
Effects of Resveratrol Supplementation and Exercise Training on Exercise Performance in Middle-Aged Mice


Abstract
:1. Introduction
2. Results
2.1. Effect of 4-Week RES Supplementation and ET on Body Weight, Food Intake, Water Intake, and Tissue Changes
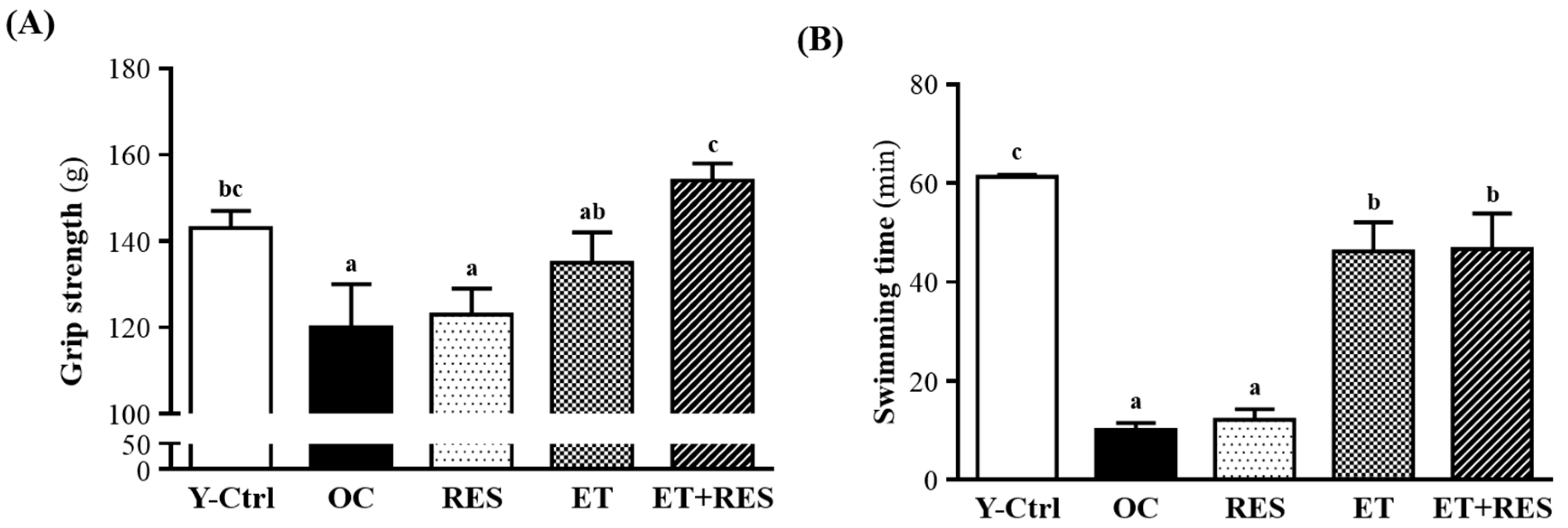
2.2. Effect of 4-Week RES Supplementation and ET on Forelimb Grip Strength
2.3. Effect of 4-Week RES Supplementation and ET on an Exhaustive Swimming Test
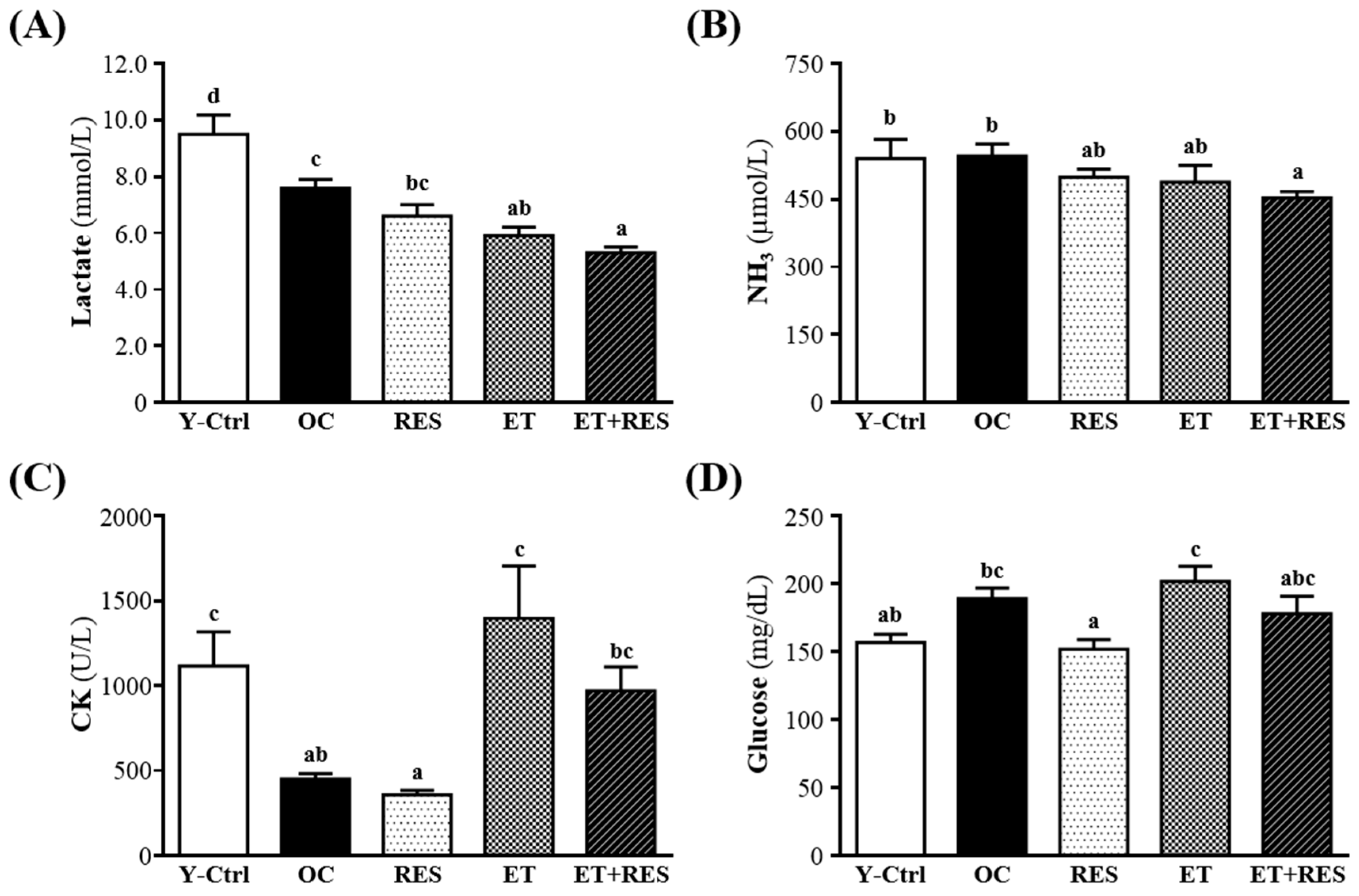
2.4. Effect of 4-Week RES Supplementation and ET on Lactate, Ammonia, CK, and Glucose after a 15-min Swimming Test
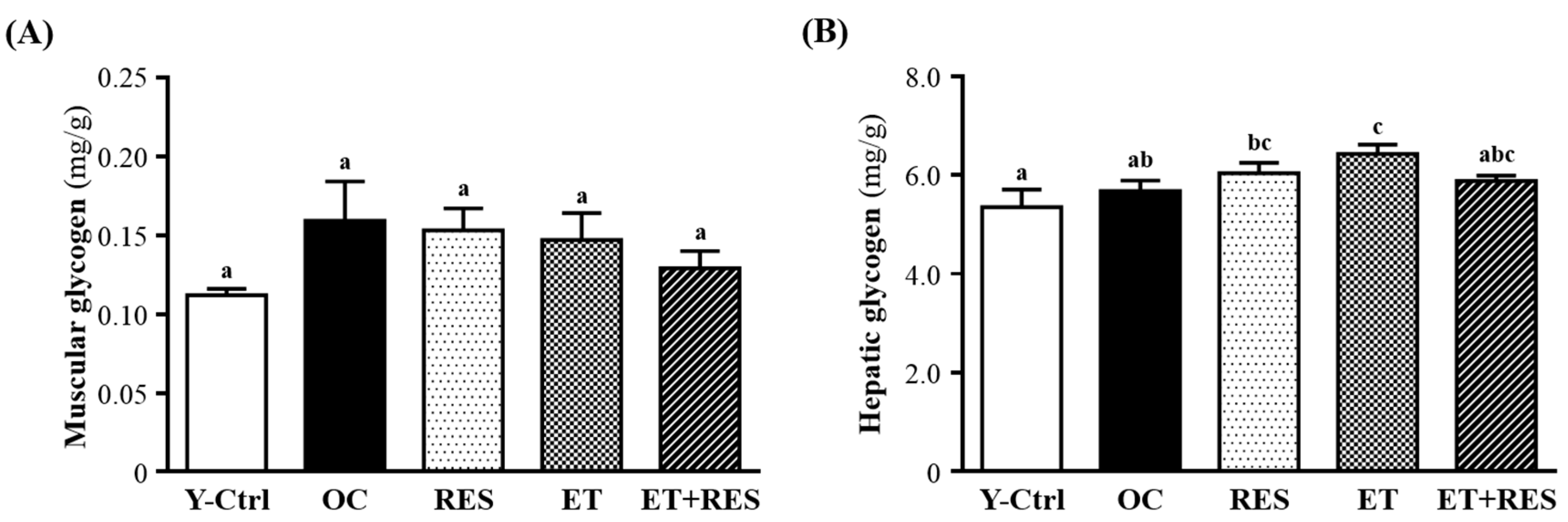
2.5. Effect of 4-Week RES Supplementation and ET on Biochemical Assessments
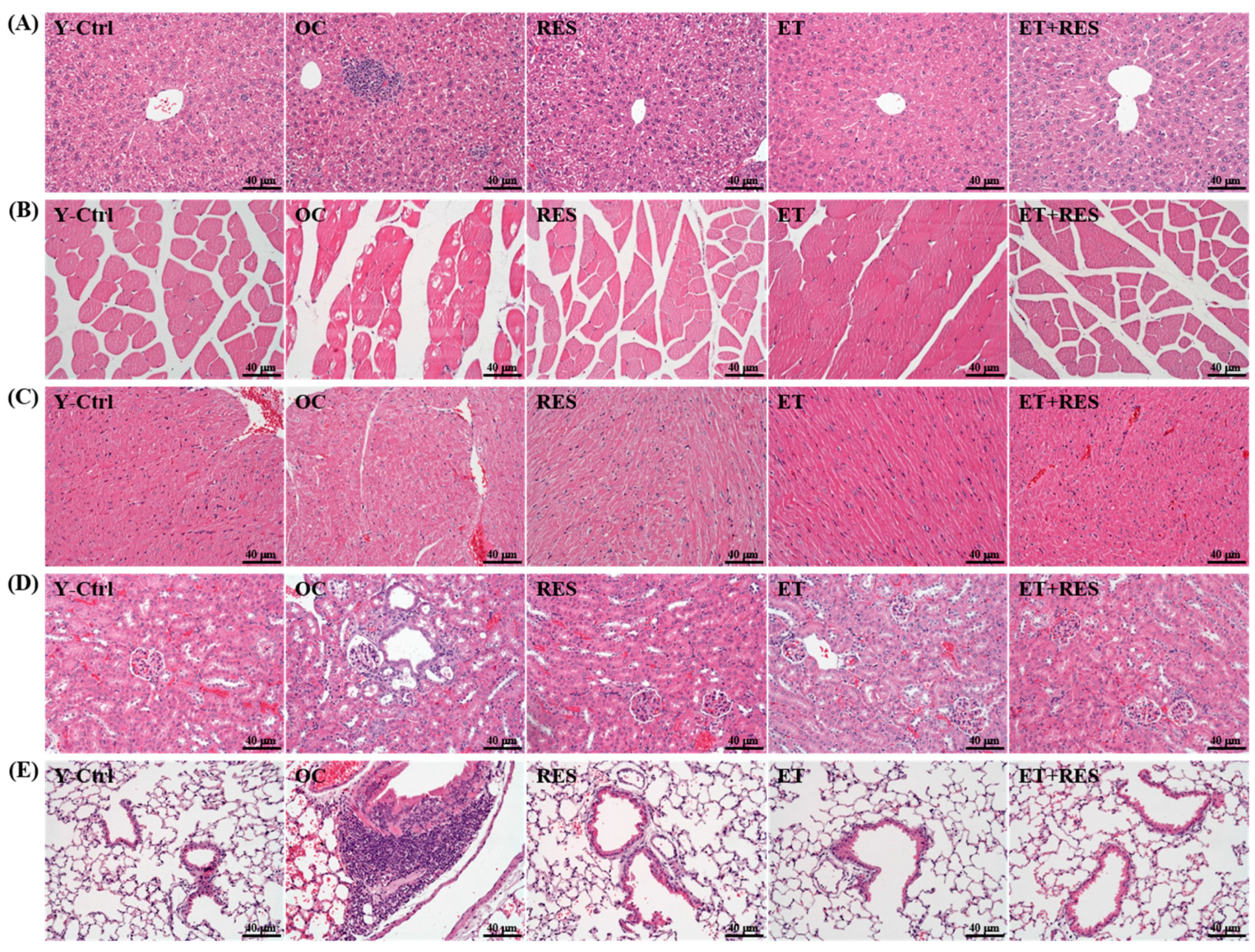
2.6. Effect of 4-Week RES Supplementation and ET on Histopathological Evaluation of Tissues
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals and Treatment
4.3. Forelimb Grip Strength
4.4. Exhaustive Swimming Test
4.5. Fatigue-Associated Biochemical Indices
4.6. Blood Biochemical Assessments
4.7. Tissue Glycogen Determination
4.8. Histological Staining of Tissues
4.9. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tanaka, M.; Baba, Y.; Kataoka, Y.; Kinbara, N.; Sagesaka, Y.M.; Kakuda, T.; Watanabe, Y. Effects of (−)-epigallocatechin gallate in liver of an animal model of combined (physical and mental) fatigue. Nutrition 2008, 24, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, H.L.; Lu, R.; Zhou, Y.J.; Ma, R.; Lv, J.Q.; Li, X.L.; Chen, L.J.; Yao, Z. The decapeptide CMS001 enhances swimming endurance in mice. Peptides 2008, 29, 1176–1182. [Google Scholar] [CrossRef] [PubMed]
- Yue, Z.; Xiuhong, Z.; Shuyan, Y.; Zhonghua, Z. Effect of inonotus obliquus polysaccharides on physical fatigue in mice. J. Tradit. Chin. Med. 2015, 35, 468–472. [Google Scholar] [PubMed]
- De Angelis, K.; Schaan, B.D.; Maeda, C.Y.; Dall’Ago, P.; Wichi, R.B.; Irigoyen, M.C. Cardiovascular control in experimental diabetes. Braz. J. Med. Biol. Res. 2002, 35, 1091–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Pulinilkunnil, T.; Yuen, G.; Kewalramani, G.; An, D.; Qi, D.; Abrahani, A.; Rodrigues, B. Cardiomyocyte apoptosis induced by short-term diabetes requires mitochondrial GSH depletion. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H768–H776. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B.; Manson, J.E.; Stampfer, M.J.; Colditz, G.; Liu, S.; Solomon, C.G.; Willett, W.C. Diet, lifestyle, and the risk of type 2 diabetes mellitus in women. N. Engl. J. Med. 2001, 345, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Anitha, V.; Asha Devi, S. Age-related responses of right ventricle in swim-trained rats: Changes in lactate and pyruvate contents and lactate dehydrogenase activity. Mech. Ageing Dev. 1996, 90, 91–102. [Google Scholar] [CrossRef]
- Kuo, S.M. Dietary flavonoid and cancer prevention: Evidence and potential mechanism. Crit. Rev. Oncog. 1997, 8, 47–69. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Hsu, M.C.; Huang, W.C.; Yang, H.R.; Hou, C.C. Triterpenoid-rich extract from Antrodia camphorata improves physical fatigue and exercise performance in mice. Evid. Based Complement. Altern. Med. 2012, 2012, 364741. [Google Scholar] [CrossRef] [PubMed]
- Olas, B.; Wachowicz, B. Resveratrol and vitamin C as antioxidants in blood platelets. Thromb. Res. 2002, 106, 143–148. [Google Scholar] [CrossRef]
- Liu, J.C.; Chen, J.J.; Chan, P.; Cheng, C.F.; Cheng, T.H. Inhibition of cyclic strain-induced endothelin-1 gene expression by resveratrol. Hypertension 2003, 42, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.E.; Huang, W.C.; Liao, C.C.; Chang, Y.K.; Kan, N.W.; Huang, C.C. Resveratrol protects against physical fatigue and improves exercise performance in mice. Molecules 2013, 18, 4689–4702. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Huang, W.C.; Chiu, C.C.; Chang, Y.K.; Huang, C.C. Whey protein improves exercise performance and biochemical profiles in trained mice. Med. Sci. Sports Exerc. 2014, 46, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Dal-Pan, A.; Pifferi, F.; Marchal, J.; Picq, J.L.; Aujard, F. RESTRIKAL Consortium. Cognitive performances are selectively enhanced during chronic caloric restriction or resveratrol supplementation in a primate. PLoS ONE 2011, 6, e16581. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yao, X.B.; Bao, B.L.; Zhang, Y. Anti-fatigue activity of a triterpenoid-rich extract from Chinese bamboo shavings (Caulis bamfusae in taeniam). Phytother. Res. 2006, 20, 872–876. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Lu, Z.X.; Bie, X.M.; Lu, F.X.; Huang, X.Q. Scavenging and anti-fatigue activity of fermented defatted soybean peptides. Eur. Food Res. Technol. 2008, 226, 415–421. [Google Scholar] [CrossRef]
- Li, M.; Donglian, C.; Huaixing, L.; Bende, T.; Lihua, S.; Ying, W. Anti-fatigue effects of salidroside in mice. J. Med. Coll. PLA 2008, 23, 88–93. [Google Scholar] [CrossRef]
- Tashiro, S. Studies on alkali genesis in tissues: I. Ammonia production in the nerve fiber during excitation. Am. J. Physiol. 1922, 60, 519–543. [Google Scholar]
- Prado, E.S.; de Rezende Neto, J.M.; de Almeida, R.D.; Dória de Melo, M.G.; Cameron, L.C. Keto analogue and amino acid supplementation affects the ammonaemia response during exercise under ketogenic conditions. Br. J. Nutr. 2011, 105, 1729–1733. [Google Scholar] [CrossRef] [PubMed]
- Warren, G.L.; Ingalls, C.P.; Lowe, D.A.; Armstrong, R.B. Excitation-contraction uncoupling: Major role in contraction-induced muscle injury. Exerc. Sport Sci. Rev. 2001, 29, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Wagenmakers, A.J.; Beckers, E.J.; Brouns, F.; Kuipers, H.; Soeters, P.B.; van der Vusse, G.J.; Saris, W.H. Carbohydrate supplementation, glycogen depletion, and amino acid metabolism during exercise. Am. J. Physiol. 1991, 260, 883–890. [Google Scholar]
- Wang, J.J.; Shieh, M.J.; Kuo, S.L.; Lee, C.L.; Pan, T.M. Effect of red mold rice on antifatigue and exercise-related changes in lipid peroxidation in endurance exercise. Appl. Microbiol. Biotechnol. 2006, 70, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.M.; Wu, C.F. Antifatigue activity of tissue culture extracts of Saussurea involucrate. Pharm. Biol. 2008, 46, 433–436. [Google Scholar] [CrossRef]
- Schwer, B.; Verdin, E. Conserved metabolic regulatory functions of sirtuins. Cell Metab. 2008, 7, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.T.; Lerin, C.; Haas, W.; Gygi, S.P.; Spiegelman, B.M.; Puigserver, P. Nutrient control of glucose homeostasis through a complex of PGC-1α and SIRT1. Nature 2005, 434, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Niu, Q.; Li, X.; Liu, T.; Xu, Y.; Han, H.; Wang, W.; Fan, N.; Tian, Q.; Zhang, H.; Wang, Z. Long-term resveratrol consumption protects ovariectomized rats chronically treated with d-galactose from developing memory decline without effects on the uterus. Brain Res. 2012, 1467, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Busch, F.; Mobasheri, A.; Shayan, P.; Lueders, C.; Stahlmann, R.; Shakibaei, M. Resveratrol modulates interleukin-1β-induced phosphatidylinositol 3-kinase and nuclear factor κB signaling pathways in human tenocytes. J. Biol. Chem. 2012, 287, 38050–38063. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Pichardo, L.; Dharmawardhane, S.F. Grape polyphenols inhibit Akt/mammalian target of rapamycin signaling and potentiate the effects of gefitinib in breast cancer. Nutr. Cancer 2012, 64, 1058–1069. [Google Scholar] [CrossRef] [PubMed]
- Ramar, M.; Manikandan, B.; Raman, T.; Priyadarsini, A.; Palanisamy, S.; Velayudam, M.; Munusamy, A.; Marimuthu Prabhu, N.; Vaseeharan, B. Protective effect of ferulic acid and resveratrol against alloxan-induced diabetes in mice. Eur. J. Pharmacol. 2012, 690, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Hayashi, R.; Suzuki, K.; Imanishi, S.; Kambara, K.; Okazawa, S.; Inomata, M.; Yamada, T.; Yamazaki, Y.; Koshimizu, Y.; et al. The Sirt1 activator SRT1720 suppresses inflammation in an ova-induced mouse model of asthma. Respirology 2013, 18, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Falchi, M.; Bertelli, A.; Galazzo, R.; Viganò, P.; Dib, B. Central antalgic activity of resveratrol. Arch. Ital. Biol. 2010, 148, 389–396. [Google Scholar] [PubMed]
- Pedersen, B.K.; Saltin, B. Evidence for prescribing exercise as therapy in chronic disease. Scand. J. Med. Sci. Sports 2006, 16, 3–63. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Huang, W.C.; Yang, S.C.; Chan, C.C.; Lin, W.T. Ganoderma tsugae hepatoprotection against exhaustive exercise-induced liver injury in rats. Molecules 2013, 18, 1741–1754. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Lin, T.J.; Chen, C.C.; Lin, W.T. Endurance training accelerates exhaustive exercise-induced mitochondrial DNA deletion and apoptosis of left ventricle myocardium in rats. Eur. J. Appl. Physiol. 2009, 107, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Dragsted, L.O. Biomarkers of exposure to vitamins A, C, and E and their relation to lipid and protein oxidation markers. Eur. J. Nutr. 2008, 47, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.B. Initial events in exercise-induced muscular injury. Med. Sci. Sports Exerc. 1990, 22, 429–435. [Google Scholar] [PubMed]
- Nie, J.; Tong, T.K.; George, K.; Fu, F.H.; Lin, H.; Shi, Q. Resting and postexercise serum biomarkers of cardiac and skeletal muscle damage in adolescent runners. Scand. J. Med. Sci. Sports 2011, 21, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Dudley, J.I.; Das, D.K. Dose-dependency of resveratrol in providing health benefits. Dose Response 2010, 8, 478–500. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Y-Ctrl | OC | RES | ET | ET+RES |
|---|---|---|---|---|---|
| Final BW (g) | 30.30.3 | 31.70.3 | 32.10.5 | 31.10.9 | 32.30.4 |
| Diet intake (g/mouse/day) | 4.50.1 d | 4.20.1 d | 3.50.0 a | 4.00.1 c | 3.70.0 b |
| Water (g/mouse/day) | 6.10.2 a | 6.80.1 ab | 7.00.3 b | 6.90.2 b | 6.10.2 a |
| Liver (g) | 1.450.04 a | 1.420.05 a | 1.490.08 a | 1.300.09 a | 1.350.02 a |
| Kidney (g) | 0.390.02 a | 0.410.02 a | 0.440.02 a | 0.410.04 a | 0.440.01 a |
| Heart (g) | 0.240.02 b | 0.230.01 ab | 0.260.02 b | 0.180.02 a | 0.220.02 ab |
| Lung (g) | 0.350.04 a | 0.310.04 a | 0.350.03 a | 0.320.06 a | 0.330.04 a |
| Muscle (g) | 0.330.01 a | 0.290.01 a | 0.290.01 a | 0.290.02 a | 0.310.01 a |
| BAT (g) | 0.080.00 a | 0.070.01 a | 0.080.01 a | 0.100.02 a | 0.110.01 a |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kan, N.-W.; Ho, C.-S.; Chiu, Y.-S.; Huang, W.-C.; Chen, P.-Y.; Tung, Y.-T.; Huang, C.-C. Effects of Resveratrol Supplementation and Exercise Training on Exercise Performance in Middle-Aged Mice. Molecules 2016, 21, 661. https://doi.org/10.3390/molecules21050661
Kan N-W, Ho C-S, Chiu Y-S, Huang W-C, Chen P-Y, Tung Y-T, Huang C-C. Effects of Resveratrol Supplementation and Exercise Training on Exercise Performance in Middle-Aged Mice. Molecules. 2016; 21(5):661. https://doi.org/10.3390/molecules21050661
Chicago/Turabian StyleKan, Nai-Wen, Chin-Shan Ho, Yen-Shuo Chiu, Wen-Ching Huang, Pei-Yu Chen, Yu-Tang Tung, and Chi-Chang Huang. 2016. "Effects of Resveratrol Supplementation and Exercise Training on Exercise Performance in Middle-Aged Mice" Molecules 21, no. 5: 661. https://doi.org/10.3390/molecules21050661
APA StyleKan, N.-W., Ho, C.-S., Chiu, Y.-S., Huang, W.-C., Chen, P.-Y., Tung, Y.-T., & Huang, C.-C. (2016). Effects of Resveratrol Supplementation and Exercise Training on Exercise Performance in Middle-Aged Mice. Molecules, 21(5), 661. https://doi.org/10.3390/molecules21050661