Bioactive Constituents of Lamium album L. as Inhibitors of Cytokine Secretion in Human Neutrophils




Abstract
:1. Introduction
2. Results
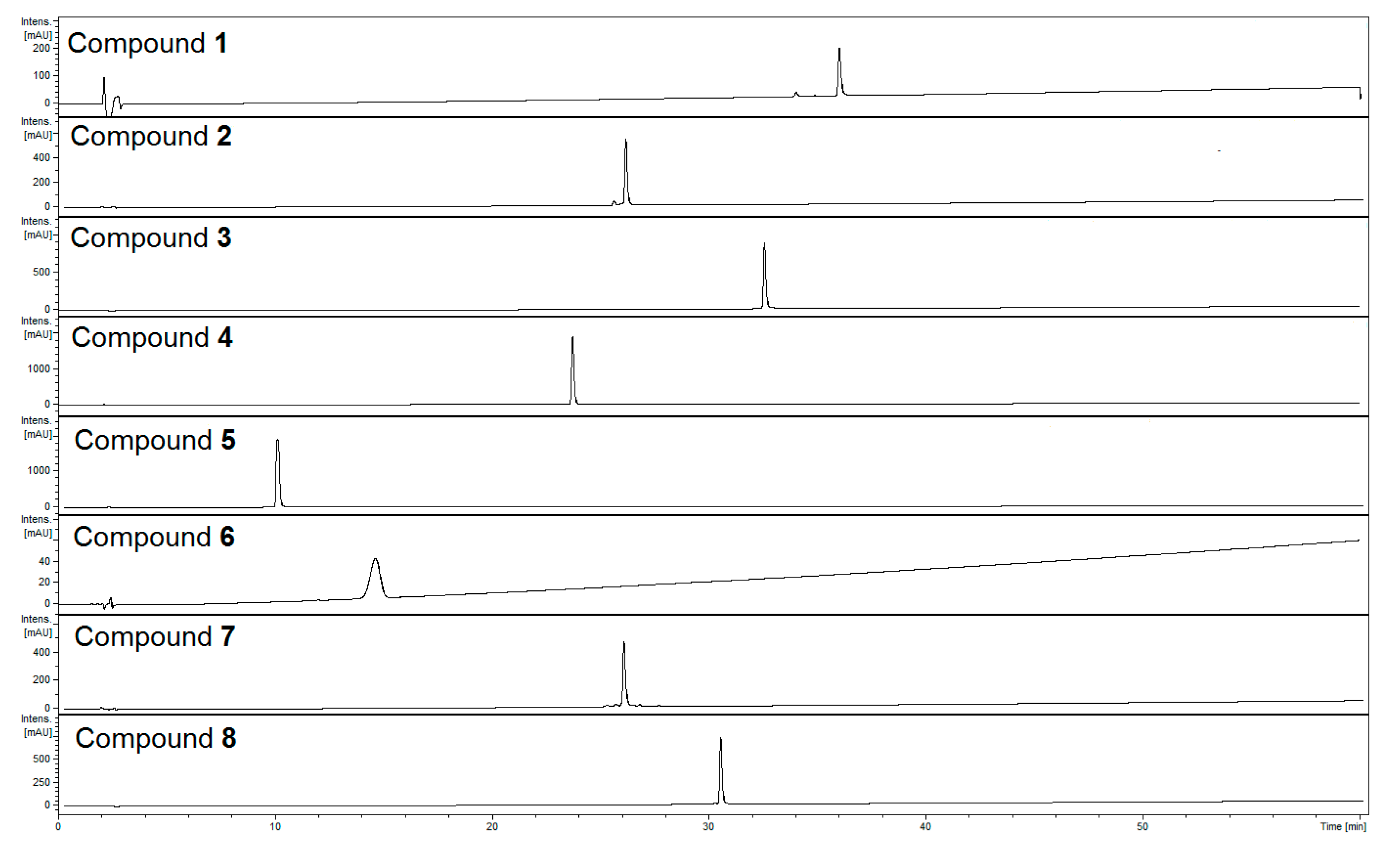
2.1. Identification of Compounds
2.2. Cytotoxicity of Compounds
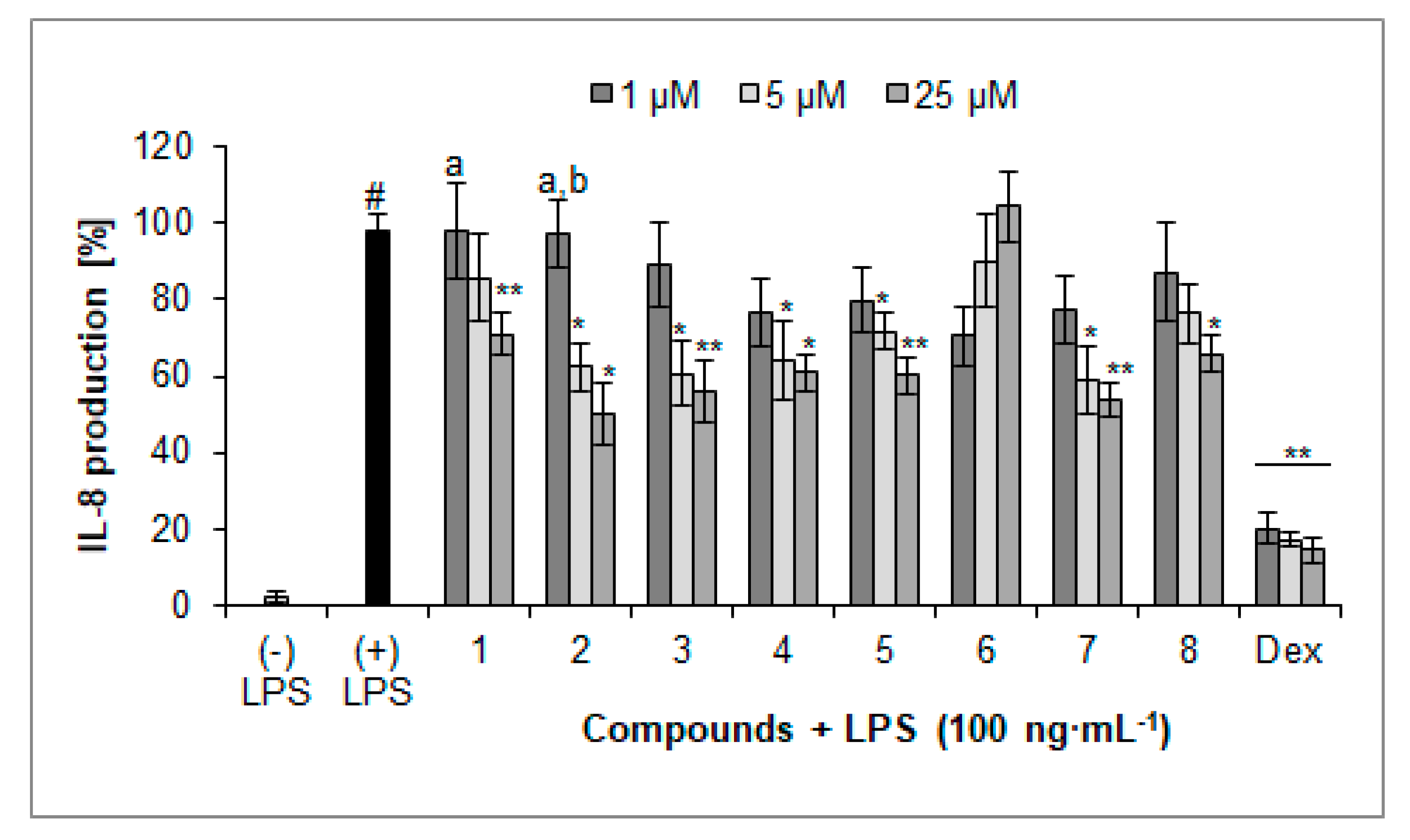
2.3. Inhibition of IL-8 Secretion
2.4. Inhibition of TNF-α Production
2.5. Inhibition of ROS Production
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material
4.3. Compounds Isolation
4.4. Phytochemical Analysis by HPLC-DAD-MSn Method
4.5. PMNs Isolation
4.6. Cytotoxicity
4.7. TNF-α and IL-8 Production by PMNs
4.8. Evaluation of ROS Production by Human Neutrophils
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yalçin, F.N.; Kaya, D. Ethnobotany, pharmacology and phytochemistry of the genus Lamium (Lamiaceae). Fabad J. Pharm. Sci. 2006, 31, 43–53. [Google Scholar]
- Turner, N.; Łuczaj, Ł.; Migliorini, P.; Pieroni, A.; Dreon, A.; Sacchetti, L. Edible and tended wild plants, traditional ecological knowledge and agroecology. Crit. Rev. Plant Sci. 2011, 30, 198–225. [Google Scholar] [CrossRef]
- Łuczaj, Ł. Archival data on wild food plants used in Poland in 1948. J. Ethnobiol. Ethnomed. 2008, 4, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinrich, M.; Müller, W.; Galli, C. Local Mediterranean Food Plants and Nutraceuticals; Forum Nutr. Karger: Basel, Switzerland, 2006; Volume 59, pp. 1–17. [Google Scholar]
- Pereira, O.R.; Domingues, M.R.M.; Silva, A.M.S.; Cardoso, S.M. Phenolic constituents of Lamium album: Focus on isoscutellarein derivatives. Food Res. Int. 2012, 48, 330–335. [Google Scholar] [CrossRef]
- Yordanova, Z.P.; Zhiponova, M.K.; Iakimova, E.T.; Dimitrova, M.A.; Kapchina-Toteva, V.M. Revealing the reviving secret of the white dead nettle (Lamium album L.). Phytochem. Rev. 2014, 13, 375–389. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Seigler, D.S. Phenylpropanoids. In Plant Secondary Metabolism; Springer: Boston, MA, USA, 1998; pp. 106–129. [Google Scholar]
- Kurkin, V. Phenylpropanoids from medicinal plants: Distribution, classification, structural analysis, and biological activity. Chem. Nat. Comp. 2003, 39, 123–153. [Google Scholar] [CrossRef]
- Bruneton, J. Pharmacognosy: Phytochemistry of Medicinal Plants; Lavoisier: Paris, France, 1999. [Google Scholar]
- Budzianowski, J.; Skrzypczak, L. Phenylpropanoid esters from Lamium album flowers. Phytochemistry 1995, 38, 997–1001. [Google Scholar] [CrossRef]
- Czerwińska, M.E.; Świerczewska, A.; Woźniak, M.; Kiss, A.K. Bioassay-guided isolation of iridoids and phenylpropanoids from aerial parts of Lamium album and their anti-inflammatory activity in human neutrophils. Planta Med. 2017, 83, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Damtoft, S.; Jensen, S.R.; Nielsen, B.J. Biosynthesis of iridoid glucosides in Lamium album. Phytochemistry 1991, 31, 135–137. [Google Scholar] [CrossRef]
- Alipieva, K.I.; Taskova, R.M.; Jensen, S.R.; Handjieva, N.V. Iridoid glucosides from Lamium album and Lamium maculatum (Lamiaceae). Biochem. Syst. Ecol. 2006, 34, 88–91. [Google Scholar] [CrossRef]
- Mitreski, I.; Stanoeva, J.P.; Stefova, M.; Stefkov, G.; Kulevanova, S. Polyphenols in representative Teucrium species in the flora of R. Macedonia: LC/DAD/ESI-MS(n) profile and content. Nat. Prod. Commun. 2014, 9, 175–180. [Google Scholar] [PubMed]
- Calis, I.; Basaran, A.A.; Saracoglu, I.; Sticher, O.; Ruedi, P. Phlinosides A, B and C, three phenylpropanoid glycosides from Phlomis linearis. Phytochemistry 1990, 29, 1253–1257. [Google Scholar] [CrossRef]
- Calis, I.; Basaran, A.A.; Saracoglu, I.; Sticher, O.; Ruedi, P. Phlinosides D and E, phenylpropanoid glycosides, and iridoids from Phlomis linearis. Phytochemistry 1991, 30, 3073–3075. [Google Scholar] [CrossRef]
- Yalçin, N.F.; Ersöz, T.; Bedir, E.; Şahpaz, S.; Bailleul, F.; Khan, I.A.; Dönmez, A.A.; Çalis, I. Phlinoside F, a new phenylethanoid glycoside from Phlomis angustissima. Turk. J. Chem. 2005, 29, 417–423. [Google Scholar]
- Veitch, N.C.; Regos, I.; Kite, G.C.; Treutter, D. Acylated flavonol glycosides from the forage legume, Onobrychis viciifolia (sainfoin). Phytochemistry 2011, 72, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yuan, Q.; Liu, E.; Qi, L.; Bi, Z.; Li, P. Fragmentation study of iridoid glycosides and phenylpropanoid glycosides in Radix Scrophulariae by rapid resolution liquid chromatography with diode-array detection and electrospray ionization time-of-flight mass spectrometry. Biomed. Chromatogr. 2010, 24, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Alipieva, K.; Kokubun, T.; Taskova, R.; Evstatieva, L.; Handjieva, N. LC-ESI-MS analysis of iridoid glucosides in Lamium species. Biochem. Syst. Ecol. 2007, 35, 17–22. [Google Scholar] [CrossRef]
- Quade, M.J.; Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopathol. 1997, 58, 239–248. [Google Scholar] [CrossRef]
- Baetta, R.; Corsini, A. Role of polymorphonuclear neutrophils in atherosclerosis: Current state and future perspectives. Atherosclerosis 2010, 210, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Palsson-McDermott, E.M.; O’Neill, L.A. Signal transduction by the lipopolysaccharide receptor, Toll-like receptor-4. Immunology 2004, 113, 153–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulua, A.C.; Simon, A.; Maddipati, R.; Pelletier, M.; Park, H.; Kim, K.Y.; Sack, M.N.; Kastner, D.L.; Siegel, R.M. Mitochondrial reactive oxygen species promote production of proinflammatory cytokines and are elevated in TNFR1-associated periodic syndrome (TRAPS). J. Exp. Med. 2011, 208, 519–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, K.A.; Smith, M.F.; Sanders, M.K.; Ernst, P.B. Reactive oxygen and nitrogen species differentially regulate Toll-like receptor 4-mediated activation of NF-kappa B and interleukin-8 expression. Infect. Immun. 2004, 72, 2123–2130. [Google Scholar] [CrossRef] [PubMed]
- Bickel, M. The role of interleukin-8 in inflammation and mechanisms of regulation. J. Periodontol. 1993, 64, 456–460. [Google Scholar] [PubMed]
- Harada, A.; Sekido, N.; Akahoshi, T.; Wada, T.; Mukaida, N.; Matsushima, K. Essential involvement of interleukin-8 (IL-8) in acute inflammation. J. Leukoc. Biol. 1994, 56, 559–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Yang, B. Phenylethanoid glycosides: Research advances in their phytochemistry, pharmacological activity and pharmacokinetics. Molecules 2016, 21, 991. [Google Scholar] [CrossRef] [PubMed]
- Tundis, R.; Loizzo, M.R.; Menichini, F.; Statti, G.A.; Menichini, F. Biological and pharmacological activities of iridoids: Recent developments. Mini Rev. Med. Chem. 2008, 8, 399–420. [Google Scholar] [CrossRef] [PubMed]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Reale, M.; Menghini, L.; Vinciguerra, I.; De Lutiis, M.A.; Felaco, M.; Grilli, A. Antiinflammatory effects in THP-1 cells treated with verbascoside. Phytother. Res. 2010, 24, 1398–1404. [Google Scholar] [CrossRef] [PubMed]
- Hausmann, M.; Obermeier, F.; Paper, D.H.; Balan, K.; Dunger, N.; Menzel, K.; Falk, W.; Schoelmerich, J.; Herfarth, H.; Rogler, G. In vivo treatment with the herbal phenylethanoid acteoside ameliorates intestinal inflammation in dextran sulphate sodium-induced colitis. Clin. Exp. Immunol. 2007, 148, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.L.; Chang, H.S.; Tseng, Y.F.; Shi, L.S. Suppression of IL-8 release by sweet olive ethanolic extract and compounds in WiDr colon adenocarcinoma cells. J. Food Sci. 2017, 82, 1792–1798. [Google Scholar] [CrossRef] [PubMed]
- Serreli, G.; Incani, A.; Atzeri, A.; Angioni, A.; Campus, M.; Cauli, E.; Zurru, R.; Deiana, M. Antioxidant effect of natural table olives phenolic extract against oxidative stress and membrane damage in enterocyte-like cells. J. Food Sci. 2017, 82, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Kupeli, E.; Harput, U.S.; Varel, M.; Yesilada, E.; Saracoglu, I. Bioassay-guided isolation of iridoid glucosides with antinociceptive and anti-inflammatory activities from Veronica anagallis-aquatica L. J. Ethnopharmacol. 2005, 102, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.J.; Lim, K.H.; Jung, H.J.; Park, E.H. Anti-inflammatory evaluation of gardenia extract, geniposide and genipin. J. Ethnopharmacol. 2006, 103, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Gyurkovska, V.; Alipieva, K.; Maciuk, A.; Dimitrova, P.; Ivanovska, N.; Haas, C.; Bley, T.; Georgiev, M. Anti-inflammatory activity of Devil’s claw in vitro systems and their active constituents. Food Chem. 2011, 125, 171–178. [Google Scholar] [CrossRef]
- Mohamed, N.M.; Makboul, M.A.; Farag, S.F.; Tarawenh, A.H.; Khan, S.I.; Brooks, T.A.; Wang, Y.-H.; Roos, S.A. Iridoid and phenylpropanoid glycosides from the roots of Lantana montevidensis. Med. Chem. Res. 2017, 26, 1117–1126. [Google Scholar] [CrossRef]
- Tutin, T.; Heywood, V.; Burges, N.; Moore, D.; Valentine, D.; Walters, S.; Webb, D. Flora Europaea; Cambridge at the University Press: Cambridge, UK, 1972; Volume 3. [Google Scholar]
- Rutkowski, L. Klucz Do Oznaczania Roślin Naczyniowych Polski Niżowej, 2nd ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2011. [Google Scholar]
- Böyum, A. A one-stage procedure for isolation of granulocytes and lymphocytes from human blood. General sedimentation properties of white blood cells in a 1g gravity field. Scand. J. Clin. Lab. Invest. 1968, 97, 51–76. [Google Scholar]
Sample Availability: Samples of the compounds 2–7 are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte No. | Compound | UV λmax [nm] | [M − H]− m/z | Fragmentary Ions |
|---|---|---|---|---|
| 1 | Apigenin 7-O-β-d-(6′′-trans-p-coumaroyl)-glucoside | 269, 317 | 577 | 307, 269 |
| 2 | Kaempferol 3-O-glucoside (astragalin) | 235sh, 265, 342 | 447 | 327, 285, 255 |
| 3 | Quercetin 3-O-[(4′′′-O-E-feruloyl)-α-rhamnopyranosyl-(1→6)]-β-glucopyranoside | 251, 333 | 785 | 623, 609, 591, 477, 301, 271 |
| 4 | Verbascoside | 245, 329 | 623 | 461, 315 |
| 5 | Lamalbid (lamiridoside) | 241 | 467 * | 421, 259 |
| 6 | Shanzhiside methyl ester | 221 | 451 * | 405, 283, 225, 179, 143 |
| 7 | Phlinoside D | 216, 328 | 769 | 637, 607, 593, 461 |
| 8 | Quercetin 3-O-α-rhamnopyranosyl-(1→2)[(4′′′′-O-E-feruloyl)-α-rhamnopyranosyl-(1→6)]-β-glucopyranoside | 251, 332 | 931 | 887, 785, 769, 755, 738, 702, 609, 562, 490, 301 |
| Quercetin | Compound 3 | Compound 8 | ||
|---|---|---|---|---|
| 1H | 13C | 1H | 13C | |
| 2 | - | 159.1 | - | 158.8 |
| 3 | - | 135.3 | - | 134.2 |
| 4 | - | 179.4 | - | 179.4 |
| 4a | - | 105.6 | - | 105.8 |
| 5 | - | 163.1 | - | 163.1 |
| 6 | 6.19 (d, J = 2.0 Hz) | 99.9 | 6.17 (d, J = 2.1 Hz) | 99.7 |
| 7 | - | 166.0 | - | 165.6 |
| 8 | 6.33 (d, J = 2.0 Hz) | 94.8 | 6.30 (d, J = 2.1 Hz) | 94.6 |
| 8a | - | 158.5 | - | 158.5 |
| 1′ | - | 123.2 | - | 123.3 |
| 2′ | 7.65 (d, J = 2.1 Hz) | 117.5 | 7.58 (d, J = 2.1 Hz) | 117.5 |
| 3′ | - | 146.0 | - | 146.1 |
| 4′ | - | 149.7 | - | 149.4 |
| 5′ | 6.92 (d, J = 8.4 Hz) | 116.1 | 6.94 (d, J = 8.2 Hz) | 116.1 |
| 6′ | 7.64 (dd, J = 8.4, 2.1 Hz) | 123.4 | 7.56 (m) | 123.5 |
| d-Glucose | ||||
| 1′′ | 5.31 (d, J = 7.7 Hz) | 103.8 | 5.78 (d, J = 7.7 Hz) | 100.1 |
| 2′′ | 3.50 (m) | 75.7 | 3.70 (m) | 79.6 |
| 3′′ | 3.45 (m) | 78.1 | 3.58 (m) | 79.1 |
| 4′′ | 3.39 (m) | 71.1 | 3.49 (m) | 71.1 |
| 5′′ | 3.40 (m) | 76.9 | 3.37 (m) | 76.6 |
| 6′′ | 3.85, 3.53 (m) | 68.0 | 3.83, 3.55 (m) | 67.6 |
| l-Rhamnose I | ||||
| 1′′′ | 4.61 (d, J = 1.6 Hz) | 102.1 | 4.61 (d, J = 1.6 Hz) | 101.7 |
| 2′′′ | 3.75 (dd, J = 3.4, 1.6 Hz) | 72.2 | 3.75 (m) | 72.2 |
| 3′′′ | 3.80 (dd, J = 9.8, 3.4 Hz) | 70.3 | 3.73 (m) | 72.2 |
| 4′′′ | 4.92 (t, J = 9.8 Hz) | 75.2 | 4.88 (t, J = 9.8 Hz) | 75.1 |
| 5′′′ | 3.66 (m) | 67.7 | 3.61 (m) | 67.6 |
| 6′′′ | 0.90 (d, J = 6.3 Hz) | 17.6 | 0.81 (d, J = 6.3 Hz) | 17.4 |
| l-Rhamnose II | ||||
| 1′′′′ | 5.24 (d, J = 1.6 Hz) | 102.3 | ||
| 2′′′′ | 3.98 (dd, J = 3.4, 1.7 Hz) | 72.3 | ||
| 3′′′′ | 3.80 (m) | 70.3 | ||
| 4′′′′ | 3.29 (m) | 74.0 | ||
| 5′′′′ | 3.95 (m) | 69.8 | ||
| 6′′′′ | 0.86 (d, J = 6.3 Hz) | 17.3 | ||
| Feruloyl (*/#) | ||||
| α | 6.26 (d, J = 15.9 Hz) | 116.3 | 6.16 (d, J = 15.9 Hz) | 116.3 |
| β | 7.56 (d, J = 15.9 Hz) | 146.8 | 7.53 (d, J = 15.9 Hz) | 146.8 |
| γ | - | 169.0 | - | 169.0 |
| 1′′′′/1′′′′′ | - | 129.9 | - | 129.0 |
| 2′′′′/2′′′′′ | 7.13 (d, J = 2.1 Hz) | 115.0 | 7.14 (d, J = 2.1 Hz) | 115.2 |
| 3′′′′/3′′′′′ | - | 147.8 | - | 147.9 |
| 4′′′′/4′′′′′ | - | 151.4 | - | 151.4 |
| 5′′′′/5′′′′′ | 6.98 (d, J = 8.4 Hz) | 112.5 | 7.00 (d, J = 8.4 Hz) | 112.6 |
| 6′′′′/6′′′′′ | 7.10 (dd, J = 8.4, 2.1 Hz) | 122.9 | 7.12 (dd, J = 8.3, 2.1 Hz) | 122.8 |
| OCH3 | 3.90 (s) | 56.3 | 3.92 (s) | 56.4 |
| Analyte | ROS Production [%] † |
|---|---|
| (−) f-MLP | 31.3 ± 2.6 |
| (+) f-MLP | 104.5 ± 8.3 # |
| 1 | 42.6 ± 5.1 * |
| 2 | 92.7 ± 6.2 |
| 3 | 16.9 ± 1.2 ** |
| 4 | 42.6 ± 4.2 * |
| 5 | 68.6 ± 5.2 * |
| 6 | 45.7 ± 1.4 * |
| 7 | 21.0 ± 3.4 ** |
| 8 | 10.0 ± 2.6 ** |
| Quercetin | 8.8 ± 1.8 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Czerwińska, M.E.; Świerczewska, A.; Granica, S. Bioactive Constituents of Lamium album L. as Inhibitors of Cytokine Secretion in Human Neutrophils. Molecules 2018, 23, 2770. https://doi.org/10.3390/molecules23112770
Czerwińska ME, Świerczewska A, Granica S. Bioactive Constituents of Lamium album L. as Inhibitors of Cytokine Secretion in Human Neutrophils. Molecules. 2018; 23(11):2770. https://doi.org/10.3390/molecules23112770
Chicago/Turabian StyleCzerwińska, Monika E., Anita Świerczewska, and Sebastian Granica. 2018. "Bioactive Constituents of Lamium album L. as Inhibitors of Cytokine Secretion in Human Neutrophils" Molecules 23, no. 11: 2770. https://doi.org/10.3390/molecules23112770
APA StyleCzerwińska, M. E., Świerczewska, A., & Granica, S. (2018). Bioactive Constituents of Lamium album L. as Inhibitors of Cytokine Secretion in Human Neutrophils. Molecules, 23(11), 2770. https://doi.org/10.3390/molecules23112770