Mesomo, M.C.; Corazza, M.L.; Ndiaye, P.M.; Dalla Santa, O.R.; Cardozo, L.; Sheer, A.D.P. Supercritical CO2 extracts and essential oil of ginger (Zingiber officinale R). Chemical composition and antibacterial activity. J. Supercrit. Fluids 2013, 80, 44–49. [Google Scholar] [CrossRef]
- Silveira, S.M.; Júnior, A.C.; Scheuermann, G.N.; Secchi, F.L.; Vieira, C.R.W. Chemical composition and antimicrobial activity of essential oils from selected herbs cultivated in the South of Brazil against food spoilage and foodborne pathogens. Ciênc. Rural 2012, 42, 1300–1306. [Google Scholar] [CrossRef]
- Erkmen, O.; Özcan, M.M. Antimicrobial effects of Turkish propolis, pollen, and laurel on spoilage and pathogenic food-related microorganisms. J. Med. Food 2008, 11, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Bouzouita, N.; Kachouri, F.; Hamdi, M.; Chaabouni, M.M. Antimicrobial activity of essential oils from Tunisian aromatic plants. Flavour Fragr. J. 2003, 18, 380–383. [Google Scholar] [CrossRef]
- Hamad, A.; Alifah, A.; Permadi, A.; Hartanti, D. Chemical constituents and antibacterial activities of crude extract and essential oils of Alpinia galanga and Zingiber officinale. IFRJ 2016, 23, 837–841. [Google Scholar]
- Aligiannis, N.; Kalpotzakis, E.; Mitaku, S.; Chinou, I.B. Composition and antimicrobial activity of the essential oil of two Origanum species. J. Agric. Food Chem. 2001, 40, 4168–4170. [Google Scholar] [CrossRef]
- Ruangamnart, A.; Buranaphalin, S.; Temsiririrkkul, R.; Chuakul, W.; Pratuangdejkul, J. Chemical compositions and antibacterial activity of essential oil from dill fruits (Anethum graveolens L.) cultivated in Thailand. Mahidol. Univ. J. Pharm. Sci. 2015, 42, 135–143. [Google Scholar]
- Snoussi, M.; Chaieb, K.; Rouabhia, M.; Bakhrouf, A. Quantitative study, identification and antibiotics sensitivity of some Vibrionaceae associated to a marine fish hatchery. Ann. Microbiol. 2006, 5, 289–293. [Google Scholar] [CrossRef]
- Toledo-Arana, A.; Valle, J.; Solano, C.; Arrizubieta, M.J.; Cucarella, C.; Lamata, C.; Amorena, B.; Leiva, J.; Penadés, J.R.; Lasa, I. The enterococcal surface protein, Esp, is involved in Enterococcus faecalis biofilm formation. Appl. Environ. Microbiol. 2001, 67, 4538–4545. [Google Scholar] [CrossRef] [PubMed]
- European Pharmacopoeia, 5th ed.; Council of Europe: Strasbourg Cedex, France, 2004; Volume 1, pp. 217–218.
- Jennings, W.; Shibamoto, T. Qualitative Analysis of Flavour and Fragrance Volatiles by Glass capillary Gas Chromatography; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicone and Carbowax 20M phases. J. Chromatogr. 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Goodner, K.L. Practical retention index models of OV-101, DB-1, DB-5, and DB-Wax for flavor and fragrance compounds. LWT-Food Sci. Technol. 2008, 41, 951–958. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Wiley Registry of Mass Spectral Data, with NIST Spectral Data CD Rom, 7th ed.; John Wiley & Sons: New York, NY, USA, 1998.
- Hatano, T.; Kagawa, H.; Yasuhara, T.; Okuda, T. Two new flavonoids and other constituents in licore root: Their relative astringency and radical scavenging affects. Chem. Pharm. Bull. 1988, 36, 1090–1097. [Google Scholar]
- Duh, P.D.; Tu, Y.Y.; Yen, G.C. Antioxidant activity of water extract of HarngJyur (Chrysanthemum morifolium Ramat). LWT-Food Sci. Technol. 1999, 32, 269–277. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on product of browning reaction prepared from glucose amine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Hajlaoui, H.; Mighri, H.; Noumi, E.; Snoussi, M.; Trabelsi, N.; Ksouri, R.; Bakhrouf, A. Chemical composition and biological activities of Tunisian Cuminum cyminum L. essential oil: A high effectiveness against Vibrio spp. strains. Food Chem. Toxicol. 2010, 48, 2186–2192. [Google Scholar] [CrossRef] [PubMed]
- Snoussi, M.; Hajlaoui, H.; Noumi, E.; Usai, D.; Sechi, L.A.; Zanetti, S.; Bakhrouf, A. In vitro anti-Vibrio spp. activity and chemical composition of some Tunisian aromatic plants. World J. Microbiol. Biotechnol. 2008, 24, 3071–3076. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [PubMed]
- Sample Availability: Samples of the essential oils compounds are available from the authors.


{kind=link}
| Samples | Code | Identification | Biotype on Api | % id | Exoenzymes Profile | Adhesive Properties | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Haemolysis | Caseinase | DNase | Amylase | Lecithinase | Slime (CRA) | OD595 ± SD | |||||
| P. longirostris | R58 | Klebsiella oxytoca | 5245573 | 93.6 | γ | + | − | + | − | Red | 0.28 ± 0.02 |
| R44 | Aeromonas hydrophila | 7772755 | 98.2 | γ | + | + | + | + | Black | 0.26 ± 0.06 | |
| R63 | Vibrio alginolyticus | 7650745 | 82.2 | + | + | + | − | Pink | 0.21 ± 0.02 | ||
| R72 | Aeromonas hydrophila | 5652355 | 88.2 | + | + | + | − | Bordeaux | 0.41 ± 0.03 | ||
| R32 | Staphylococcus lugdunensis | 6314150 | 42.6 | + | + | + | − | Pink | 0.21 ± 0.09 | ||
| R53 | Staphylococcus sciuri | 6314050 | 72.5 | + | + | + | − | Pink | 0.19 ± 0.01 | ||
| R30 | Staphylococcus lentus | 6731771 | 99.6 | + | + | + | − | Black | 0.31 ± 0.04 | ||
| R64 | Staphylococcus xylosus | 6735753 | 99 | + | + | + | − | Red | 0.18 ± 0.02 | ||
| D. labrax | R14 | Aeromonas hydrophila | 7576755 | 83.8 | + | + | + | + | Bordeaux | 0.27 ± 0.02 | |
| R35 | Staphylococcus sciuri | 6314050 | 72.5 | + | + | + | − | Pink | 0.19 ± 0.04 | ||
| R3 | Aeromonas hydrophila | 5567745 | 82 | γ | + | − | + | − | Red | 0.22 ± 0.01 | |
| R23 | Aeromonas hydrophila | 5777745 | 99.5 | γ | + | − | + | − | Pink | 0.21 ± 0.02 | |
| R2 | Staphylococcus sciuri | 6334070 | 99 | + | + | + | − | Red | 0.28 ± 0.12 | ||
| R71 | Aeromonas hydrophila | 7535755 | 99.9 | + | + | + | + | Pink | 0.15 ± 0.01 | ||
| R16 | Enterobacter cloacae | 5245573 | 38.4 | γ | + | − | + | − | Pink | 0.26 ± 0.05 | |
| M. galloprovincialis | R5 | Aeromonas hydrophila | 7577754 | 99.9 | + | − | + | + | Bordeaux | 0.18 ± 0.04 | |
| R29 | Aeromonas hydrophila | 7577776 | 99.9 | + | + | + | − | Red | 0.19 ± 0.03 | ||
| R70 | Aeromonas hydrophila | 7577755 | 99.3 | + | − | + | + | Bordeaux | 0.36 ± 0.09 | ||
| R47 | Aeromonas hydrophila | 7573744 | 99.8 | + | + | + | + | Bordeaux | 0.20 ± 0.01 | ||
| R22 | Aeromonas hydrophila | 5677254 | 99.2 | + | + | + | − | Black | 0.20 ± 0.01 | ||
| R13 | Aeromonas hydrophila | 7477754 | 99.9 | + | + | + | − | Black | 0.26 ± 0.01 | ||
| S. aurata | R46 | Aeromonas hydrophila | 7777757 | 99.9 | γ | + | − | + | − | Pink | 0.19 ± 0.02 |
| R11 | Aeromonas hydrophila | 7777757 | 99.9 | + | + | + | + | Red | 0.16 ± 0.01 | ||
| R68 | Enterobacter cloacae | 5245573 | 38.4 | γ | + | − | + | − | Pink | 0.13 ± 0.01 | |
| R40 | Staphylococcus sciuri | 6314050 | 72.5 | + | + | + | − | Red | 0.22 ± 0.05 | ||
| R9 | Klebsiella ornithinolytica | 5345573 | 91 | γ | + | + | + | + | Pink | 0.19 ± 0.01 | |
| R4 | Aeromonas hydrophila | 7537745 | 99.6 | + | − | + | + | Red | 0.25 ± 0.10 | ||
| R36 | Staphylococcus sciuri | 6314050 | 72.5 | + | + | + | − | Pink | 0.27 ± 0.04 | ||
| R48 | Aeromonas hydrophila | 7577755 | 99.9 | + | + | + | + | Bordeaux | 0.20 ± 0.01 | ||
| R37 | Staphylococcus sciuri | 6314050 | 72.5 | + | + | + | − | Pink | 0.22 ± 0.05 | ||
| R18 | Aeromonas hydrophila | 7477777 | 78.1 | + | + | + | + | Pink | 0.19 ± 0.03 | ||
| R55 | Aeromonas hydrophila | 7777744 | 99.6 | + | + | + | + | Black | 0.30 ± 0.02 | ||
| R69 | Serratia odorifera | 7347573 | 95.2 | γ | + | + | + | − | Red | 0.37 ± 0.03 | |
| R1 | Staphylococcus lugdunensis | 6314150 | 42.6 | + | − | + | − | Red | 0.18 ± 0.03 | ||
| Constituents | RI | Essential Oils | Constituents | RI | Essential Oils | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ln | Ag | Zo | Ln | Ag | Zo | ||||
| (E)-3-Hexen-1-ol | 853 | 0.3 | Cumin aldehyde | 1241 | 0.2 | ||||
| 2-Heptanone | 891 | Tr | Neral | 1241 | 27.0 | 0.5 | |||
| 2-Heptanol | 901 | 0.4 | Carvone | 1244 | 25.7 | 0.3 | |||
| Tricyclene | 928 | 0.3 | Piperitone | 1254 | |||||
| α-Thujene | 933 | 0.2 | Tr | Geraniol | 1256 | 0.7 | |||
| α-Pinene | 941 | 3.8 | 0.8 | 3.0 | Geranial | 1269 | 0.6 | ||
| Camphene | 955 | 0.5 | 0.1 | 11.5 | Phellandral | 1274 | 0.4 | ||
| Sabinene | 977 | 3.5 | 0.5 | Bornyl acetate | 1287 | 0.6 | 0.9 | ||
| β-Pinene | 982 | 3.6 | 0.2 | 0.2 | 2-Undecanone | 1292 | 1.0 | ||
| 6-Methyl-5-hepten-2-one | 987 | 1.0 | α-Terpinyl acetate | 1352 | 9.0 | ||||
| Myrcene | 993 | 0.3 | 1.5 | 1.0 | α-Copaene | 1377 | 0.2 | ||
| δ-2-Carene | 1003 | 0.6 | Geranyl acetate | 1383 | 0.8 | ||||
| α-Phellandrene | 1006 | Tr | 1.4 | 2.0 | β-Elemene | 1392 | Tr | ||
| δ-3-Carene | 1013 | Tr | Methyl eugenol | 1403 | 3.6 | ||||
| α-Terpinene | 1020 | 0.3 | 0.1 | 1.4 | β-Caryophyllene | 1419 | Tr | ||
| p-Cymene | 1027 | 0.5 | 0.7 | 1.2 | Allo-Aromadendrene | 1461 | 1.1 | ||
| Limonene | 1032 | 0.7 | 20.6 | β-Chamigrene | 1477 | 0.4 | |||
| β-Phellandrene | 1033 | 10.7 | γ-Curcumene | 1481 | 1.1 | ||||
| 1.8-Cineole | 1034 | 56.0 | 10.4 | ar-Curcumene | 1483 | 4.6 | |||
| γ-Terpinene | 1063 | 0.6 | 0.2 | Tr | Valencene | 1493 | 0.4 | ||
| cis-Sabinene hydrate | 1070 | 0.2 | Tr | α-Zingiberene | 1495 | 6.9 | |||
| cis-Linalool oxide (furanoid) | 1077 | Tr | 0.2 | β-Bisabolene | 1509 | 3.2 | |||
| Terpinolene | 1090 | 0.2 | 0.2 | β-Sesquiphellandrene | 1524 | 4.1 | |||
| p-Cymenene | 1091 | 0.7 | Selina-3.7(11)-diene | 1544 | 0.2 | ||||
| 2-Nonanone | 1092 | 0.4 | Elemol | 1549 | 0.2 | ||||
| Linalool | 1101 | 3.8 | 1.9 | (E)-Nerolidol | 1565 | 0.5 | |||
| exo-Fenchol | 1118 | 0.4 | Spathulenol | 1577 | 0.4 | ||||
| cis-p-Menth-2-en-1-ol | 1123 | Tr | 0.1 | 0.3 | Caryophyllene oxide | 1582 | 0.3 | ||
| trans-Pinocarveol | 1141 | 0.3 | 10-epi-γ-Eudesmol | 1622 | 2.5 | ||||
| trans-p-Menth-2-en-1-ol | 1142 | 0.1 | 0.2 | Dill apiole | 1623 | 8.0 | |||
| cis-Verbenol | 1142 | Tr | γ-Eudesmol | 1631 | 0.8 | ||||
| Camphor | 1145 | 4.4 | 0.5 | β-Eudesmol | 1650 | 0.8 | |||
| Camphene hydrate | 1150 | 0.8 | α-Eudesmol | 1653 | 0.5 | ||||
| Isoborneol | 1158 | 0.5 | α-Cadinol | 1654 | 1.0 | ||||
| Borneol | 1168 | Tr | 6.4 | β-Bisabolol | 1672 | 0.7 | |||
| 4-Terpineol | 1178 | 5.2 | 0.3 | 1.0 | α-Bisabolol | 1684 | 0.2 | ||
| p-Cymen-8-ol | 1185 | 0.4 | Monoterpene hydrocarbons | 26.9 | 14.2 | 32.0 | |||
| Dill ether | 1186 | 0.6 | Oxygenated monoterpenes | 64.8 | 79.8 | 31.0 | |||
| α-Terpineol | 1190 | 4.7 | 3.2 | Sesquiterpene hydrocarbons | 0 | 0 | 22.2 | ||
| Myrtenol | 1195 | 0.2 | Oxygenated sesquiterpenes | 0 | 0.7 | 7.2 | |||
| cis-Dihydrocarvone | 1195 | 1.7 | Phenylpropenoids | 8.0 | 3.6 | 0 | |||
| trans-Dihydrocarvone | 1202 | 4.9 | Others | 0 | 0.3 | 2.8 | |||
| trans-Piperitol | 1207 | 0.2 | Total identified | 99.7 | 98.6 | 95.2 | |||
| Activities | Essential Oils | Standards | ||||
|---|---|---|---|---|---|---|
| L. nobilis | Z. officinale | A. graveolens | BHT | Ascorbic Acid | BHA | |
| DPPH (IC50; µg/mL) | 135 | 470 | 3000 | 11.5 | ||
| RP (EC50; µg/mL) | 1850 | 1900 | 2400 | 37.3 | ||
| β-carotenes (µg/mL) | 3600 | 1900 | 4000 | 48 | ||
| Ionsuperoxyde (O2−) (µg/mL) | 620 | 320 | 400 | 1.5 | ||
| Species | Code | Essential Oils | Antibiotics | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L. nobilis | Z. officinale | A. graveolens | ||||||||||
| ZI ± SD | MIC | MBC | ZI ± SD | MIC | MBC | ZI ± SD | MIC | MBC | 1 | 2 | ||
| A. hydrophila | R4 | 12.33 ± 0.58 hi | 0.05 | >12.5 | 8.67 ± 0.58 ij | 0.05 | >12.5 | 11.33 ± 0.58 ghi | 0.05 | 50 | 8 | 6 |
| A. hydrophila | R47 | 20 ± 1 b | 0.05 | 25 | 13.67 ± 0.58 cd | 0.05 | >0.78 | 15.33 ± 0.58 d | 0.05 | >25 | 15 | 7 |
| A. hydrophila | R48 | 22.33 ± 0.58 a | 0.10 | 50 | 14.33 ± 0.58 bc | 0.20 | 12.5 | 11.67 ± 0.58 gh | 0.10 | 50 | 10 | 6 |
| A. hydrophila | R71 | 16.67 ± 0.58 c | 0.20 | >25 | 13.67 ± 0.58 cd | 0.05 | >25 | 13 ± 1 ef | 0.20 | >12.5 | 15 | 6 |
| A. hydrophila | R29 | 14.33 ± 0.58 efg | 0.10 | >6.25 | 12.33 ± 0.58 efg | 0.10 | >25 | 13 ± 0 ef | 0.10 | 50 | 6 | 20 |
| A. hydrophila | R5 | 20.67 ± 0.58 b | 0.10 | 12.5 | 12.67 ± 0.58 ef | 0.10 | 12.5 | 25.33 ± 0.58 a | 0.10 | 50 | 11 | 8 |
| A. hydrophila | R13 | 11.33 ± 0.58 j | 0.10 | >12.5 | 7 ± 0 k | 0.05 | >25 | 6 ± 0 l | 0.10 | >12 | 13 | 6 |
| A. hydrophila | R3 | 15.33 ± 0.58 d | 0.10 | >25 | 15.33 ± 0.58 a | 0.10 | >12.5 | 13.33 ± 0.58 e | 0.10 | >12.5 | 19 | 6 |
| A. hydrophila | R46 | 14.33 ± 0.58 def | 0.10 | 25 | 14.33 ± 0.58 bc | 0.05 | >25 | 12.33 ± 0.58 efg | 0.10 | >12.5 | 11 | 7 |
| A. hydrophila | R22 | 11.67 ± 0.58 ij | 0.05 | 6.25 | 8.67 ± 0.58 ij | 0.05 | 12.5 | 7.33 ± 0.58 k | 0.10 | >12.5 | 11 | 6 |
| A. hydrophila | R14 | 13.67 ± 0.58 g | 0.05 | >6.25 | 10.33 ± 0.58 h | 0.05 | >25 | 8.67 ± 0.58 j | 0.05 | >25 | 14 | 6 |
| A. hydrophila | R70 | 11.67 ± 0.58 ij | 0.20 | >25 | 11.67 ± 0.58 fg | 0.10 | 50 | 11.33 ± 0.58 ghi | 0.05 | >12.5 | 7 | 6 |
| A. hydrophila | R11 | 14.67 ± 0.58 de | 0.10 | >25 | 12.67 ± 0.58 ef | 0.05 | >6.25 | 14.67 ± 0.58 d | 0.05 | >12.5 | 10 | 7 |
| A. hydrophila | R55 | 12 ± 1 ij | 0.05 | 12.5 | 12.00 ± 0 fg | 0.05 | >3.13 | 12 ± 1 fgh | 0.10 | 12.5 | 10 | 6 |
| A. hydrophila | R18 | 13.67 ± 0.58 efg | 0.05 | 12.5 | 12.67 ± 0.58 ef | 0.05 | 50 | 11 ± 0 hi | 0.05 | >25 | 11 | 7 |
| A. hydrophila | R23 | 14.67 ± 0.58 de | 0.05 | >6.25 | 12.33 ± 0.58 efg | 0.05 | 50 | 13 ± 0 ef | 0.10 | >25 | 12 | 6 |
| A. hydrophila | R44 | 13.33 ± 0.58 fg | 0.10 | >25 | 6 ± 0 l | 0.05 | 50 | 6 ± 0 l | 0.05 | >12.5 | 12 | 6 |
| A. hydrophila | R72 | 15.33 ± 0.58 d | 0.05 | 12.5 | 12.33 ± 0.58 efg | 0.05 | >0.78 | 12.33 ± 0.58 efg | 0.05 | >25 | 11 | 6 |
| E. cloacae | R68 | 10.33 ± 0.58 k | 0.20 | >25 | 9 ± 0 i | 0.05 | >25 | 11.67 ± 0.58 gh | 0.05 | >25 | 8 | 8 |
| E. cloacae | R16 | 11.67 ± 0.58 ij | 0.05 | >50 | 8 ± 0 j | 0.10 | >25 | 7.67 ± 0.58 k | 0.10 | 50 | 12 | 7 |
| K. ornithinolytica | R9 | 17 ± 0 c | 0.05 | 6.25 | 11.33 ± 0.58 g | 0.20 | >25 | 8.67 ± 0.58 j | 0.05 | >25 | 15 | 7 |
| K. oxytoca | R58 | 14.67 ± 0.58 de | 0.05 | >12.5 | 13 ± 0 de | 0.05 | >12.5 | 12 ± 0 fgh | 0.10 | 50 | 10 | 11 |
| S. lentus | R30 | 6 ± 0 l | 0.05 | >6.25 | 6 ± 0 l | 0.05 | >3.13 | 11.33 ± 0.58 ghi | 0.05 | >25 | 22 | 6 |
| S. lugdunensis | R1 | 14.33 ± 0.58 def | 0.10 | >25 | 10.33 ± 0.58 h | 0.05 | >12.5 | 10.33 ± 0.58 i | 0.10 | >25 | 19 | 6 |
| S. lugdunensis | R32 | 15.33 ± 0.58 d | 0.05 | >12.5 | 14.67 ± 0.58 ab | 0.20 | >12.5 | 17.67 ± 0.58 c | 0.39 | 50 | 22 | 20 |
| S. odorifera | R69 | 14.33 ± 0.58 def | 0.10 | >12.5 | 11.33 ± 0.58 g | 0.05 | >12.5 | 11.33 ± 0.58 ghi | 0.10 | >12.5 | 8 | 7 |
| S. sciuri | R40 | 14.67 ± 0.58 de | 0.05 | >25 | 11.33 ± 0.58 g | 0.10 | >25 | 11.33 ± 0.58 ghi | 0.10 | 50 | 15 | 7 |
| S. sciuri | R35 | 14.67 ± 0.58 de | 0.05 | >25 | 11.67 ± 0.58 fg | 0.05 | >25 | 13.33 ± 0.58 e | 0.10 | >25 | 24 | 6 |
| S. sciuri | R36 | 14.33 ± 0.58 def | 0.05 | >6.25 | 12.33 ± 0.58 efg | 0.20 | >6.25 | 14.67 ± 0.58 d | 0.05 | >25 | 10 | 13 |
| S. sciuri | R37 | 14.67 ± 0.58 de | 0.05 | >25 | 8.67 ± 0.58 ij | 0.05 | >6.25 | 11.67 ± 0.58 gh | 0.10 | >25 | 7 | 15 |
| S. sciuri | R2 | 14 ± 1 ef | 0.05 | >12.5 | 11.67 ± 0.58 fg | 0.10 | >12.5 | 11.67 ± 0.58 gh | 0.10 | 50 | 8 | 20 |
| S. sciuri | R53 | 14.67 ± 0.58 de | 0.05 | 12.5 | 13.67 ± 0.58 cd | 0.05 | 1.56 | 20 ± 1 b | 0.05 | >12.5 | 10 | 7 |
| S. xylosus | R64 | 14.33 ± 0.58 def | 0.05 | >6.25 | 12.33 ± 0.58 efg | 0.05 | >12.5 | 15.33 ± 0.58 d | 0.10 | >12.5 | 12 | 8 |
| V. alginolyticus | R63 | 13 ± 0 gh | 0.05 | >6.25 | 10.33 ± 0.58 h | 0.05 | >25 | 12.33 ± 0.58 efg | 0.10 | 50 | 8 | 7 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snuossi, M.; Trabelsi, N.; Ben Taleb, S.; Dehmeni, A.; Flamini, G.; De Feo, V. Laurus nobilis, Zingiber officinale and Anethum graveolens Essential Oils: Composition, Antioxidant and Antibacterial Activities against Bacteria Isolated from Fish and Shellfish. Molecules 2016, 21, 1414. https://doi.org/10.3390/molecules21101414
Snuossi M, Trabelsi N, Ben Taleb S, Dehmeni A, Flamini G, De Feo V. Laurus nobilis, Zingiber officinale and Anethum graveolens Essential Oils: Composition, Antioxidant and Antibacterial Activities against Bacteria Isolated from Fish and Shellfish. Molecules. 2016; 21(10):1414. https://doi.org/10.3390/molecules21101414
Chicago/Turabian StyleSnuossi, Mejdi, Najla Trabelsi, Sabrine Ben Taleb, Ameni Dehmeni, Guido Flamini, and Vincenzo De Feo. 2016. "Laurus nobilis, Zingiber officinale and Anethum graveolens Essential Oils: Composition, Antioxidant and Antibacterial Activities against Bacteria Isolated from Fish and Shellfish" Molecules 21, no. 10: 1414. https://doi.org/10.3390/molecules21101414
APA StyleSnuossi, M., Trabelsi, N., Ben Taleb, S., Dehmeni, A., Flamini, G., & De Feo, V. (2016). Laurus nobilis, Zingiber officinale and Anethum graveolens Essential Oils: Composition, Antioxidant and Antibacterial Activities against Bacteria Isolated from Fish and Shellfish. Molecules, 21(10), 1414. https://doi.org/10.3390/molecules21101414