
Phenolic Composition, Antioxidant Properties, and Inhibition toward Digestive Enzymes with Molecular Docking Analysis of Different Fractions from Prinsepia utilis Royle Fruits


Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterization and Quantification of Phenolic Compounds
2.2. Antioxidant Capacity
2.2.1. DPPH Radical Scavenging Activity
2.2.2. ABTS Radical Scavenging Activity
2.2.3. Inhibition of Intracellular ROS in H2O2-Induced HepG2 Cells
2.3. Pancreatic Lipase Inhibitory Activities of Phenolic Fractions from P. utilis Fruits and Authentic Standards
2.4. α-Glucosidase Inhibitory Activities of Phenolic Fractions from P. utilis Fruits and Authentic Standards
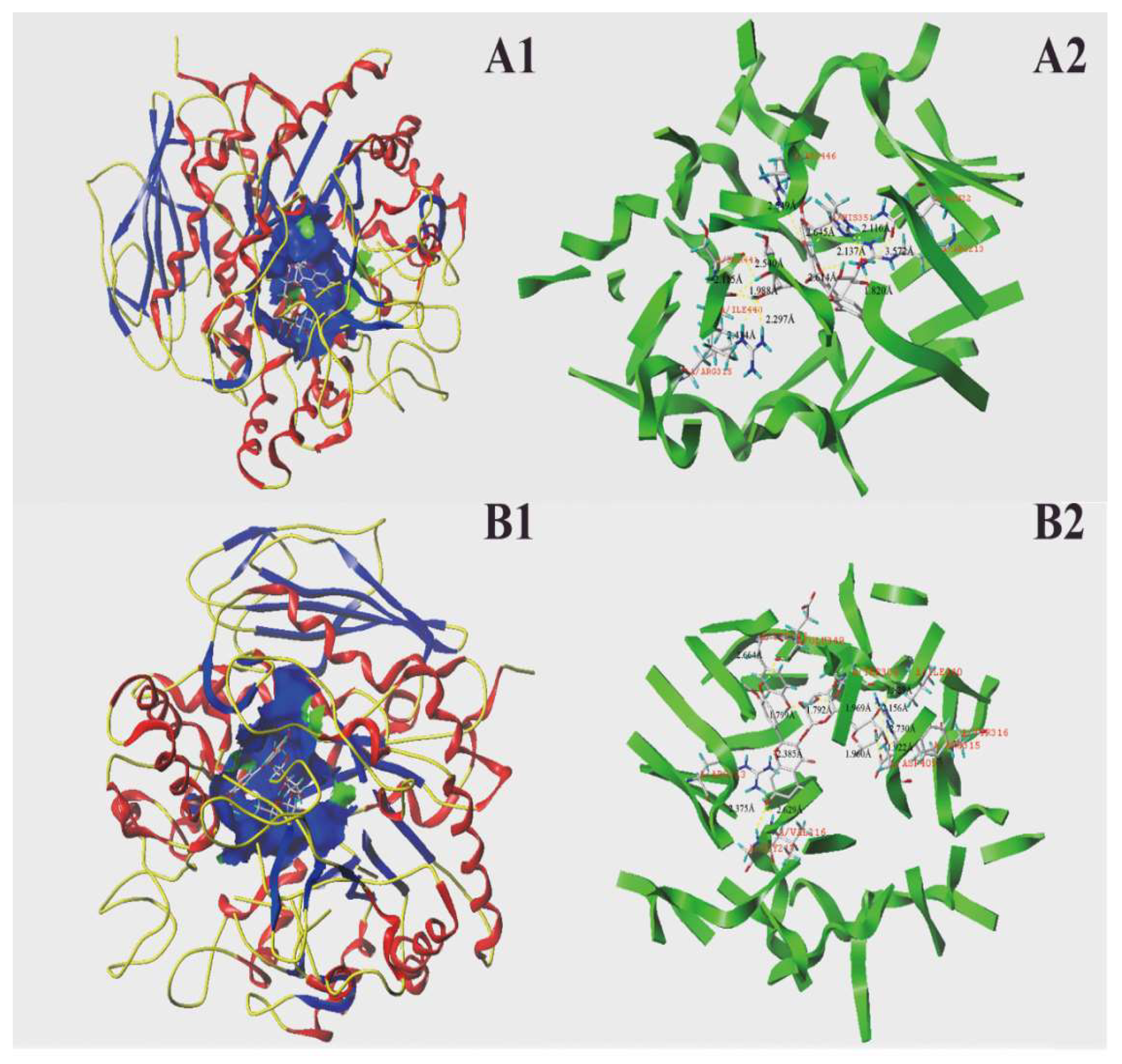
2.5. Molecular Docking Results
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Extraction and Fractionation
3.3. Identification and Quantification of Phenolics by UHPLC-ESI-HRMS/MS
3.4. DPPH Radical Scavenging Assay
3.5. ABTS Radical Scavenging Assay
3.6. Determination of Intracellular ROS
3.7. Determination of Pancreatic Lipase Inhibition
3.8. Determination of α-Glucosidase Inhibition
3.9. Molecular Docking
3.10. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Monforte, M.; Sánchez, E.; Barrio, F.; Costa, B.; Flores-Mateo, G. Metabolic syndrome and dietary patterns: A systematic review and meta-analysis of observational studies. Eur. J. Nutr. 2017, 56, 925–947. [Google Scholar] [CrossRef] [PubMed]
- Godos, J.; Zappalà, G.; Bernardini, S.; Giambini, I.; Bes-Rastrollo, M.; Martinez-Gonzalez, M. Adherence to the Mediterranean diet is inversely associated with metabolic syndrome occurrence: A meta-analysis of observational studies. Int. J. Jood Sci. Nutr. 2017, 68, 138–148. [Google Scholar] [CrossRef]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhang, D.; Jiang, X.; Jiang, W. Fruit and vegetable consumption and risk of type 2 diabetes mellitus: A dose-response meta-analysis of prospective cohort studies. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Tong, X.; Li, L.; Cao, S.; Yin, X.; Gao, C.; Herath, C.; Li, W.; Jin, Z.; Chen, Y. Consumption of fruit and vegetable and risk of coronary heart disease: A meta-analysis of prospective cohort studies. Int. J. Cardiol. 2015, 183, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.; Liu, Y.-J.; Cai, L.-B.; Xu, F.R.; Xie, T.; He, Q.Q. Fruit and vegetable consumption and risk of cardiovascular disease: A meta-analysis of prospective cohort studies. CRC Crit. Rev. Food Sci. Nutr. 2017, 57, 1650–1663. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K.; Tagami, M.; Yamori, Y. Dietary polyphenols regulate endothelial function and prevent cardiovascular disease. Nutrition 2015, 31, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Amiot, M.J.; Riva, C.; Vinet, A. Effects of dietary polyphenols on metabolic syndrome features in humans: A systematic review. Obes. Rev. 2016, 17, 573–586. [Google Scholar] [CrossRef]
- Chandrasekara, A.; Rasek, O.A.; John, J.A.; Chandrasekara, N.; Shahidi, F. Solvent and extraction conditions control the assayable phenolic content and antioxidant activities of seeds of black beans, canola and millet. J. Am. Oil Chem. Soc. 2016, 93, 275–283. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on α-amylase, α-glucosidase, pancreatic lipase and tyrosinas. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ma, Y.; Zhao, Y.; Cai, S.; Pang, M. Phenolic composition, antioxidant and pancreatic lipase inhibitory activities of Chinese sumac (Rhus chinensis Mill.) fruits extracted by different solvents and interaction between myricetin-3-O-rhamnoside and quercetin-3-O-rhamnoside. Int. J. Food Sci. Technol. 2018, 53, 1045–1053. [Google Scholar] [CrossRef]
- Zhang, C.; Ma, Y.; Gao, F.; Zhao, Y.; Cai, S.; Pang, M. The free, esterified, and insoluble-bound phenolic profiles of Rhus chinensis Mill. fruits and their pancreatic lipase inhibitory activities with molecular docking analysis. J. Funct. Foods 2018, 40, 729–735. [Google Scholar] [CrossRef]
- De Camargo, A.C.; Regitano-D’arce, M.A.B.; Biasoto, A.C.T.; Shahidi, F. Enzyme-assisted extraction of phenolics from winemaking by-products: Antioxidant potential and inhibition of alpha-glucosidase and lipase activities. Food Chem. 2016, 212, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Ma, Y.; Zhang, C.; Cai, S.; Pang, M. Bioaccessibility and antioxidant activity of phenolics in native and fermented Prinsepia utilis Royle seed during a simulated gastrointestinal digestion in vitro. J. Funct. Foods 2017, 37, 354–362. [Google Scholar] [CrossRef]
- Guan, B.; Li, T.; Xu, X.-K.; Wei, P.-L.; Peng, C.-C.; Fu, J.-J.; Zeng, Q.; Cheng, X.-R.; Zhang, S.-D. γ-Hydroxynitrile glucosides from the seeds of Prinsepia utilis. Phytochemistry 2014, 105, 135–140. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, H.-X.; Tan, H.-B.; Qiu, S.-X. Novel highly oxygenated and B-ring-seco-ent-diterpene glucosides from the seeds of Prinsepia utilis. Tetrahedron 2015, 71, 9415–9419. [Google Scholar] [CrossRef]
- Guan, B.; Peng, C.-C.; Zeng, Q.; Cheng, X.-R.; Yan, S.-K.; Jin, H.-Z.; Zhang, W.-D. Cytotoxic pentacyclic triterpenoids from Prinsepia utilis. Planta Med. 2013, 79, 365–368. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Oliveira, B.D.; Rodrigues, A.C.; Cardoso, B.M.I.; Ramos, A.L.C.C.; Bertoldi, M.C.; Taylor, J.G.; da Cunha, L.R.; Pinto, U.M. Antioxidant, antimicrobial and anti-quorum sensing activities of Rubus rosaefolius phenolic extract. Ind. Crops Prod. 2016, 84, 59–66. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Aguilera, Y.; Martin-Cabrejas, M.A.; De Mejia, E.G. Phenolic compounds in fruits and beverages consumed as part of the mediterranean diet: Their role in prevention of chronic diseases. Phytochem. Rev. 2016, 15, 405–423. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Arráez-Román, D.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Profiling of phenolic and other polar constituents from hydro-methanolic extract of watermelon (Citrullus lanatus) by means of accurate-mass spectrometry (HPLC–ESI–QTOF–MS). Food Res. Int. 2013, 51, 354–362. [Google Scholar] [CrossRef]
- Zhang, L.Z.; Zong, C.T.; Wang, H.; Fu, Z.F.; Wen, Q.H.; Dan, F. Metabolic profiling of antioxidants constituents in Artemisia selengensis leaves. Food Chem. 2015, 186, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Pantelić, M.; Dabić, D.; Matijašević, S.; Davidović, S.; DojIinović, B.; Milojković-Opsenica, D.; Tešić, Ž.; Natić, M. Chemical Characterization of Fruit Wine Made from Oblačinska Sour Cherry. Sci. World J. 2014, 2014, 454797. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Hu, Y.; Wang, H.; Huang, Y.; Zhang, P.; Liao, Z.; Chen, M. Profiling of Anthocyanins in transgenic purple-fleshed sweet potatoes by HPLC-MS/MS. J. Sci. Food Agric. 2017, 97, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.-Q.; Xiao, J.; Fan, H.-X.; Yu, Y.; He, R.-R.; Feng, X.-L.; Kurihara, H.; So, K.-F.; Yao, X.-S.; Gao, H. Polyphenols from wolfberry and their bioactivities. Food Chem. 2017, 214, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yano, S.; Chen, J.; Hisanaga, A.; Sakao, K.; He, X.; He, J.; Hou, D.-X. Polyphenols from Lonicera caerulea L. Berry Inhibit LPS-Induced Inflammation through Dual Modulation of Inflammatory and Antioxidant Mediators. J. Agric. Food Chem. 2017, 65, 5133–5141. [Google Scholar] [CrossRef] [PubMed]
- Ambigaipalan, P.; Al-Khalifa, A.S.; Shahidi, F. Antioxidant and angiotensin I converting enzyme (ACE) inhibitory activities of date seed protein hydrolysates prepared using Alcalase, Flavourzyme and Thermolysin. J. Funct. Foods 2015, 18, 1125–1137. [Google Scholar] [CrossRef]
- Patil, S.L.; Mallaiah, S.H.; Patil, R.K. Antioxidative and radioprotective potential of rutin and quercetin in Swiss albino mice exposed to gamma radiation. J. Med. Phys. 2013, 38, 87–92. [Google Scholar] [CrossRef]
- Galati, E.M.; Mondello, M.R.; Giuffrida, D.; Dugo, G.; Miceli, N.; Pergolizzi, S.; Taviano, M.F. Chemical Characterization and Biological Effects of Sicilian Opuntia ficus indica (L.) Mill. Fruit Juice: Antioxidant and Antiulcerogenic Activity. J. Agric. Food Chem. 2003, 51, 4903–4908. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Huang, S.; Cai, S.; Cao, J.; Han, P. Digestion property and synergistic effect on biological activity of purple rice (Oryza sativa L.) anthocyanins subjected to a simulated gastrointestinal digestion in vitro. Food Res. Int. 2015, 78, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Wang, O.; Wu, W.; Zhu, S.; Zhou, F.; Ji, B.; Gao, F.; Zhang, D.; Liu, J.; Cheng, Q. Comparative study of the effects of solid-state fermentation with three filamentous fungi on the total phenolics content (TPC), flavonoids, and antioxidant activities of subfractions from oats (Avena sativa L.). J. Agric. Food Chem. 2011, 60, 507–513. [Google Scholar] [CrossRef]
- Umeno, A.; Biju, V.; Yoshida, Y. In vivo ROS production and use of oxidative stress-derived biomarkers to detect the onset of diseases such as Alzheimer’s disease, Parkinson’s disease, and diabetes. Free Radic. Res. 2017, 51, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Ma, Q.; Guo, Y.; Sun, L. Protective effects of rambutan (Nephelium lappaceum) peel phenolics on H2O2-induced oxidative damages in HepG2 cells and d-galactose-induced aging mice. Food Chem. Toxicol. 2017, 108, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Cilla, A.; Rodrigo, M.J.; Zacarías, L.; De Ancos, B.; Sánchez-Moreno, C.; Barberá, R.; Alegría, A. Protective effect of bioaccessible fractions of citrus fruit pulps against H2O2-induced oxidative stress in Caco-2 cells. Food Res. Int. 2018, 103, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Weng, S.; Mao, L.; Gong, Y.; Sun, T.; Gu, Q. Role of quercetin in protecting ARPE-19 cells against H2O2-induced injury via nuclear factor erythroid 2 like 2 pathway activation and endoplasmic reticulum stress inhibition. Mol. Med. Rep. 2017, 16, 3461–3468. [Google Scholar] [CrossRef]
- Li, Y.; Cao, Z.; Zhu, H. Upregulation of endogenous antioxidants and phase 2 enzymes by the red wine polyphenol, resveratrol in cultured aortic smooth muscle cells leads to cytoprotection against oxidative and electrophilic stress. Pharmacol. Res. 2006, 53, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.J.; Ambigaipalan, P.; Shahidi, F. Biological Activities of Camelina and Sophia Seeds Phenolics: Inhibition of LDL Oxidation, DNA Damage, and Pancreatic Lipase and α-Glucosidase Activities. J. Food Sci. 2018, 83, 237–245. [Google Scholar] [CrossRef] [PubMed]
- De Camargo, A.C.; Regitano-D’arce, M.A.B.; Shahidi, F. Phenolic Profile of Peanut By-products: Antioxidant Potential and Inhibition of Alpha-Glucosidase and Lipase Activities. J. Am. Oil Chem. Soc. 2017, 94, 959–971. [Google Scholar] [CrossRef]
- Cai, S.; Wang, O.; Wang, M.; He, J.; Wang, Y.; Zhang, D.; Zhou, F.; Ji, B. In vitro inhibitory effect on pancreatic lipase activity of subfractions from ethanol extracts of fermented oats (Avena sativa L.) and synergistic effect of three phenolic acids. J. Agric. Food Chem. 2012, 60, 7245–7251. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Cui, F.; Yin, F.; Zeng, X.; Sun, Y.; Li, Y. Caffeoylquinic acids competitively inhibit pancreatic lipase through binding to the catalytic triad. Int. J. Biol. Macromol. 2015, 80, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhou, Q.; Chen, X.-Y.; Li, X.; Wang, Y.; Zhang, J.-l. Comparison and screening of bioactive phenolic compounds in different blueberry cultivars: Evaluation of anti-oxidation and α-glucosidase inhibition effect. Food Res. Int. 2017, 100, 312–324. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Chang, S.K.C.; Zhang, Y. Comparison of α-amylase, α-glucosidase and lipase inhibitory activity of the phenolic substances in two black legumes of different genera. Food Chem. 2017, 214, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradeep, P.M.; Sreerama, Y.N. Phenolic antioxidants of foxtail and little millet cultivars and their inhibitory effects on α-amylase and α-glucosidase activities. Food Chem. 2018, 247, 46–55. [Google Scholar] [CrossRef]
- Han, L.; Fang, C.; Zhu, R.; Peng, Q.; Li, D.; Wang, M. Inhibitory effect of phloretin on α-glucosidase: Kinetics, interaction mechanism and molecular docking. Int. J. Biol. Macromol. 2017, 95, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ding, H.; Hu, X.; Pan, J.; Liao, Y.; Gong, D.; Zhang, G. Exploring inhibitory mechanism of gallocatechin gallate on a-amylase and a-glucosidase relevant to postprandial hyperglycemia. J. Funct. Foods 2018, 48, 200–209. [Google Scholar] [CrossRef]
- Park, H.; Hwang, K.Y.; Oh, K.H.; Kim, Y.H.; Lee, J.Y.; Kim, K. Discovery of novel α-glucosidase inhibitors based on the virtual screening with the homology-modeled protein structure. Bioorg. Mmed. Chem. 2008, 16, 284–292. [Google Scholar] [CrossRef]
- Bolam, D.N.; Roberts, S.; Proctor, M.R.; Turkenburg, J.P.; Dodson, E.J.; Martinez-Fleites, C.; Yang, M.; Davis, B.G.; Davies, G.J.; Gilbert, H.J. The crystal structure of two macrolide glycosyltransferases provides a blueprint for host cell antibiotic immunity. Proc. Natl. Acad. Sci. USA 2007, 104, 5336–5341. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, D.; Zhang, D.; Lv, Y.; Wei, Y.; Wu, W.; Zhou, F.; Tang, M.; Mao, T.; Li, M. Inhibitory effect of blueberry polyphenolic compounds on oleic acid-induced hepatic steatosis in vitro. J. Agric. Food Chem. 2011, 59, 12254–12263. [Google Scholar] [CrossRef]
- Justino, A.B.; Miranda, N.C.; Franco, R.R.; Martins, M.M.; Silva, N.M.D.; Espindola, F.S. Annona muricata Linn. leaf as a source of antioxidant compounds with in vitro antidiabetic and inhibitory potential against α-amylase, α-glucosidase, lipase, non-enzymatic glycation and lipid peroxidation. Biomed. Pharmacother. 2018, 100, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Barakat, A.; Ali, M.; Al-Majid, A.M.; Yousuf, S.; Choudhary, M.I.; Khalil, R.; Ul-Haq, Z. Synthesis of thiobarbituric acid derivatives: In vitro α-glucosidase inhibition and molecular docking studies. Bioorg. Chem. 2017, 75, 99–105. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Compounds | RT a (min) | [M − H]− (m/z) | [M]+ (m/z) | Error (ppm) | Molecular Formula | MS/MS Fragment Ions | Reference | TPF * | FF * | AF * |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Protocatechuic acid | 3.68 | 153.0187 | 2.907 | C7H6O4 | 109.0284(66) | Standard | √ | √ | ||
| 2 | Penstemide | 4.77 | 443.1935 | 5.272 | C21H32O10 | 101.0233(90) | [23] | √ | |||
| 3 | p-Coumaric acid | 6.43 | 163.0394 | 0.495 | C9H8O3 | 119.0492(100) | Standard | √ | √ | ||
| 4 | Kaempferol-3-O-glucoside | 8.96 | 447.0946 | 5.328 | C21H20O11 | 284.0333(100), 285.0408(60) | Standard | √ | √ | ||
| 5 | Kaempferol-3-O-rhamnosylhexose | 10.32 | 593.1531 | 5.131 | C27H30O15 | 284.0328(10), 593.1530(100) | [24] | √ | √ | ||
| 6 | Dihydroquercetin rhamnoside | 10.88 | 449.1102 | 5.326 | C21H22O11 | 125.0235(61), 151.0030(20) | [24] | √ | √ | ||
| 7 | Catechin | 10.92 | 289.0726 | 6.695 | C15H14O6 | 123.0442(80), 109.0284(100) | Standard | √ | √ | ||
| 8 | Isoschaftoside | 13.56 | 563.1422 | 4.649 | C26H28O14 | 563.1422(100), 353.0677(61) | Standard | √ | √ | ||
| 9 | Rutin | 15.42 | 609.1476 | 4.217 | C27H30O16 | 300.0282(77), 609.1479(100) | Standard | √ | √ | √ | |
| 10 | Quercetin-3-O-glucoside | 16.01 | 463.0896 | 5.329 | C21H20O12 | 271.0254(30), 300.0282(100) | Standard | √ | √ | √ | |
| 11 | Quercetin 3-(6-O-acetyl-beta-glucoside) | 16.92 | 505.1003 | 5.173 | C23H22O13 | 300.0283(100), 301.0351(45) | Mass bank | √ | √ | ||
| 12 | Isorhamnetin-3-O-rutinoside | 17.39 | 623.1636 | 4.668 | C28H32O16 | 315.0519(100), 623.1638(38) | Standard | √ | √ | √ | |
| 13 | Kaempferol-3-O-hexoside | 17.63 | 447.0946 | 5.328 | C21H20O11 | 285.0395(30), 284.0334(100) | [24] | √ | √ | ||
| 14 | Isorhamnetin-3-O-glucoside | 18 | 477.1051 | 4.858 | C22H22O12 | 314.0441(100), 477.1050(20) | Standard | √ | √ | √ | |
| 15 | Cyanidin-3-O-glucoside | 9.98 | 449.1074 | −1.398 | C21H21O11 | 287.0548(100), 288.0580(17) | Standard | √ | √ | √ | |
| 16 | Cyanidin-3-O-rutinoside | 10.91 | 595.1655 | −1.086 | C27H31O15 | 287.0549(100), 449.1072(10) | [25] | √ | √ | √ | |
| 17 | Peonidin-3-O-rutinoside | 12.87 | 609.1810 | −0.700 | C28H33O15 | 301.0705(100), 463.1230(10) | [25] | √ | √ | ||
| 18 | Peonidin-3-O-sophoroside-5-O-glucoside | 13.38 | 787.2283 | −1.251 | C34H43O21 | 463.1229(10) | [26] | √ | √ | ||
| 19 | Delphinidin-3-O-rutinoside | 14.71 | 611.1603 | −0.722 | C27H31O16 | 303.049(100), 304.0528(20) | Mass bank | √ | √ | ||
| 20 | Petunidin-3-O-glucoside | 16.83 | 479.1178 | −1.070 | C22H23O12 | 317.0653(100) | [25] | √ | √ |
| Peak No. | Compounds | TPF (μg/g) | FF (μg/g) | AF (μg/g) |
|---|---|---|---|---|
| 1 | Protocatechuic acid | 861.69 ± 23.72d | 297.66 ± 33.54c | ND * |
| 3 | Coumaric acid | 1354.42 ± 97.94d | 1459.71 ± 94.02c | ND |
| 4 | Kaempferol-3-O-glucoside b | 218.02 ± 34.34d | 4195.15 ± 116.62c | ND |
| 5 | Kaempferol-3-O-rhamnosylhexose b | 131.91 ± 20.82d | 1909.04 ± 78.39c | ND |
| 6 | Dihydroquercetin rhamnoside b | 302.07 ± 1.13c | Trace | ND |
| 7 | Catechin | 486.50 ± 9.92d | 3258.80 ± 119.01c | ND |
| 8 | Isoschaftoside | 645.95 ± 25.38c | 67.65 ± 6.65c | ND |
| 9 | Rutin | 10,177.90 ± 468.53c | 5082.50 ± 200.46d | Trace |
| 10 | Quercetin-3-O-glucoside b | 1274.17 ± 98.72d | 8685.38 ± 179.22c | Trace |
| 11 | Quercetin 3-(6-O-acetyl-beta-glucoside) b | 63.10 ± 7.07d | 189.02 ± 12.70c | ND |
| 12 | Isorhamnetin-3-O-rutinoside | 7072.17 ± 27.93c | 7566.58 ± 503.86c | Trace |
| 13 | Kaempferol-3-O-hexoside b | 44.01 ± 0.05d | 459.90 ± 21.08c | ND |
| 14 | Isorhamnetin-3-O-glucoside | 566.02 ± 17.17d | 7443.42 ± 119.55c | Trace |
| 15 | Cyanidin-3-O-glucoside | 2806.80 ± 159.50 | Trace | 15,800.42 ± 822.41c |
| 16 | Cyanidin-3-O-rutinoside b | 1003.42 ± 347.20 | Trace | 5011.44 ± 282.87c |
| 17 | Peonidin-3-O-rutinoside b | Trace | ND | 684.64 ± 65.97c |
| 18 | Peonidin-3-O-sophoroside-5-glucoside b | Trace | ND | 192.08 ± 18.51c |
| 19 | Delphinidin-3-O-rutinoside b | Trace | ND | 639.78 ± 61.64c |
| 20 | Petunidin-3-O-glucoside b | Trace | ND | 246.67 ± 23.77c |
| Total content | 27,008.15 | 40,614.81 | 22,575.03 | |
| C-Score | T-Score | PMF-Score | CHEM-Score | G-Score | D-Score | ||
|---|---|---|---|---|---|---|---|
| Lipase | Rutin | 4 | 8.3565 | 17.9959 | −13.5442 | −186.0455 | −186.0455 |
| Isorhamnetin-3-O-rutinoside | 5 | 5.7784 | 9.6420 | −18.6390 | −171.0586 | −175.6447 | |
| Cyanidin-3-O-glucoside | 4 | 5.5067 | −3.5225 | −17.2561 | −196.237 | −118.6521 | |
| α-Glucosidase | Rutin | 5 | 7.8139 | −127.513 | −21.6289 | −56.3033 | −155.2835 |
| Isorhamnetin 3-O-rutinoside | 5 | 7.1493 | −106.7521 | −27.5 | −201.3377 | −225.5207 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Jia, Y.; Ma, Y.; Cheng, G.; Cai, S. Phenolic Composition, Antioxidant Properties, and Inhibition toward Digestive Enzymes with Molecular Docking Analysis of Different Fractions from Prinsepia utilis Royle Fruits. Molecules 2018, 23, 3373. https://doi.org/10.3390/molecules23123373
Zhang X, Jia Y, Ma Y, Cheng G, Cai S. Phenolic Composition, Antioxidant Properties, and Inhibition toward Digestive Enzymes with Molecular Docking Analysis of Different Fractions from Prinsepia utilis Royle Fruits. Molecules. 2018; 23(12):3373. https://doi.org/10.3390/molecules23123373
Chicago/Turabian StyleZhang, Xuan, Yijia Jia, Yanli Ma, Guiguang Cheng, and Shengbao Cai. 2018. "Phenolic Composition, Antioxidant Properties, and Inhibition toward Digestive Enzymes with Molecular Docking Analysis of Different Fractions from Prinsepia utilis Royle Fruits" Molecules 23, no. 12: 3373. https://doi.org/10.3390/molecules23123373
APA StyleZhang, X., Jia, Y., Ma, Y., Cheng, G., & Cai, S. (2018). Phenolic Composition, Antioxidant Properties, and Inhibition toward Digestive Enzymes with Molecular Docking Analysis of Different Fractions from Prinsepia utilis Royle Fruits. Molecules, 23(12), 3373. https://doi.org/10.3390/molecules23123373