Metabolomic Approach for Discrimination of Cultivation Age and Ripening Stage in Ginseng Berry Using Gas Chromatography-Mass Spectrometry


 and
and
Abstract
:1. Introduction
2. Results and Discussion
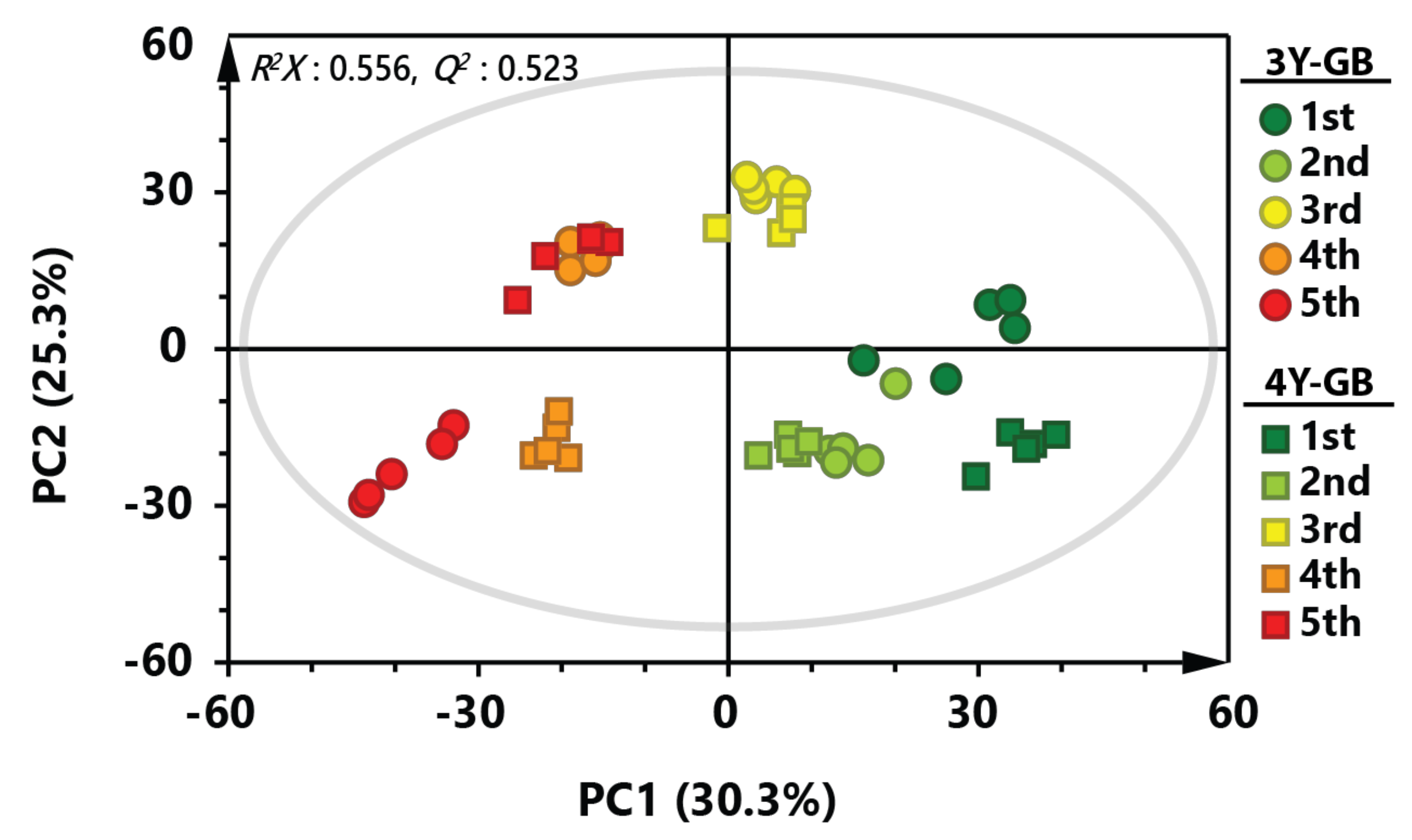
2.1. Metabolic Profiling of GB with Different Cultivation Ages and Ripening Stages
2.2. Metabolic Changes of GB by Ripening Stage
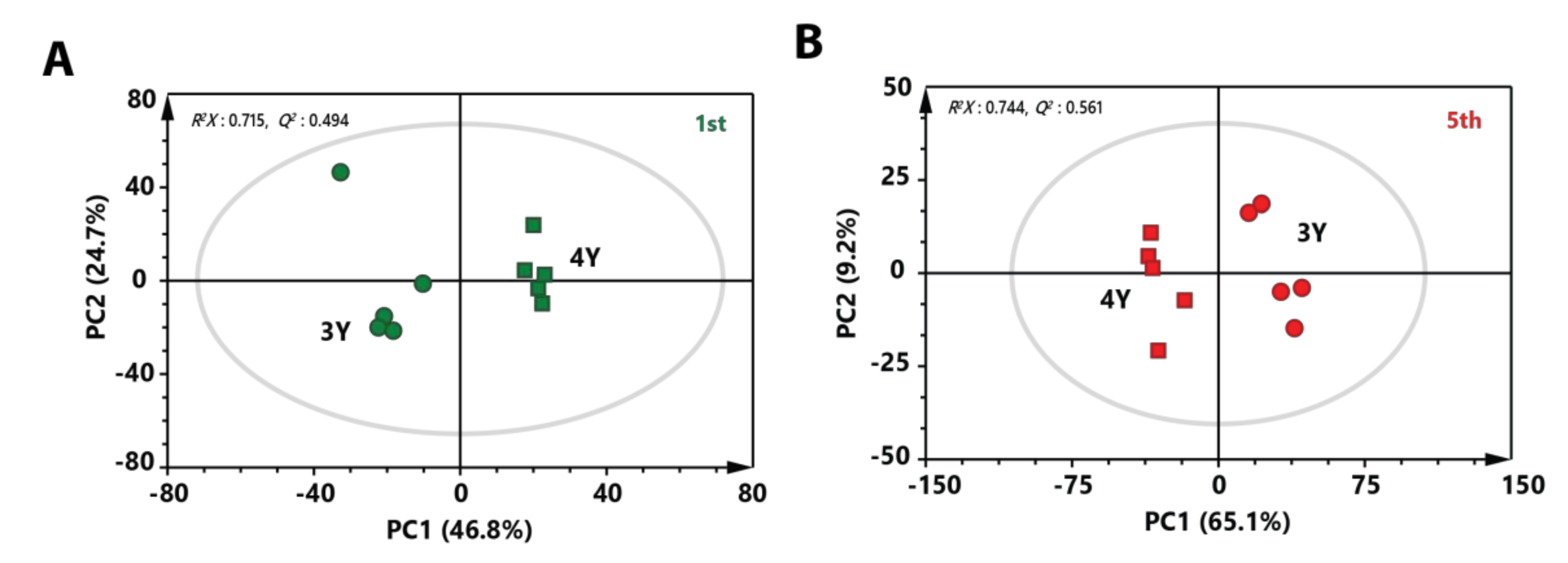
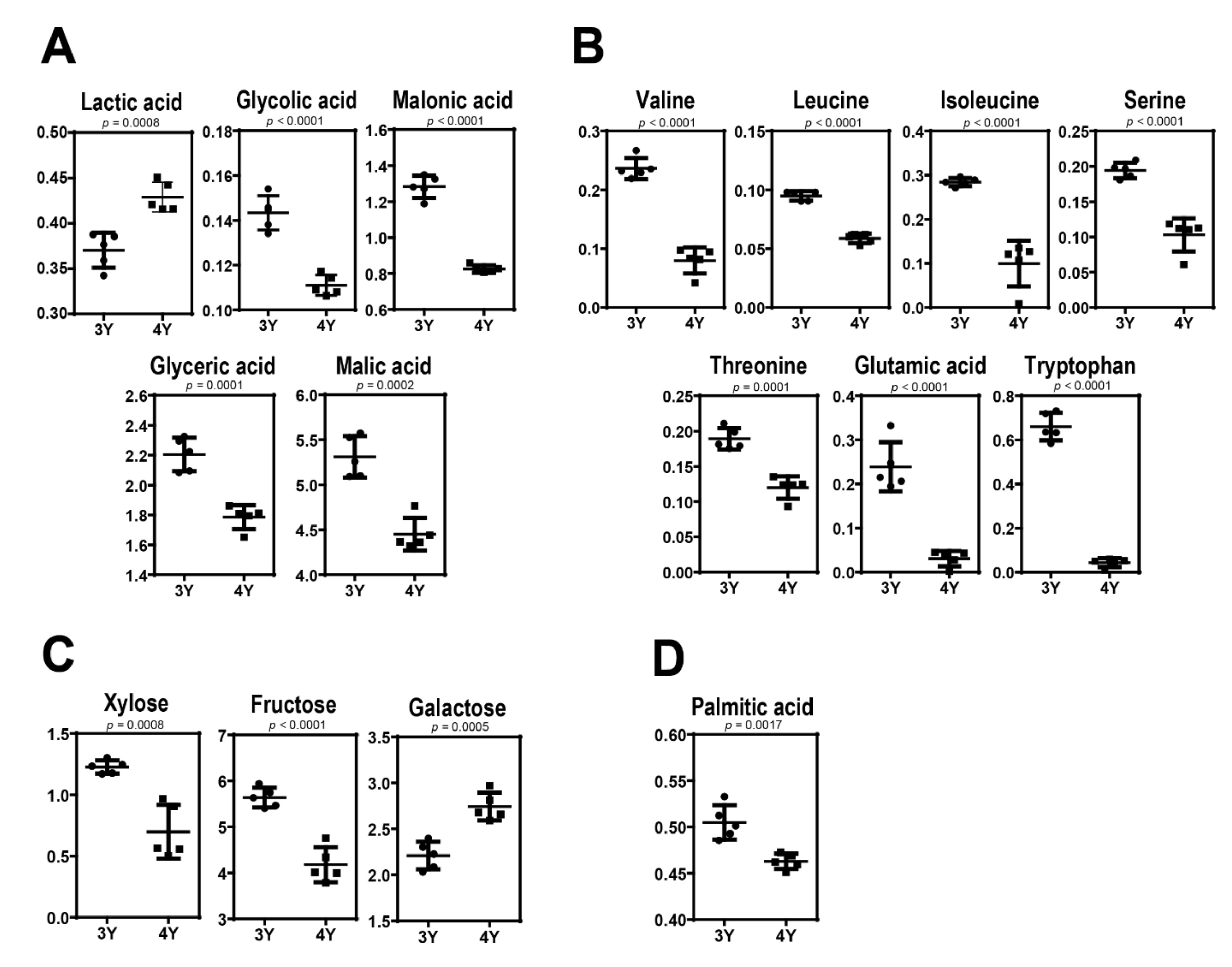
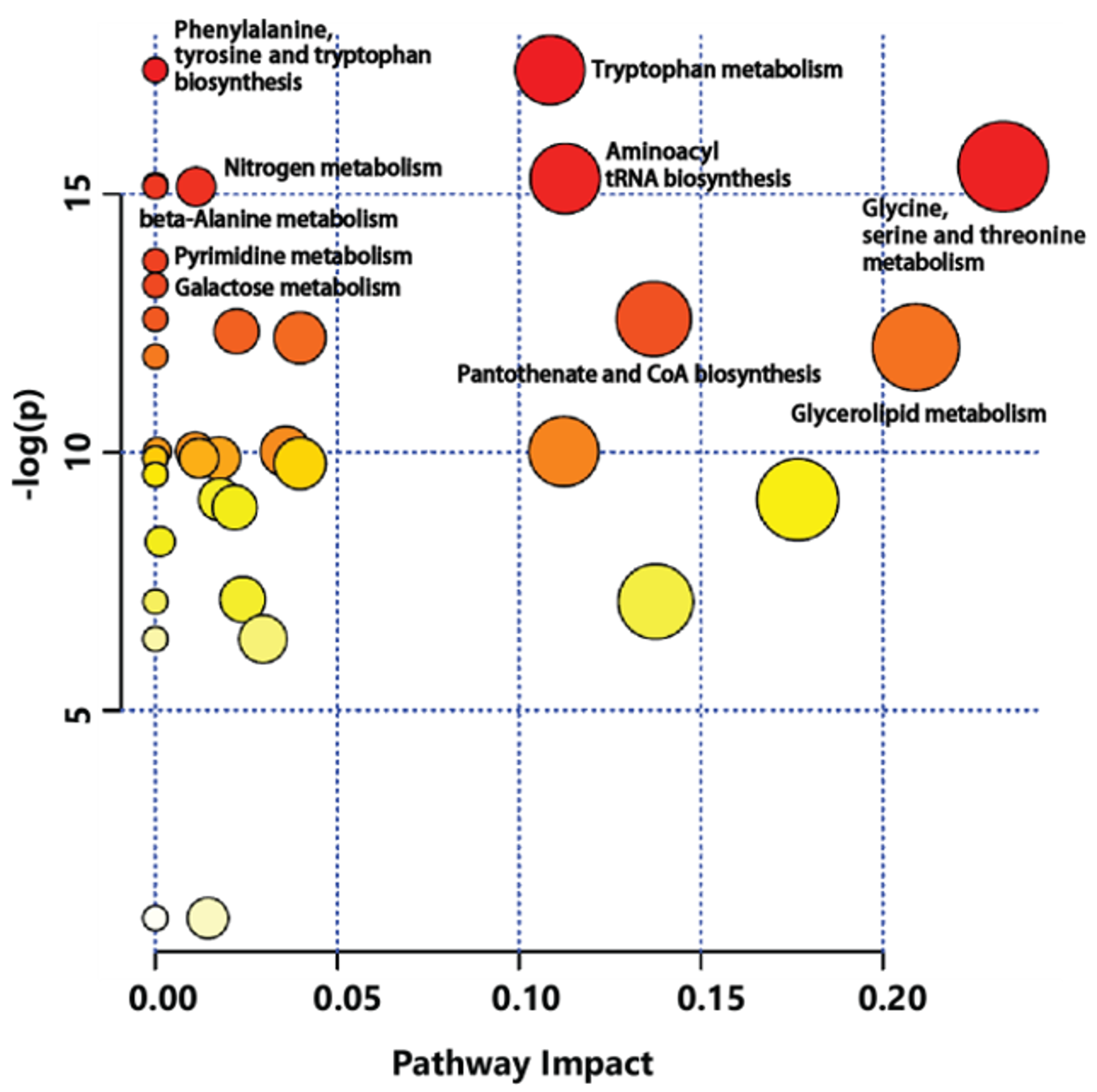
2.3. Metabolic Differences Between Three-Year-Old GB and Four-Year-Old GB
2.4. Classification of GB by Random Forest Machine Learning Algorithm
3. Materials and Methods
3.1. Plant Materials and Preprocessing
3.2. Sample Derivatization
3.3. GC-MS Analysis
3.4. Data Processing
3.5. Random Forest Machine Learning Algorithm
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jeon, B.H.; Kim, C.S.; Park, K.S.; Lee, J.W.; Park, J.B.; Kim, K.J.; Kim, S.H.; Chang, S.J.; Nam, K.Y. Effect of Korea red ginseng on the blood pressure in conscious hypertensive rats. Gen. Pharmacol. 2000, 35, 135–141. [Google Scholar] [CrossRef]
- Cho, W.C.; Chung, W.S.; Lee, S.K.; Leung, A.W.; Cheng, C.H.; Yue, K.K. Ginsenoside Re of Panax ginseng possesses significant antioxidant and antihyperlipidemic efficacies in streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 2006, 550, 173–179. [Google Scholar] [CrossRef]
- Kim, D.Y.; Yang, W.M. Panax ginseng ameliorates airway inflammation in an ovalbumin-sensitized mouse allergic asthma model. J. Ethnopharmacol. 2011, 136, 230–235. [Google Scholar] [CrossRef]
- Helmes, S. Cancer prevention and therapeutics: Panax ginseng. Altern. Med. Rev. 2004, 9, 259–275. [Google Scholar]
- Ko, S.K.; Bae, H.M.; Cho, O.S.; Im, B.O.; Chung, S.H.; Lee, B.Y. Analysis of ginsenoside composition of ginseng berry and seed. Food Sci. Biotechnol. 2008, 17, 1379–1382. [Google Scholar]
- Wang, C.Z.; Wu, J.A.; McEntee, E.; Yuan, C.S. Saponins composition in American ginseng leaf and berry assayed by high-performance liquid chromatography. J. Agric. Food Chem. 2006, 54, 2261–2266. [Google Scholar] [CrossRef]
- Kim, Y.K.; Yoo, D.S.; Xu, H.; Park, N.I.; Kim, H.H.; Choi, J.E.; Park, S.U. Ginsenoside content of berries and roots of three typical Korean ginseng (Panax ginseng) cultivars. Nat. Prod. Commun. 2009, 4, 903–906. [Google Scholar] [CrossRef]
- Chen, C.F.; Chiou, W.F.; Zhang, J.T. Comparison of the pharmacological effects of Panax ginseng and Panax quinquefolium. Acta Pharmacol. Sin. 2008, 29, 1103–1108. [Google Scholar] [CrossRef]
- Li, F.Y. The origin and classification of Chinese ginseng and American ginseng. In Chinese Ginseng and American Ginseng, 1st ed.; China Agricultural Scientech Press: Beijing, China, 2002; pp. 1–20. [Google Scholar]
- Lee, S.Y.; Kim, Y.K.; Park, N.I.; Kim, C.S.; Lee, C.Y.; Park, S.U. Chemical constituents and biological activities of the berry of Panax ginseng. J. Med. Plant Res. 2010, 4, 349–353. [Google Scholar]
- Dey, L.; Xie, J.; Wang, A.; Wu, J.; Maleckar, S.; Yuan, C.S. Anti-hyperglycemic effects of ginseng: Comparison between root and berry. Phytomedicine 2003, 10, 600–605. [Google Scholar] [CrossRef]
- Choi, H.S.; Kim, S.; Kim, M.J.; Kim, M.S.; Kim, J.; Park, C.W.; Seo, D.; Shin, S.S.; Oh, S.W. Efficacy and safety of Panax ginseng berry extract on glycemic control: A 12-wk randomized, double-blind, and placebo-controlled clinical trial. J. Ginseng Res. 2018, 42, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jeong, J.; Alves, A.C.; Han, S.T.; In, G.; Kim, E.H.; Jeong, W.S.; Hong, Y.S. Metabolomic understanding of intrinsic physiology in Panax ginseng during whole growing seasons. J. Ginseng Res. 2019, 43, 654–665. [Google Scholar] [CrossRef]
- Song, H.H.; Kim, D.Y.; Woo, S.; Lee, H.K.; Oh, S.R. An approach for simultaneous determination for geographical origins of Korean Panax ginseng by UPLC-QTOF/MS coupled with OPLS-DA models. J. Ginseng Res. 2013, 37, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, Y.; Wang, Y.; Abozeid, A.; Zu, Y.G.; Tang, Z.H. The integration of GC–MS and LC–MS to assay the metabolomics profiling in Panax ginseng and Panax quinquefolius reveals a tissue-and species-specific connectivity of primary metabolites and ginsenosides accumulation. J. Pharm. Biomed. Anal. 2017, 135, 176–185. [Google Scholar] [CrossRef]
- Kim, S.W.; Gupta, R.; Lee, S.H.; Min, C.W.; Agrawal, G.K.; Rakwal, R.; Kim, J.B.; Jo, I.H.; Park, S.Y.; Kim, J.K.; et al. An integrated biochemical, proteomics, and metabolomics approach for supporting medicinal value of Panax ginseng fruits. Front. Plant Sci. 2016, 7, 994. [Google Scholar] [CrossRef]
- Chung, I.M.; Kim, J.W.; Seguin, P.; Jun, Y.M.; Kim, S.H. Ginsenosides and phenolics in fresh and processed Korean ginseng (Panax ginseng CA Meyer): Effects of cultivation location, year, and storage period. Food Chem. 2012, 130, 73–83. [Google Scholar] [CrossRef]
- Chung, I.M.; Lim, J.J.; Ahn, M.S.; Jeong, H.N.; An, T.J.; Kim, S.H. Comparative phenolic compound profiles and antioxidative activity of the fruit, leaves, and roots of Korean ginseng (Panax ginseng Meyer) according to cultivation years. J. Ginseng Res. 2016, 40, 68–75. [Google Scholar] [CrossRef]
- Song, S.Y.; Park, D.H.; Seo, S.W.; Park, K.M.; Bae, C.S.; Son, H.S.; Kim, H.G.; Lee, J.H.; Yoon, G.; Shim, J.H.; et al. Effects of harvest time on phytochemical constituents and biological activities of Panax ginseng berry extracts. Molecules 2019, 24, 3343. [Google Scholar] [CrossRef]
- Kim, N.; Kim, K.; Choi, B.Y.; Lee, D.; Shin, Y.S.; Bang, K.H.; Cha, S.W.; Lee, J.W.; Choi, H.K.; Jang, D.S.; et al. Metabolomic approach for age discrimination of Panax ginseng using UPLC-Q-Tof MS. J. Agric. Food Chem. 2011, 59, 10435–10441. [Google Scholar] [CrossRef]
- Lee, M.Y.; Seo, H.S.; Singh, D.; Lee, S.J.; Lee, C.H. Unraveling dynamic metabolomes underlying different maturation stages of berries harvested from Panax ginseng. J. Ginseng Res. 2019, in press. [Google Scholar] [CrossRef]
- Yang, S.O.; Park, H.R.; Sohn, E.S.; Lee, S.W.; Kim, H.D.; Kim, Y.C.; Kim, K.H.; Na, S.W.; Choi, H.K.; Arasu, M.V.; et al. Classification of ginseng berry (Panax ginseng CA MEYER) extract using 1 H NMR spectroscopy and its inhibition of lipid accumulation in 3 T3-L1 cells. BMC Complement. Altern. Med. 2014, 14, 455. [Google Scholar] [CrossRef] [PubMed]
- Doerfler, H.; Lyon, D.; Nägele, T.; Sun, X.; Fragner, L.; Hadacek, F.; Egelhofer, V.; Weckwerth, W. Granger causality in integrated GC–MS and LC–MS metabolomics data reveals the interface of primary and secondary metabolism. Metabolomics 2013, 9, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Schluter, C.; Punja, Z.K. Floral biology and seed production in cultivated North American ginseng (Panax quinquefolius). J. Am. Soc. Hortic. Sci. 2000, 125, 567–575. [Google Scholar] [CrossRef]
- Osorio, S.; Alba, R.; Nikoloski, Z.; Kochevenko, A.; Fernie, A.R.; Giovannoni, J.J. Integrative comparative analyses of transcript and metabolite profiles from pepper and tomato ripening and development stages uncovers species-specific patterns of network regulatory behavior. Plant Physiol. 2012, 159, 1713–1729. [Google Scholar] [CrossRef] [PubMed]
- Redgwell, R.J.; Fischer, M.; Kendal, E.; MacRae, E.A. Galactose loss and fruit ripening: High-molecular-weight arabinogalactans in the pectic polysaccharides of fruit cell walls. Planta 1997, 203, 174–181. [Google Scholar] [CrossRef]
- Oikawa, A.; Otsuka, T.; Nakabayashi, R.; Jikumaru, Y.; Isuzugawa, K.; Murayama, H.; Satio, K.; Shiratake, K. Metabolic profiling of developing pear fruits reveals dynamic variation in primary and secondary metabolites, including plant hormones. PLoS ONE 2015, 10, e0131408. [Google Scholar] [CrossRef]
- Mastrangelo, A.; Ferrarini, A.; Rey-Stolle, F.; García, A.; Barbas, C. From sample treatment to biomarker discovery: A tutorial for untargeted metabolomics based on GC-(EI)-Q-MS. Anal. Chim. Acta 2015, 900, 21–35. [Google Scholar] [CrossRef]
- Park, S.E.; Seo, S.H.; Kim, E.J.; Byun, S.; Na, C.S.; Son, H.S. Changes of microbial community and metabolite in kimchi inoculated with different microbial community starters. Food Chem. 2019, 274, 558–565. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Metabolites | Three-Year-Old | Trend 1) | Four-Year-Old | Trend | ||
|---|---|---|---|---|---|---|---|
| First Harvest | Fifth Harvest | First Harvest | Fifth Harvest | ||||
| Organicacids | Lactic Acid | 0.540 ± 0.062 | 0.370 ± 0.019 | ↓(***) | 0.877 ± 0.052 | 0.429 ± 0.016 | ↓(***) |
| Glycolic acid | 0.150 ± 0.019 | 0.143 ± 0.008 | ↓(NS) | 0.138 ± 0.009 | 0.111 ± 0.005 | ↓(***) | |
| Malonic acid | 1.317 ± 0.146 | 1.283 ± 0.062 | ↓(NS) | 1.249 ± 0.083 | 0.825 ± 0.021 | ↓(***) | |
| Succinic acid | 0.933 ± 0.094 | 0.626 ± 0.077 | ↓(***) | 0.958 ± 0.049 | 0.667 ± 0.061 | ↓(***) | |
| Glyceric acid | 2.160 ± 0.169 | 2.204 ± 0.112 | ↑(NS) | 1.924 ± 0.085 | 1.785 ± 0.079 | ↓(*) | |
| Malic acid | 3.316 ± 0.287 | 5.310 ± 0.229 | ↑(***) | 3.278 ± 0.116 | 4.451 ± 0.181 | ↑(***) | |
| Amino acids | Valine | 0.223 ± 0.053 | 0.237 ± 0.018 | ↑(NS) | 0.319 ± 0.058 | 0.080 ± 0.022 | ↓(***) |
| Leucine | 0.066 ± 0.018 | 0.095 ± 0.004 | ↑(NS) | 0.080 ± 0.019 | 0.059 ± 0.004 | ↓(NS) | |
| Isoleucine | 0.322 ± 0.051 | 0.284 ± 0.009 | ↓(NS) | 0.242 ± 0.099 | 0.100 ± 0.052 | ↓(*) | |
| Serine | 0.252 ± 0.036 | 0.194 ± 0.011 | ↓(**) | 0.162 ± 0.081 | 0.103 ± 0.024 | ↓(NS) | |
| Threonine | 0.195 ± 0.039 | 0.189 ± 0.015 | ↓(NS) | 0.108 ± 0.044 | 0.120 ± 0.016 | ↑(NS) | |
| Glutamic acid | 0.337 ± 0.064 | 0.239 ± 0.056 | ↓(*) | 0.531 ± 0.046 | 0.031 ± 0.018 | ↓(***) | |
| Tryptophan | 0.361 ± 0.093 | 0.661 ± 0.063 | ↑(***) | 0.581 ± 0.067 | 0.043 ± 0.020 | ↓(***) | |
| Sugars & Sugar derivatives | Xylose | 0.925 ± 0.285 | 1.225 ± 0.055 | ↑(*) | 0.587 ± 0.254 | 0.698 ± 0.218 | ↑(*) |
| Fructose | 3.577 ± 0.285 | 5.635 ± 0.216 | ↑(***) | 2.577 ± 0.163 | 4.176 ± 0.379 | ↑(***) | |
| Glucose | 4.859 ± 0.515 | 9.367 ± 0.501 | ↑(***) | 4.094 ± 0.257 | 7.163 ± 0.411 | ↑(***) | |
| Galactose | 1.626 ± 0.234 | 2.209 ± 0.150 | ↑(**) | 1.054 ± 0.021 | 2.742 ± 0.151 | ↑(***) | |
| Glycerol | 1.983 ± 0.158 | 2.505 ± 0.071 | ↑(***) | 2.411 ± 0.052 | 2.032 ± 0.090 | ↓(***) | |
| Myo-inositol | 1.774 ± 0.377 | 1.290 ± 0.119 | ↓(NS) | 1.111 ± 0.117 | 2.290 ± 0.158 | ↑(***) | |
| Fatty acid | Palmitic acid | 0.507 ± 0.085 | 0.505 ± 0.019 | ↓(NS) | 0.521 ± 0.069 | 0.463 ± 0.008 | ↓(NS) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-E.; Seo, S.-H.; Kim, E.-J.; Park, D.-H.; Park, K.-M.; Cho, S.-S.; Son, H.-S. Metabolomic Approach for Discrimination of Cultivation Age and Ripening Stage in Ginseng Berry Using Gas Chromatography-Mass Spectrometry. Molecules 2019, 24, 3837. https://doi.org/10.3390/molecules24213837
Park S-E, Seo S-H, Kim E-J, Park D-H, Park K-M, Cho S-S, Son H-S. Metabolomic Approach for Discrimination of Cultivation Age and Ripening Stage in Ginseng Berry Using Gas Chromatography-Mass Spectrometry. Molecules. 2019; 24(21):3837. https://doi.org/10.3390/molecules24213837
Chicago/Turabian StylePark, Seong-Eun, Seung-Ho Seo, Eun-Ju Kim, Dae-Hun Park, Kyung-Mok Park, Seung-Sik Cho, and Hong-Seok Son. 2019. "Metabolomic Approach for Discrimination of Cultivation Age and Ripening Stage in Ginseng Berry Using Gas Chromatography-Mass Spectrometry" Molecules 24, no. 21: 3837. https://doi.org/10.3390/molecules24213837
APA StylePark, S. -E., Seo, S. -H., Kim, E. -J., Park, D. -H., Park, K. -M., Cho, S. -S., & Son, H. -S. (2019). Metabolomic Approach for Discrimination of Cultivation Age and Ripening Stage in Ginseng Berry Using Gas Chromatography-Mass Spectrometry. Molecules, 24(21), 3837. https://doi.org/10.3390/molecules24213837