Effect of Indolic-Amide Melatonin on Blood Cell Population: A Biophysical Gaussian Statistical Analysis

 ,
, 
Abstract
:1. Introduction
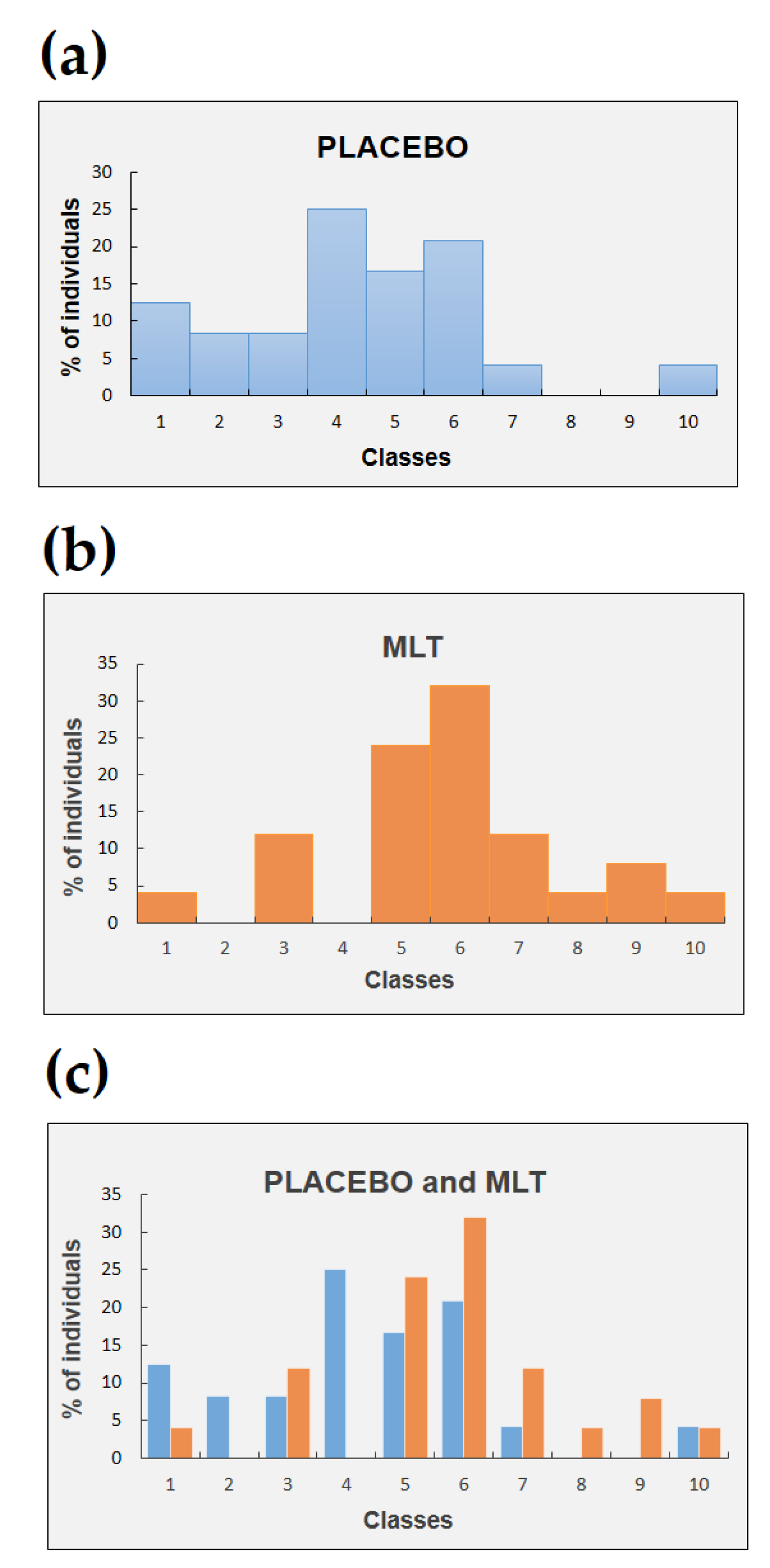
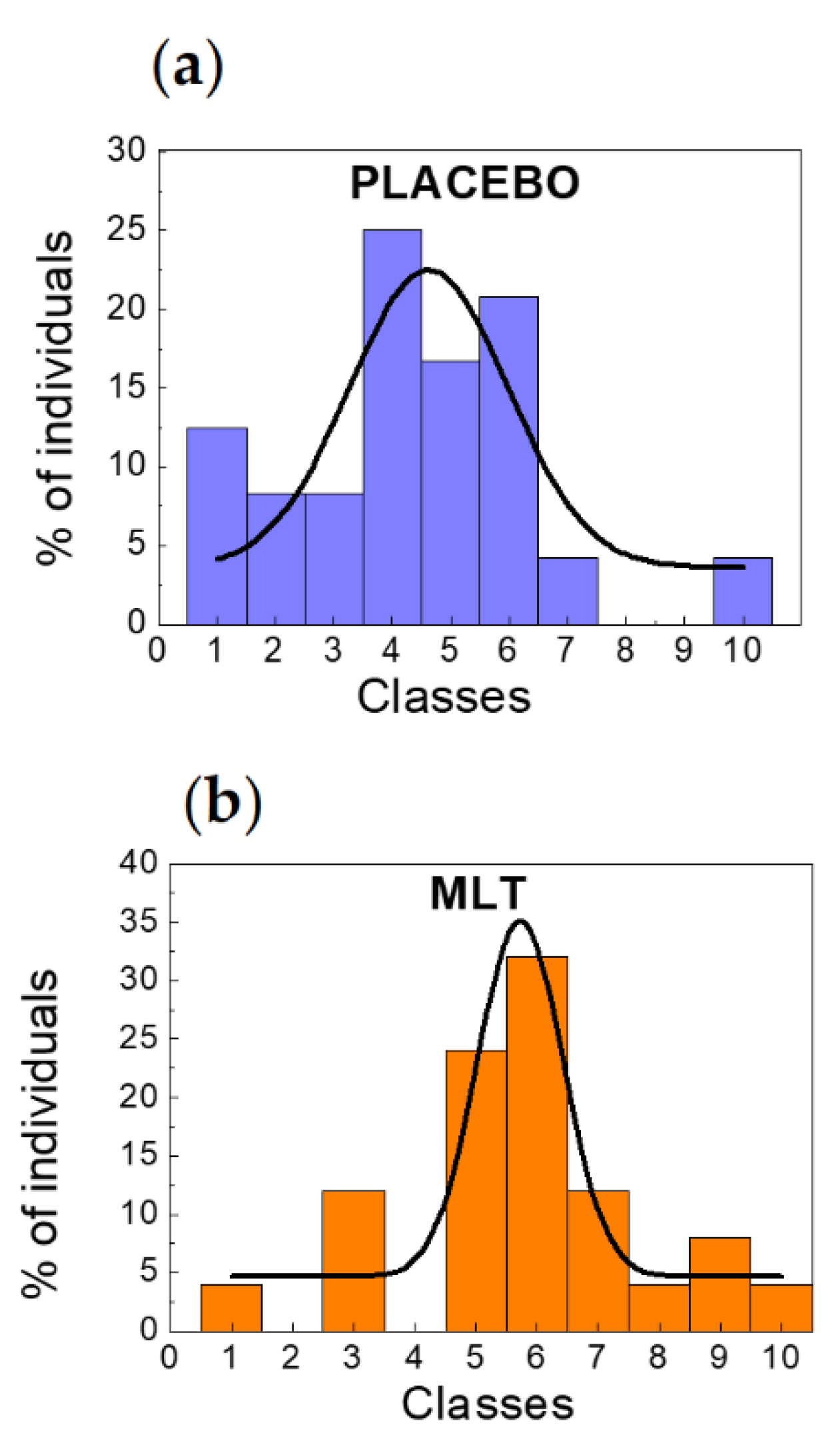
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Tsuchiya, M.; Ross, J. Advantages of external periodic events to the evolution of biochemical oscillatory reactions. Proc. Natl. Acad. Sci. USA 2003, 100, 9691–9695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stringari, C.; Wang, H.; Geyfman, M.; Crosignani, V.; Kumar, V.; Takahashi, J.S.; Andersen, B.; Gratton, E. In vivo single-cell detection of metabolic oscillations in stem cells. Cell Rep. 2015, 10, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, E.A.; Abelson, J.; Lightman, S.L. Cortisol pulsatility and its role in stress regulation and health. Front. Neuroendocrinol. 2004, 25, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.H.; Yang, L.; Qu, Z.; Weiss, J.N. Glycolytic oscillations in isolated rabbit ventricular myocytes. J. Biol. Chem. 2008, 283, 36321–36327. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.M.; Loros, J.J.; Dunlap, J.C. Circadian oscillators: Around the transcription-translation feedback loop and on to output. Trends Biochem. Sci. 2016, 41, 834–846. [Google Scholar] [CrossRef] [PubMed]
- Zivieri, R.; Pacini, N.; Finocchio, G.; Carpentieri, M. Rate of entropy model for irreversible processes in living systems. Sci. Rep. 2017, 7, 9134. [Google Scholar] [CrossRef] [PubMed]
- Pacini, N.; Borziani, F. Cancer stem cell theory and the Warburg effect, two sides of the same coin? Int. J. Mol. Sci. 2014, 15, 8893–8930. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.E.; Dougherty, T.F. The diurnal variation of blood leukocyes in normal and adrenalectomized mice. Endocrinology 1956, 58, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Richens, E.; Brown, K.A.; Collins, A.J.; Bacon, P.A. Circadian rhythms in cell migration in vitro, and its effect on antigen-induced migration inhibition. Clin. Exp. Immunol. 1975, 2, 343–346. [Google Scholar]
- Bertouch, J.V.; Roberts-Thomson, P.J.; Bradley, J. Diurnal variation of lymphocyte subsets identified by monoclonal antibodies. Br. Med. J. (Clin. Res. Ed.) 1983, 286, 1171–1172. [Google Scholar] [CrossRef]
- Ritchie, A.W.; Oswald, I.; Micklem, H.S.; Boyd, J.E.; Elton, R.A.; Jazwinska, E.; James, K. Circadian variation of lymphocyte subpopulations: A study with monoclonal antibodies. Br. Med. J. (Clin. Res. Ed.) 1983, 286, 1773–1775. [Google Scholar] [CrossRef]
- Sennels, H.P.; Jørgensen, H.L.; Hansen, A.L.; Goetze, J.P.; Fahrenkrug, J. Diurnal variation of hematology parameters in healthy young males: The Bispebjerg study of diurnal variations. Scand. J. Clin. Lab. Investig. 2011, 71, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, E.E. Suprachiasmatic nucleus and melatonin: Reciprocal interactions and clinical correlations. Neurology 2008, 71, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Pacini, N.; Borziani, F. Oncostatic-Cytoprotective Effect of Melatonin and Other Bioactive Molecules: A Common Target in Mitochondrial Respiration. Int. J. Mol. Sci. 2016, 17, 341. [Google Scholar] [CrossRef] [PubMed]
- Pacini, N.; Ferrari, A.; Menozzi, M.; Borziani, F. Melatonin enhances the in vitro action of cytochalasin B on globular resistance and osmotic fragility of erythrocytes. Neuro Endocrinol. Lett. 2011, 32, 292–300. [Google Scholar] [PubMed]
- Margheri, M.; Pacini, N.; Tani, A.; Nosi, D.; Squecco, R.; Dama, A.; Masala, E.; Francini, F.; Zecchi-Orlandini, S.; Formigli, L. Combined effects of melatonin and all-trans retinoic acid and somatostatin on breast cancer cell proliferation and death: Molecular basis for the anticancer effect of these molecules. Eur. J. Pharmacol. 2012, 681, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Gil, B.; Moneim, A.E.A.; Ortiz, F.; Shen, Y.-Q.; Soto-Mercado, V.; Mendivil-Perez, M.; Guerra-Librero, A.; Acuña-Castroviejo, D.; Molina-Navarro, M.M.; García-Verdugo, J.M.; et al. Melatonin protects rats from radiotherapy-induced small intestine toxicity. PLoS ONE 2017, 12, e0174474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardinali, D.P.; Pagano, E.S.; Scacchi Bernasconi, P.A.; Reynoso, R.; Scacchi, P. Melatonin and mitochondrial dysfunction in the central nervous system. Horm. Behav. 2013, 63, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Pacini, N.; Borziani, F. Action of melatonin on bone marrow depression induced by cyclophosphamide in acute toxicity phase. Neuro Endocrinol. Lett. 2009, 30, 582–591. [Google Scholar] [PubMed]
- Srinivasan, V.; Pandi-Perumal, S.R.; Brzezinski, A.; Bhatnagar, K.P.; Cardinali, D.P. Melatonin, immune function and cancer. Recent Pat. Endocr. Metab. Immune Drug Discov. 2011, 5, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Fedosov, D.A.; Fornleitner, J.; Gompper, G. Margination of white blood cells in microcapillary flow. Phys. Rev. Lett. 2012, 108, 028104. [Google Scholar] [CrossRef] [PubMed]
- Steffen, P.; Verdier, C.; Wagner, C. Quantification of depletion induced adhesion of red blood cells. Phys. Rev. Lett. 2013, 110, 018102. [Google Scholar] [CrossRef]
- Kaoui, B.; Biros, G.; Misbah, C. Why do red blood cells have asymmetric shapes even in a symmetric flow? Phys. Rev. Lett. 2009, 103, 188101. [Google Scholar] [CrossRef]
- Stroebel, A.M.; Bergner, M.; Reulbach, U.; Biermann, T.; Groemer, T.W.; Klein, I.; Kornhuber, J. Statistical methods for detecting and comparing periodic data and their application to the nycthemeral rhythm of bodily harm: A population based study. J. Circadian Rhythms 2010, 8, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, E.; Brodan, V.; Brodanová, M.; Rysánek, K. Metabolic reflection of sleep deprivation. Act. Nerv. Super 1969, 11, 165–174. [Google Scholar]
- Ackermann, K.; Revell, V.L.; Lao, O.; Rombouts, E.J.; Skene, D.J.; Kayser, M. Diurnal rhythms in blood cell populations and the effect of acute sleep deprivation in healthy young men. Sleep 2012, 35, 933–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boudjeltia, K.Z.; Faraut, B.; Stenuit, P.; Esposito, M.J.; Dyzma, M.; Brohée, D.; Ducobu, J.; Vanhaeverbeek, M.; Kerkhofs, M. Sleep restriction increases white blood cells, mainly neutrophil count, in young healthy men: A pilot study. Vasc. Health Risk Manag. 2008, 4, 1467–1470. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.D.; Pollak, A.; Akman, S.A.; Bachur, N.R. Diurnal variation of circulating human myeloid progenitor cells. Exp. Hematol. 1980, 7, 954–960. [Google Scholar]
- Kang, J.C.; Ahn, M.; Kim, Y.S.; Moon, C.; Lee, Y.; Wie, M.B.; Lee, Y.J.; Shin, T. Melatonin ameliorates autoimmune encephalomyelitis through suppression of intercellular adhesion molecule-1. J. Vet. Sci. 2001, 2, 85–89. [Google Scholar] [PubMed]
- Wen, J.; Ariyannur, P.S.; Ribeiro, R.; Tanaka, M.; Moffett, J.R.; Kirmani, B.F.; Namboodiri, A.M.; Zhang, Y. Efficacy of N-Acetylserotonin and Melatonin in the EAE Model of Multiple Sclerosis. J. Neuroimmune Pharmacol. 2016, 4, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu, C.S.; Hilliard, B.; Ventura, E.; Rostami, A. Luzindole, a melatonin receptor antagonist, suppresses experimental autoimmune encephalomyelitis. Pathobiology 1997, 65, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.L.; Li, Y.J.; Hu, B.B.; Wang, H.; Hu, B. Melatonin regulates the rhythmic migration of neutrophils in live zebrafish. J. Pineal Res. 2015, 58, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Černyšiov, V.; Mauricas, M.; Girkontaite, I. Melatonin inhibits granulocyte adhesion to ICAM via MT3/QR2 and MT2 receptors. Int. Immunol. 2015, 12, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Lotufo, C.M.; Yamashita, C.E.; Farsky, S.H.; Markus, R.P. Melatonin effect on endothelial cells reduces vascular permeability increase induced by leukotriene B4. Eur. J. Pharmacol. 2006, 534, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Jockers, R.; Maurice, P.; Boutin, J.A.; Delagrange, P. Melatonin receptors, heterodimerization, signal transduction and binding sites: What’s new? Br. J. Pharmacol. 2008, 154, 1182–1195. [Google Scholar] [CrossRef] [PubMed]
- Gabel, V.; Reichert, C.F.; Maire, M.; Schmidt, C.; Schlangen, L.J.M.; Kolodyazhniy, V.; Garbazza, C.; Cajochen, C.; Viola, A.U. Differential impact in young and older individuals of blue-enriched white light on circadian physiology and alertness during sustained wakefulness. Sci. Rep. 2017, 7, 7620. [Google Scholar] [CrossRef] [PubMed]
- Torres-Farfan, C.; Richter, H.G.; Rojas-García, P.; Vergara, M.; Forcelledo, M.L.; Valladares, L.E.; Torrealba, F.; Valenzuela, G.J.; Serón-Ferré, M. mt1 Melatonin receptor in the primate adrenal gland: Inhibition of adrenocorticotropin-stimulated cortisol production by melatonin. J. Clin. Endocrinol. Metab. 2003, 88, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Campino, C.; Valenzuela, F.; Arteaga, E.; Torres-Farfán, C.; Trucco, C.; Velasco, A.; Guzmán, S.; Serón-Ferré, M. Melatonin reduces cortisol response to ACTH in humans. Rev. Med. Chile 2008, 136, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Lv, J.; Li, W.; Zhang, P.; Mao, C.; Xu, Z. Exogenous melatonin reduced blood pressure in late-term ovine fetus via MT1/MT2 receptor pathways. Reprod. Biol. 2016, 16, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Campino, C.; Valenzuela, F.J.; Torres-Farfan, C.; Reynolds, H.E.; Abarzua-Catalan, L.; Arteaga, E.; Trucco, C.; Guzmán, S.; Valenzuela, GJ.; Seron-Ferre, M. Melatonin exerts direct inhibitory actions on ACTH responses in the human adrenal gland. Horm. Metab. Res. 2011, 43, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Presman, D.M.; Levi, V.; Pignataro, O.P.; Pecci, A. Melatonin inhibits glucocorticoid-dependent GR-TIF2 interaction in newborn hamster kidney (BHK) cells. Mol. Cell. Endocrinol. 2012, 349, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Jilma, B.; Hergovich, N.; Stohlawetz, P.; Eichler, H.G.; Bauer, P.; Wagner, O.F. Circadian variation of granulocyte colony stimulating factor levels in man. Br. J. Haematol. 1999, 106, 368–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perdiz, P.; Wacher, N.; Laredo-Sánchez, F.; Halabe Cherem, J.; Lifshitz, A. Circadian variation of human acute phase response. Arch. Med. Res. 1996, 27, 157–163. [Google Scholar] [PubMed]
- Dubocovich, M.L.; Delagrange, P.; Krause, D.N.; Sugden, D.; Cardinali, D.P.; Olcese, J. International union of basic and clinical pharmacology. LXXV. Nomenclature, classification, and pharmacology of G protein-coupled melatonin receptors. Pharmacol. Rev. 2010, 62, 343–380. [Google Scholar] [CrossRef] [PubMed]
- Lacoste, B.; Angeloni, D.; Dominguez-Lopez, S.; Calderoni, S.; Mauro, A.; Fraschini, F.; Descarries, L.; Gobbi, G. Anatomical and cellular localization of melatonin MT1 and MT2 receptors in the adult rat brain. J. Pineal Res. 2015, 58, 397–417. [Google Scholar]
- Lopez-Canul, M.; Palazzo, E.; Dominguez-Lopez, S.; Luongo, L.; Lacoste, B.; Comai, S.; Angeloni, D.; Fraschini, F.; Boccella, S.; Spadoni, G. Selective melatonin MT2 receptor ligands relieve neuropathic pain through modulation of brainstem descending antinociceptive pathways. Pain 2015, 156, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Zivieri, R.; Pacini, N. Is an entropy-based approach suitable for an understanding of the metabolic pathways of fermentation and respiration? Entropy 2017, 19, 662. [Google Scholar] [CrossRef]
- Halberg, F.; Cornélissen, G.; Katinas, G.; Syutkina, E.V.; Sothern, R.B.; Zaslavskaya, R.; Halberg, F.; Watanabe, Y.; Schwartzkopff, O.; Otsuka, K.; et al. Transdisciplinary unifying implications of circadian findings in the 1950s. J. Circadian Rhythms 2003, 1, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheiermann, C.; Kunisaki, Y.; Frenette, P.S. Circadian control of the immune system. Nat. Rev. Immunol. 2013, 13, 13–198. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Age | (44.7 ± 3.6) years |
| Sex | 54 f - 49 m |
| BMI | 24.3 kg/m2 |
| Time | 9–9.30 p.m. | 9–9.30 a.m. | % Variations (t-Student) |
|---|---|---|---|
| RBCs and PLTs | Average value | Average value | |
| RBCs (106/µL) | 4.796 | 4.883 | +1.81% (p > 0.05) |
| HG (g/dL) | 13.620 | 13.810 | +1.40% (p > 0.05) |
| HCT (%) | 43.320 | 43.620 | +0.69% (p > 0.05) |
| MCV (fL) | 91.620 | 90.140 | −1.62% (p > 0.05) |
| PLT (103/µL) | 261.420 | 255.200 | −2.38% (p > 0.05) |
| WBCs | Average value | Average value | |
| Leukocytes (103/µL) | 8.944 | 7.786 | −12.95% (p < 0.001) |
| Lymphocytes | 3.018 | 2.675 | −11.37% (p < 0.05) |
| Monocytes | 0.602 | 0.521 | −13.46% (p > 0.05) |
| Neutrophils | 5.056 | 4.341 | −14.14% (p < 0.001) |
| Eosinophils | 0.200 | 0.186 | −7.00% (p > 0.05) |
| Basophils | 0.068 | 0.063 | −7.35% (p > 0.05) |
| Time | 9–9.30 p.m. | 9–9.30 a.m. | % Variations (t-Student) |
|---|---|---|---|
| RBCs and PLTs | Average value | Average value | |
| RBCs (106/µL) | 4.770 | 4.848 | +1.64% (p > 0.05) |
| HG (g/dL) | 14.540 | 14.580 | +0.28% (p > 0.05) |
| HCT% | 42.130 | 42.580 | +1.07% (p > 0.05) |
| MCV (fL) | 88.770 | 88.420 | −0.39% (p > 0.05) |
| PLT (103/µL) | 248.120 | 236.440 | −4.71% (p < 0.05) |
| WBCs | Average value | Average value | |
| Leukocytes (103/µL) | 8.187 | 6.401 | −21.81% (p < 0.001) |
| Lymphocytes | 3.043 | 2.405 | −20.97% (p < 0.001) |
| Monocytes | 0.438 | 0.345 | −21.23% (p < 0.001) |
| Neutrophils | 4.393 | 3.366 | −23.38% (p < 0.001) |
| Eosinophils | 0.246 | 0.234 | −4.88% (p > 0.05) |
| Basophils | 0.067 | 0.051 | −23.88% (p > 0.05) |
| Group | Untreated | MLT | % Variations (t-Student) |
|---|---|---|---|
| RBCs and PLTs | Average value | Average value | |
| RBCs (106/µL) | 4.864 | 4.848 | −0.33% |
| HG (g/dL) | 14.598 | 14.580 | −0.12% |
| HCT% | 42.622 | 42.580 | −0.10% |
| MCV (fL) | 88.241 | 88.420 | +0.20% |
| PLT (103/µL) | 242.963 | 236.440 | −2.64% |
| WBCs | Average value | Average value | |
| Leukocytes (103/µL) | 8.149 | 6.401 | −21.45% (p < 0.001) |
| Lymphocytes | 2.662 | 2.405 | −9.65% (p < 0.001) |
| Monocytes | 0.448 | 0.345 | −22.99% (p < 0.001) |
| Neutrophils | 4.727 | 3.366 | −28.79% (p < 0.001) |
| Eosinophils | 0.258 | 0.234 | −9.30% (p > 0.05) |
| Basophils | 0.054 | 0.051 | −5.56% (p > 0.05) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zivieri, R.; Borziani, F.; Strazzanti, A.; Fragomeni, A.; Pacini, N. Effect of Indolic-Amide Melatonin on Blood Cell Population: A Biophysical Gaussian Statistical Analysis. Molecules 2018, 23, 1378. https://doi.org/10.3390/molecules23061378
Zivieri R, Borziani F, Strazzanti A, Fragomeni A, Pacini N. Effect of Indolic-Amide Melatonin on Blood Cell Population: A Biophysical Gaussian Statistical Analysis. Molecules. 2018; 23(6):1378. https://doi.org/10.3390/molecules23061378
Chicago/Turabian StyleZivieri, Roberto, Fabio Borziani, Angela Strazzanti, Angela Fragomeni, and Nicola Pacini. 2018. "Effect of Indolic-Amide Melatonin on Blood Cell Population: A Biophysical Gaussian Statistical Analysis" Molecules 23, no. 6: 1378. https://doi.org/10.3390/molecules23061378
APA StyleZivieri, R., Borziani, F., Strazzanti, A., Fragomeni, A., & Pacini, N. (2018). Effect of Indolic-Amide Melatonin on Blood Cell Population: A Biophysical Gaussian Statistical Analysis. Molecules, 23(6), 1378. https://doi.org/10.3390/molecules23061378