Monitoring Protein Dynamics in Protein O-Mannosyltransferase Mutants In Vivo by Tandem Fluorescent Protein Timers

Abstract
:1. Introduction
2. Results
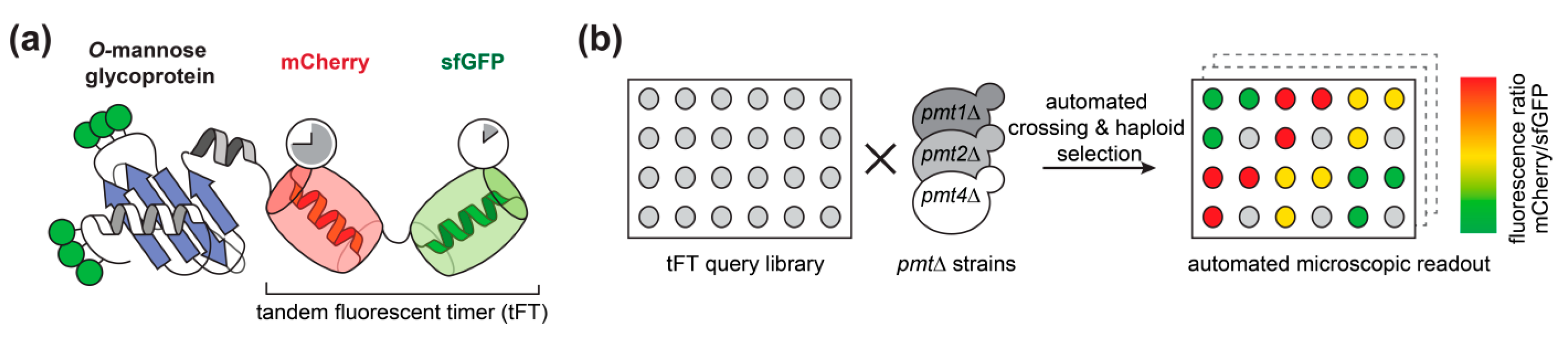
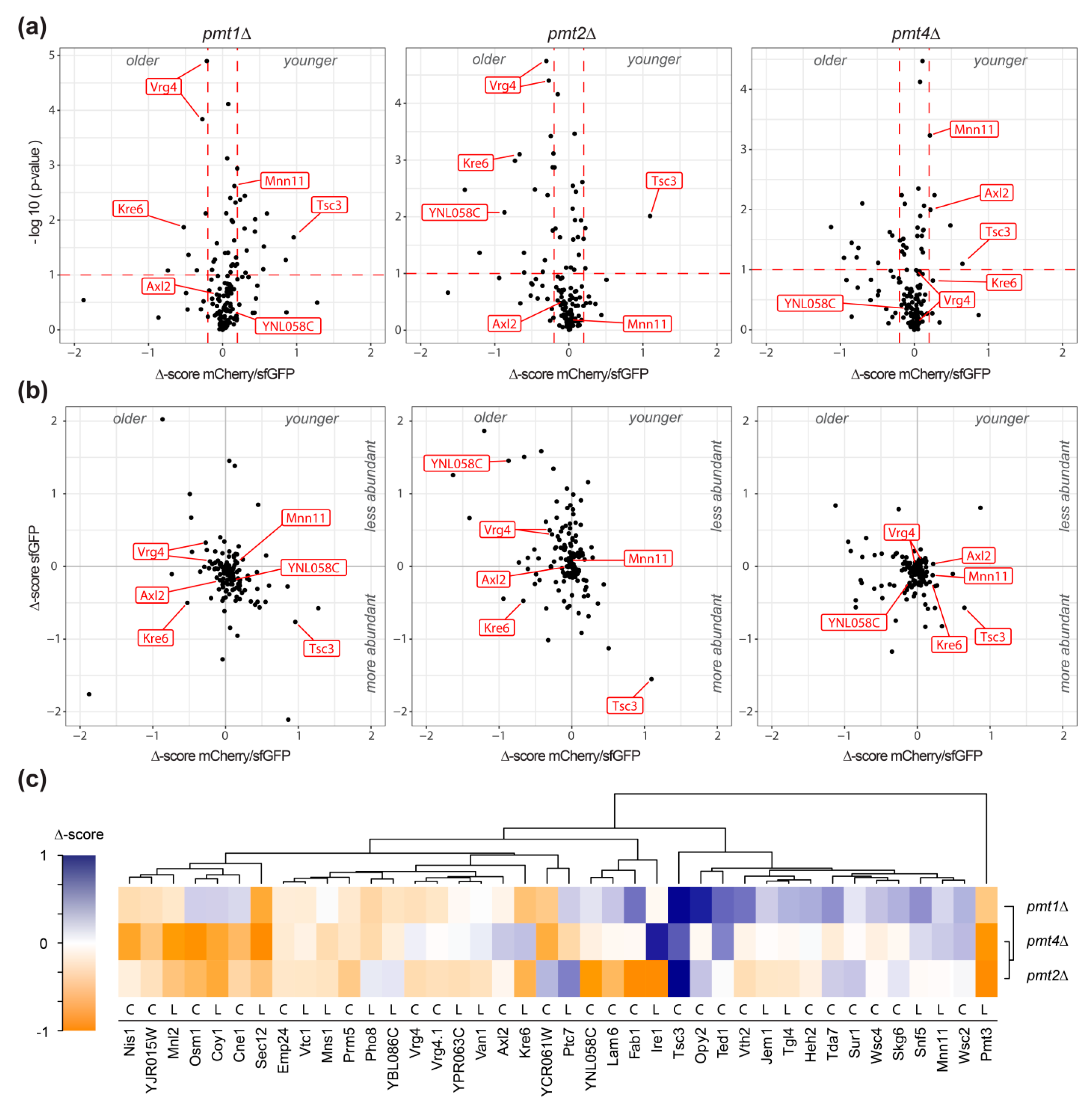
2.1. Large Scale Analysis of the Impact of O-Mannosylation on Protein Stability Using Tandem Fluorescent Protein Timers
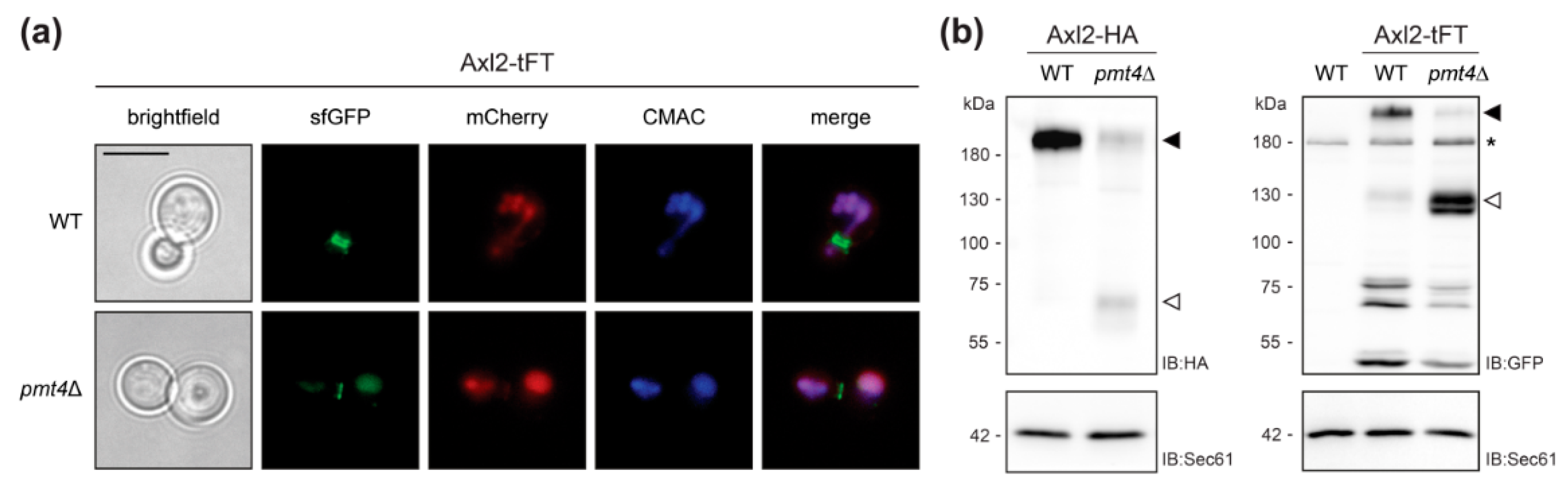
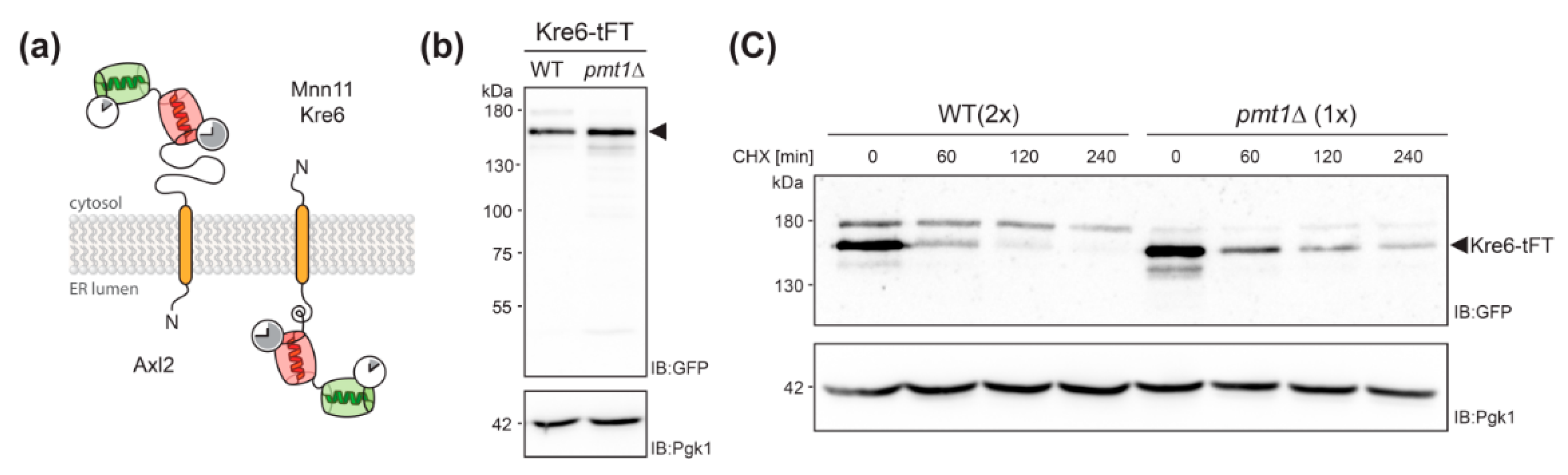
2.2. Validation of the Screening Results on the Example of Axl2
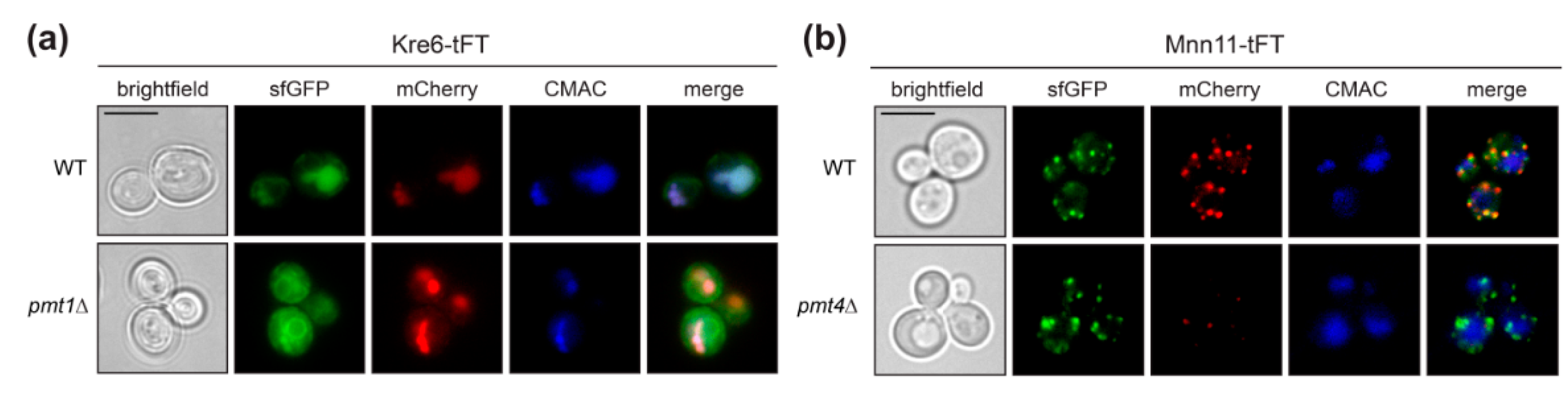
2.3. Defects in O-Mannosylation Result in Protein Stabilization
3. Discussion
4. Materials and Methods
4.1. Yeast Strains and Growth Conditions
4.2. Tandem Fluorescent Protein Timers Screening and Data Analyses
4.3. Fluorescence Flow Cytometry
4.4. Preparation of Cell Extracts And Membranes
4.5. Cycloheximide Chase Experiments
4.6. Western Blot Analyses
4.7. Microscopy
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sentandreu, R.; Northcote, D.H. The characterization of oligosaccharide attachment to threonine and serine in mannan glycopeptides obtained from the cell wall of yeast. Carbohydr. Res. 1969, 10, 584–585. [Google Scholar] [CrossRef]
- Larsen, I.S.B.; Narimatsu, Y.; Joshi, H.J.; Siukstaite, L.; Harrison, O.J.; Brasch, J.; Goodman, K.M.; Hansen, L.; Shapiro, L.; Honig, B.; et al. Discovery of an O-mannosylation pathway selectively serving cadherins and protocadherins. Proc. Natl. Acad. Sci. USA 2017, 114, 11163–11168. [Google Scholar] [CrossRef] [PubMed]
- Neubert, P.; Strahl, S. Protein O-mannosylation in the early secretory pathway. Curr. Opin. Cell Biol. 2016, 41, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Loibl, M.; Strahl, S. Protein O-mannosylation: What we have learned from baker’s yeast. Biochim. Biophys. Acta 2013, 1833, 2438–2446. [Google Scholar] [CrossRef] [PubMed]
- Ecker, M.; Mrsa, V.; Hagen, I.; Deutzmann, R.; Strahl, S.; Tanner, W. O-mannosylation precedes and potentially controls the N-glycosylation of a yeast cell wall glycoprotein. EMBO Rep. 2003, 4, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Lussier, M.; Sdicu, A.M.; Bussey, H. The KTR and MNN1 mannosyltransferase families of Saccharomyces cerevisiae. Biochim. Biophys. Acta 1999, 1426, 323–334. [Google Scholar] [CrossRef]
- Neubert, P.; Halim, A.; Zauser, M.; Essig, A.; Joshi, H.J.; Zatorska, E.; Larsen, I.S.B.; Loibl, M.; Castells-Ballester, J.; Aebi, M.; et al. Mapping the O-mannose glycoproteome in Saccharomyces cerevisiae. Mol. Cell Prot. 2016. [Google Scholar] [CrossRef] [PubMed]
- Gentzsch, M.; Tanner, W. The PMT gene family: Protein O-glycosylation in Saccharomyces cerevisiae is vital. EMBO J. 1996, 15, 5752–5759. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.; Hutzler, J.; Bermejo, C.; Ragni, E.; García-Cantalejo, J.; Botías, P.; Piberger, P.; Schott, A.; Sanz, A.B.; Strahl, S. Functional and genomic analyses of blocked protein O-mannosylation in baker’s yeast. Mol. Microbiol. 2011, 79, 1529–1546. [Google Scholar] [CrossRef] [PubMed]
- Zatorska, E.; Gal, L.; Schmitt, J.; Bausewein, D.; Schuldiner, M.; Strahl, S. Cellular Consequences of Diminished Protein O-Mannosyltransferase Activity in Baker’s Yeast. Int. J. Mol. Sci. 2017, 18, 1226. [Google Scholar] [CrossRef] [PubMed]
- Lommel, M.; Bagnat, M.; Strahl, S. Aberrant processing of the WSC family and Mid2p cell surface sensors results in cell death of Saccharomyces cerevisiae O-mannosylation mutants. Mol. Cell Biol. 2004, 24, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Petkova, M.I.; Pujol-Carrion, N.; Torre-Ruiz, M.A. Mtl1 O-mannosylation mediated by both Pmt1 and Pmt2 is important for cell survival under oxidative conditions and TOR blockade. Fungal Genet. Biol. 2012, 49, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Weber, Y.; Prill, S.K.; Ernst, J.F. Pmt-mediated O-mannosylation stabilizes an essential component of the secretory apparatus, Sec20p, in Candida albicans. Eukaryot. Cell 2004, 3, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Ng, D.T.W. O-mannosylation: The other glycan player of ER quality control. Semin. Cell Dev. Biol. 2015, 41, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Vashist, S.; Kim, W.; Belden, W.J.; Spear, E.D.; Barlowe, C.; Ng, D.T.W. Distinct retrieval and retention mechanisms are required for the quality control of endoplasmic reticulum protein folding. J. Cell Biol. 2001, 155, 355–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatsukasa, K.; Okada, S.; Umebayashi, K.; Fukuda, R.; Nishikawa, S.I.; Endo, T. Roles of O-mannosylation of aberrant proteins in reduction of the load for endoplasmic reticulum chaperones in yeast. J. Biol. Chem. 2004, 279, 49762–49772. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Spooner, R.A.; Hampton, R.A.; Lord, J.M.; Roberts, L.M. Cytosolic entry of Shiga-like toxin a chain from the yeast endoplasmic reticulum requires catalytically active Hrd1p. PLoS ONE 2012, 7, e41119. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, H.; Fujita, M.; Yoko-o, T.; Jigami, Y. O-mannosylation is required for degradation of the endoplasmic reticulum-associated degradation substrate Gas1*p via the ubiquitin/proteasome pathway in Saccharomyces cerevisiae. J. Biochem. 2008, 143, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Harty, C.; Strahl, S.; Romisch, K. O-mannosylation protects mutant alpha-factor precursor from endoplasmic reticulum-associated degradation. Mol. Biol. Cell 2001, 12, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, W.; Thibault, G.; Ng, D.T.W. Futile protein folding cycles in the ER are terminated by the unfolded protein O-mannosylation pathway. Science 2013, 340, 978–981. [Google Scholar] [CrossRef] [PubMed]
- Khmelinskii, A.; Keller, P.J.; Bartosik, A.; Meurer, M.; Barry, J.D.; Mardin, B.R.; Kaufmann, A.; Trautmann, S.; Wachsmuth, M.; Pereira, G.; et al. Tandem fluorescent protein timers for in vivo analysis of protein dynamics. Nat. Biotechnol. 2012, 30, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Khmelinskii, A.; Blaszczak, E.; Pantazopoulou, M.; Fischer, B.; Omnus, D.J.; Dez, G.L.; Brossard, A.; Gunnarsson, A.; Barry, J.D.; Meurer, M.; et al. Protein quality control at the inner nuclear membrane. Nature 2014, 516, 410–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khmelinskii, A.; Meurer, M.; Ho, C.T.; Besenbeck, B.; Füller, J.; Lemberg, M.K.; Bukau, B.; Mogk, B.; Knop, M. Incomplete proteasomal degradation of green fluorescent proteins in the context of tandem fluorescent protein timers. Mol. Biol. Cell 2016, 27, 360–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baryshnikova, A.; Costanzo, M.; Dixon, S.; Vizeacoumar, F.J.; Myers, C.L.; Andrews, B.; Boone, C. Synthetic genetic array (SGA) analysis in Saccharomyces cerevisiae and Schizosaccharomyces pombe. Methods Enzymol. 2010, 470, 145–179. [Google Scholar] [PubMed]
- Tsirigos, K.D.; Peters, C.; Shu, N.; Käll, L.; Elofsson, A. The TOPCONS web server for consensus prediction of membrane protein topology and signal peptides. Nucleic Acids Res. 2015, 43, W401–W407. [Google Scholar] [CrossRef] [PubMed]
- Girrbach, V.; Strahl, S. Members of the evolutionarily conserved PMT family of protein O-mannosyltransferases form distinct protein complexes among themselves. J. Biol. Chem. 2003, 278, 12554–12562. [Google Scholar] [CrossRef] [PubMed]
- Verna, J.; Lodder, A.; Lee, K.; Vagts, A.; Ballester, R. A family of genes required for maintenance of cell wall integrity and for the stress response in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1997, 94, 13804–13809. [Google Scholar] [CrossRef] [PubMed]
- Lodder, A.L.; Lee, T.K.; Ballester, R. Characterization of the Wsc1 protein, a putative receptor in the stress response of Saccharomyces cerevisiae. Genetics 1999, 152, 1487–1499. [Google Scholar] [PubMed]
- Roemer, T.; Madden, K.; Chang, J.; Snyder, M. Selection of axial growth sites in yeast requires Axl2p, a novel plasma membrane glycoprotein. Genes Dev. 1996, 10, 777–793. [Google Scholar] [CrossRef] [PubMed]
- Sanders, S.L.; Gentzsch, M.; Tanner, W.; Herskowitz, I. O-Glycosylation of Axl2/Bud10p by Pmt4p is required for its stability, localization, and function in daughter cells. J. Cell Biol. 1999, 145, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Roemer, T.; Bussey, H. Yeast beta-glucan synthesis: KRE6 encodes a predicted type II membrane protein required for glucan synthesis in vivo and for glucan synthase activity in vitro. Proc. Natl. Acad. Sci. USA 1991, 88, 11295–11299. [Google Scholar] [CrossRef] [PubMed]
- Kurita, T.; Noda, Y.; Takagi, T.; Osumi, M.; Yoda, K. Kre6 Protein Essential for Yeast Cell Wall beta-1,6-Glucan Synthesis Accumulates at Sites of Polarized Growth. J. Biol. Chem. 2011, 286, 7429–7438. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, M.; Kimata, Y.; Kohno, K. Saccharomyces cerevisiae Rot1 is an essential molecular chaperone in the endoplasmic reticulum. Mol. Biol. Cell 2008, 19, 3514–3525. [Google Scholar] [CrossRef] [PubMed]
- Kurita, T.; Noda, Y.; Yoda, K. Action of Multiple Endoplasmic Reticulum Chaperon-like Proteins Is Required for Proper Folding and Polarized Localization of Kre6 Protein Essential in Yeast Cell Wall beta-1,6-Glucan Synthesis. J. Biol. Chem. 2012, 287, 17415–17424. [Google Scholar] [CrossRef] [PubMed]
- Ast, T.; Cohen, G.; Schuldiner, M. A network of cytosolic factors targets SRP-independent proteins to the endoplasmic reticulum. Cell 2013, 152, 1134–1145. [Google Scholar] [CrossRef] [PubMed]
- Kats, I.; Khmelinskii, A.; Kschonsak, M.; Huber, M.; Knieß, R.A.; Bartosik, A.; Knop, M. Mapping Degradation Signals and Pathways in a Eukaryotic N-terminome. Mol. Cell 2018, 70, 488–501. [Google Scholar] [CrossRef] [PubMed]
- Gable, K.; Slife, H.; Bacikova, D.; Monaghan, E.; Dunn, T.M. Tsc3p is an 80-amino acid protein associated with serine palmitoyltransferase and required for optimal enzyme activity. J. Biol. Chem. 2000, 275, 7597–7603. [Google Scholar] [CrossRef] [PubMed]
- Helms, J.B.; Zurzolo, C. Lipids as targeting signals: Lipid rafts and intracellular trafficking. Traffic 2004, 5, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Goder, V.; Melero, A. Protein O-mannosyltransferases participate in ER protein quality control. J. Cell Sci. 2011, 124, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Bausewein, D.; Engel, J.; Jank, T.; Schoedl, M.; Strahl, S. Functional similarities between the protein O-mannosyltransferases Pmt4 from baker’s yeast and human POMT1. J. Biol. Chem. 2016. [Google Scholar] [CrossRef] [PubMed]
- Gentzsch, M.; Tanner, W. Protein-O-glycosylation in yeast: Protein-specific mannosyltransferases. Glycobiology 1997, 7, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Willer, T.; Brandl, M.; Sipiczki, M.; Strahl, S. Protein O-mannosylation is crucial for cell wall integrity, septation and viability in fission yeast. Mol. Microbiol. 2005, 57, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Jungmann, J.; Munro, S. Multi-protein complexes in the cis Golgi of Saccharomyces cerevisiae with alpha-1,6-mannosyltransferase activity. EMBO J. 1998, 17, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Gueldener, U.; Heinisch, J.; Koehler, G.J.; Voss, D.; Hegemann, J.H. A second set of loxP marker cassettes for Cre-mediated multiple gene knockouts in budding yeast. Nucleic Acids Res. 2002, 30. [Google Scholar] [CrossRef]
- Hill, J.; Donald, K.A.; Griffiths, D.E. DMSO-enhanced whole cell yeast transformation. Nucleic Acids Res. 1991, 19. [Google Scholar] [CrossRef]
- Meurer, M.; Duan, Y.; Sass, E.; Kats, L.; Herbst, K.; Buchmuller, B.C.; Buchmuller, V.; Huber, F.; Kirrmaier, D.; Štefl, M. Genome-wide C-SWAT library for high-throughput yeast genome tagging. Nat. Methods 2018, 15, 598–600. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 12 September 2018).
- Wickham, H. Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Slowikowski, K. Repulsive Text and Label Geoms for ‘ggplot2’. Available online: https://CRAN.R-project.org/package=ggrepel (accessed on 12 September 2018).
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Huber, W.; Liaw, A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; et al. Various R Programming Tools for Plotting Data. Available online: https://CRAN.R-project.org/package=gplots (accessed on 12 September 2018).
- Treco, D.A.; Winston, F. Growth and manipulation of yeast. Curr. Protoc. Mol. Biol. 2008, 13. [Google Scholar] [CrossRef]
- Tran, J.R.; Brodsky, J.L. Assays to measure ER-associated degradation in yeast. Methods Mol. Biol. 2012, 832, 505–518. [Google Scholar] [PubMed]
Sample Availability: Yeast strains and oligonucleotides are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pmtΔ Mutant | tFT-Fusion Protein | WTmCherry/sfGFP/pmtΔmCherry/sfGFP ± SD | p-Value |
|---|---|---|---|
| pmt4Δ | Axl2 | 1.118 ± 0.033 | 0.088 |
| pmt1Δ | Kre6 | 0.842 ± 0.027 | 0.037 |
| pmt1Δ | Vrg4 | 0.873 ± 0.040 | 0.047 |
| pmt2Δ | YNL058C | 0.787± 0.042 | 0.010 |
| Strain | Genotype | Source |
|---|---|---|
| BY4741 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | Euroscarf |
| MLY201 | BY4741 except pmt1Δ::KANMX6 | This study |
| MLY202 | BY4741 except pmt2Δ::KANMX6 | This study |
| MLY204 | BY4741 except pmt4Δ::KANMX6 | This study |
| YMaM330 | MATα can1Δ::STE2pr-SpHIS5 lyp1Δ::STE3pr-LEU2 his3Δ1 leu2Δ0::GAL1pr-I-SCEI-natNT2 ura3Δ0 | [22] |
| WT Kre6-tFT | YMaM330 except KRE6::mCherry-sfGFP | [22] |
| EZY91 | WT Kre6-tFT except pmt1Δ::KANMX6 | This study |
| WT Vrg4-tFT | YMaM330 except VRG4::mCherry-sfGFP | [22] |
| EYZ96 | WT Vrg4-tFT except pmt1Δ::KANMX6 | This study |
| WT Axl2-tFT | YMaM330 except AXL2::mCherry-sfGFP | [22] |
| EZY107 | WT Axl2-tFT except pmt4Δ | This study |
| WT YNL058C-tFT | YMaM330 except YNL058C::mCherry-sfGFP | [22] |
| EZY106 | WT YNL058C-tFT except pmt2Δ | This study |
| WT Mnn11-tFT | YMaM330 except MNN11::mCherry-sfGFP | [22] |
| EZY109 | WT Mnn11-tFT except pmt4Δ | This study |
| MGY69 | AXL2::HA | [30] |
| MGY72 | AXL2::HA except pmt4Δ | [30] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castells-Ballester, J.; Zatorska, E.; Meurer, M.; Neubert, P.; Metschies, A.; Knop, M.; Strahl, S. Monitoring Protein Dynamics in Protein O-Mannosyltransferase Mutants In Vivo by Tandem Fluorescent Protein Timers. Molecules 2018, 23, 2622. https://doi.org/10.3390/molecules23102622
Castells-Ballester J, Zatorska E, Meurer M, Neubert P, Metschies A, Knop M, Strahl S. Monitoring Protein Dynamics in Protein O-Mannosyltransferase Mutants In Vivo by Tandem Fluorescent Protein Timers. Molecules. 2018; 23(10):2622. https://doi.org/10.3390/molecules23102622
Chicago/Turabian StyleCastells-Ballester, Joan, Ewa Zatorska, Matthias Meurer, Patrick Neubert, Anke Metschies, Michael Knop, and Sabine Strahl. 2018. "Monitoring Protein Dynamics in Protein O-Mannosyltransferase Mutants In Vivo by Tandem Fluorescent Protein Timers" Molecules 23, no. 10: 2622. https://doi.org/10.3390/molecules23102622
APA StyleCastells-Ballester, J., Zatorska, E., Meurer, M., Neubert, P., Metschies, A., Knop, M., & Strahl, S. (2018). Monitoring Protein Dynamics in Protein O-Mannosyltransferase Mutants In Vivo by Tandem Fluorescent Protein Timers. Molecules, 23(10), 2622. https://doi.org/10.3390/molecules23102622