Identification of the Pol Gene as a Species-Specific Diagnostic Marker for Qualitative and Quantitative PCR Detection of Tricholoma matsutake



Abstract
:1. Introduction
2. Results and Discussion
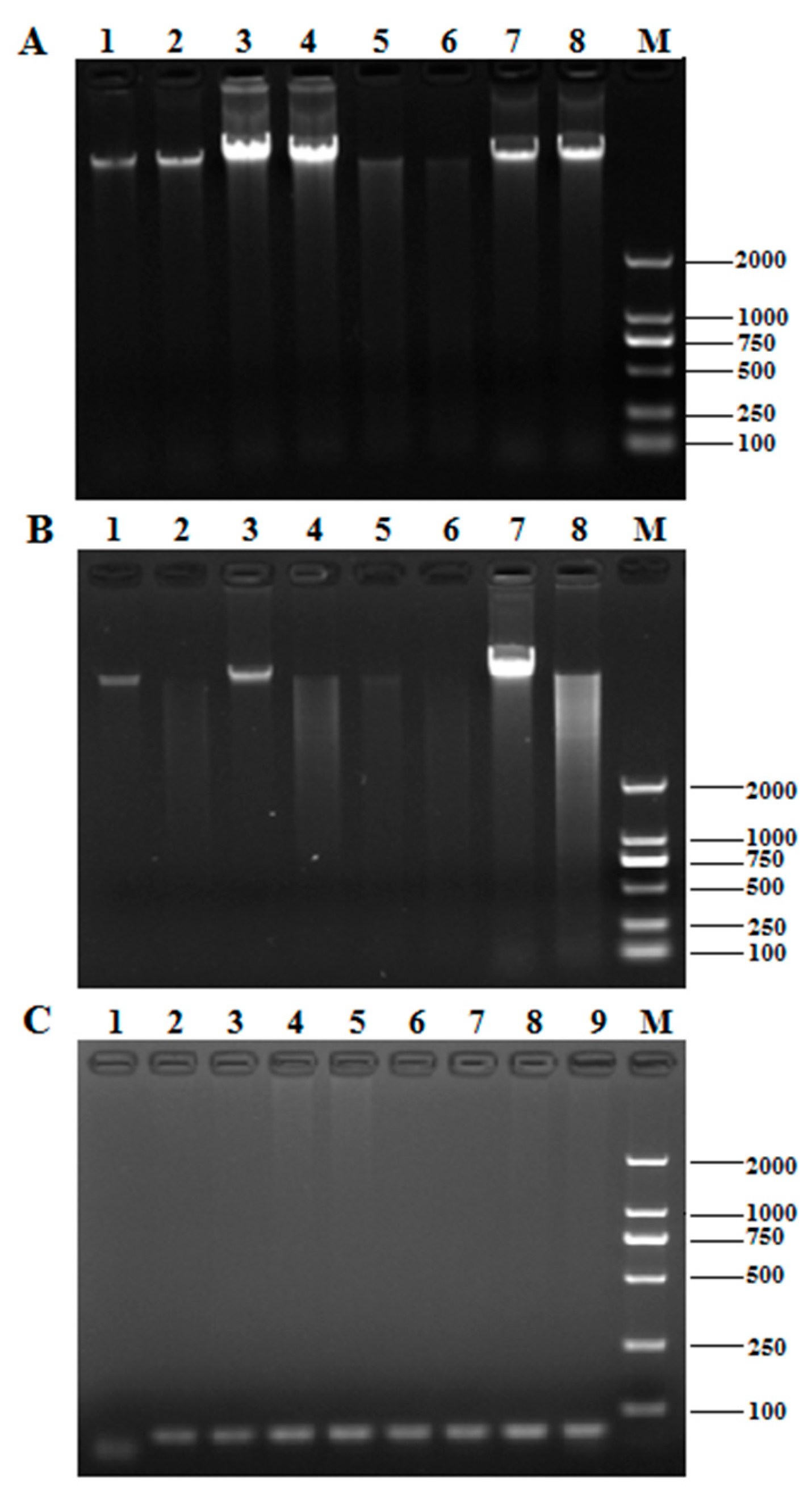
2.1. Comparison of DNA Extraction Methods
2.2. Species-Specific Diagnostic Marker of T. Matsutake
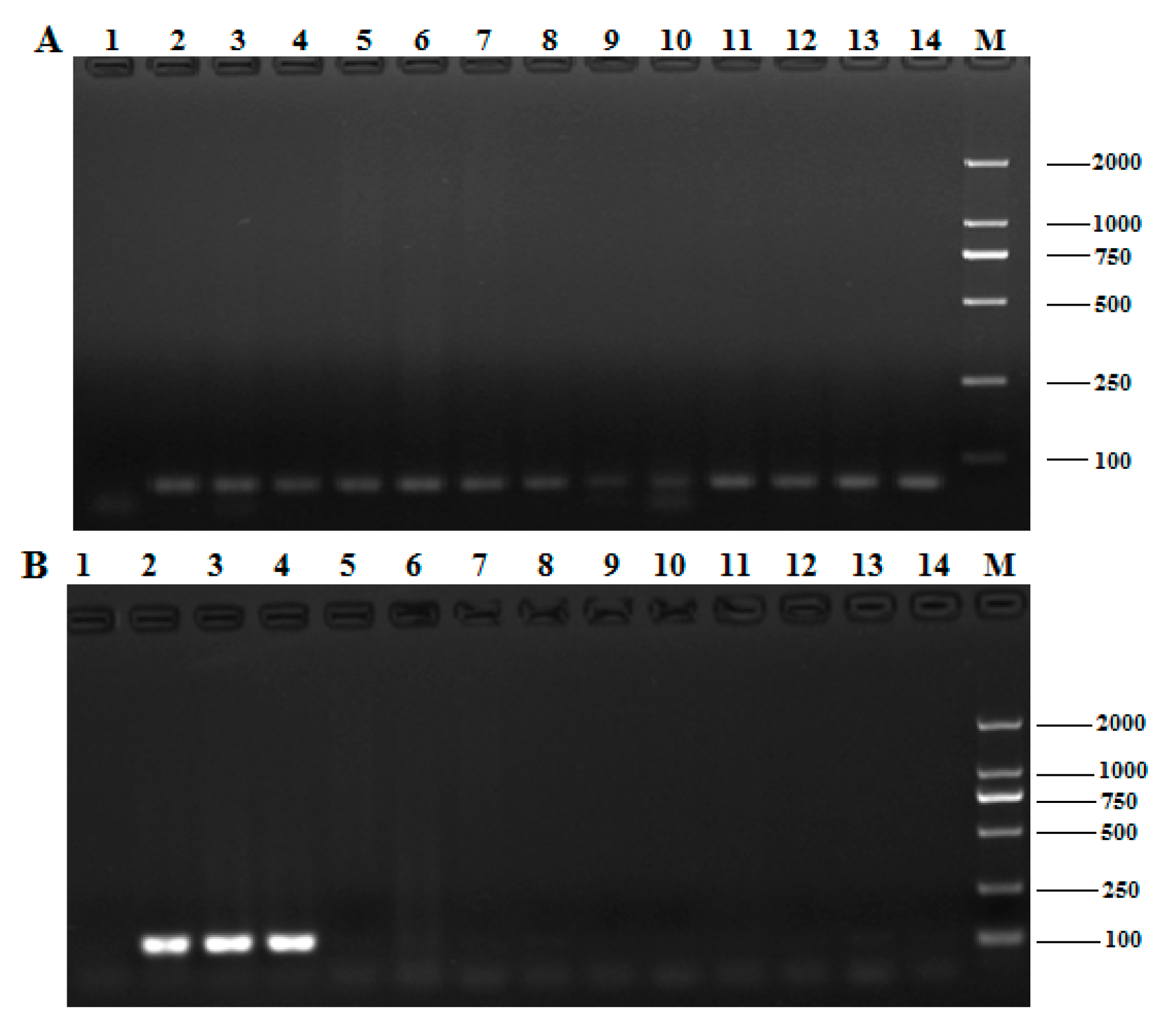
2.3. Species Specificity of Qualitative PCR aSSAYS
2.4. Homology Analysis of the Pol Gene among Different T. Matsutake Varieties
2.5. Confirmation of the Pol Gene Copy Number by Southern Blot
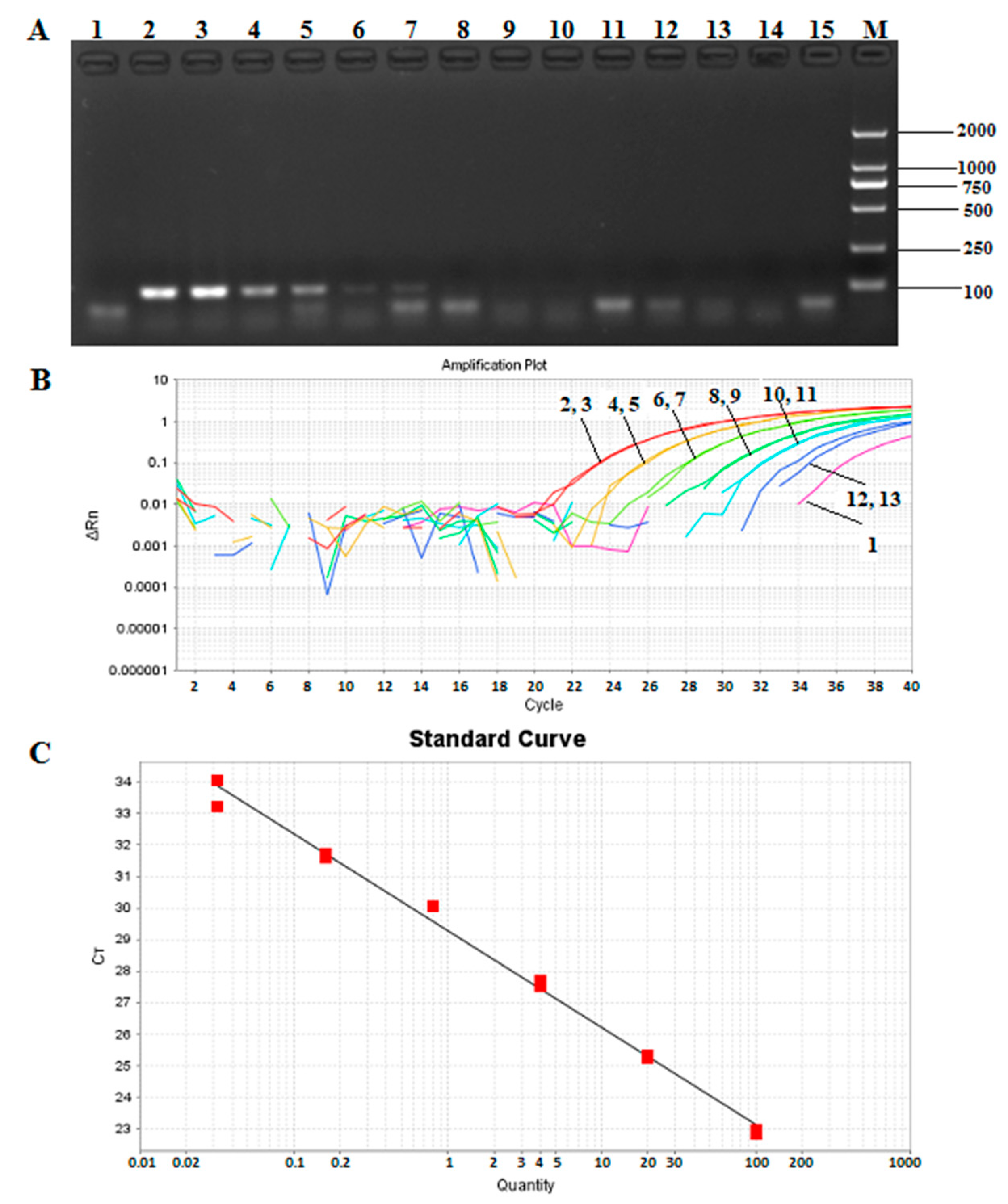
2.6. Sensitivity of the Qualitative and Taqman-Based Real-Time Quantitative PCR Assays
2.7. Application of the Pol gene to Detect Processed T. Matsutake Products
3. Materials and Methods
3.1. Materials
3.2. Genomic DNA Extraction
3.3. Enzyme Digestion of T. Matsutake Genomic DNA
3.4. Southern Blot
3.5. Species-Specific Diagnostic Marker Selection of T. Matsutake
3.6. Primers and Probe
3.7. PCR Conditions
3.8. Sensitivity of the Qualitative and Taqman-Based Real-Time Quantitative PCR Assays
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Xu, J.P.; Sha, T.; Li, Y.C.; Zhao, Z.W.; Yang, Z.L. Recombination and genetic differentiation among natural populations of the ectomycorrhizal mushroom Tricholoma matsutake from southwestern China. Mol. Ecol. 2008, 17, 1238–1247. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.; Tian, G.; Zhao, Y.; Zhao, L.; Wang, H.; Ng, B.T. A Fungal α-Galactosidase from Tricholoma matsutake with Broad Substrate Specificity and Good Hydrolytic Activity on Raffinose Family Oligosaccharides. Molecules 2015, 20, 13550–13562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, X.; You, Q.; Su, X. A comparison study on extraction of polysaccharides from Tricholoma matsutake by response surface methodology. Carbohydr. Polym. 2014, 102, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Vaario, L.-M.; Pennanen, T.; Lu, J.; Palmén, J.; Stenman, J.; Leveinen, J.; Kilpeläinen, P.; Kitunen, V. Tricholoma matsutake can absorb and accumulate trace elements directly from rock fragments in the shiro. Mycorrhiza 2015, 25, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Shi, Y. FT-TRS Analysis of Tricholoma matsutake Sing and Agaricus Blazei Murrill. Phys. Test. Chem. Anal. (Part B Chem. Anal.) 2013, 49, 1076–1078. [Google Scholar]
- Zhu, Y.; Zhang, H.; Zhu, Y.; Pu, X. An Appraisal of Past and Present Methods for Identifying Poisonous Mushrooms. J. Gansu Sci. 2008, 20, 44–48. [Google Scholar]
- Liu, Y.; Liu, N.; Zhang, Y. Research progress of food authentication technology. Sci. Technol. Food Ind. 2016, 37, 374–383. [Google Scholar]
- Chen, Y.; Dong, W.; Wu, Y.; Yuan, F.; Huang, W.; Ge, Y. Research and application of food authentication system. Sci. Technol. Food Ind. 2008, 29, 216–218. [Google Scholar]
- Qiang, L.; Li, Z.; Li, W.; Li, X.; Huang, W.; Hua, Y.; Zheng, L. Chemical compositions and volatile compounds of Tricholoma matsutake from different geographical areas at different stages of maturity. Food Sci. Biotechnol. 2016, 25, 71–77. [Google Scholar]
- Ding, X.; Hou, Y.L. Identification of genetic characterization and volatile compounds of Tricholoma matsutake from different geographical origins. Biochem. Syst. Ecol. 2012, 44, 233–239. [Google Scholar] [CrossRef]
- Yang, L.; Yang, Y.; Jin, W.; Zhang, X.; Li, X.; Wu, Y.; Li, J.; Li, L. Development and Interlaboratories Validation of Event-Specific Quantitative Real-Time PCR Method for Genetically Modified Rice G6H1 Event. J. Agric. Food Chem. 2018, 66, 8179. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, D.; Lu, X.; Wang, W.; Xu, Y.; He, Q. Phage Medicated Immuno-PCR for Ultrasensitive Detection of Cry1Ac protein based on Nanobody. J. Agric. Food Chem. 2016, 64, 7882–7889. [Google Scholar] [CrossRef]
- Xu, W.; Bai, W.; Guo, F.; Luo, Y.; Yuan, Y.; Huang, K. A papaya-specific gene, papain, used as an endogenous reference gene in qualitative and real-time quantitative PCR detection of transgenic papayas. Eur. Food Res. Technol. 2008, 228, 301–309. [Google Scholar] [CrossRef]
- Shang, Y.; Zhu, P.; Huang, K.; Liu, W.; Tian, W.; Luo, Y.; Xu, W. A peach (Prunus persica)-specific gene, Lhcb2, used as an endogenous reference gene for qualitative and real-time quantitative PCR to detect fruit products. LWT-Food Sci. Technol. 2014, 55, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.-M.; Shi, X.-M.; Pan, F. Species-specific diagnostic marker for rapid identification of Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 2007, 59, 379–382. [Google Scholar] [CrossRef]
- Liu, Y.K.; Li, H.P.; Huang, T.; Cheng, W.; Gao, C.S.; Zuo, D.Y.; Zhao, Z.X.; Liao, Y.C. Wheat-Specific Gene, Ribosomal Protein L21, Used as the Endogenous Reference Gene for Qualitative and Real-Time Quantitative Polymerase Chain Reaction Detection of Transgenes. J. Agric. Food Chem. 2014, 62, 10405–10413. [Google Scholar] [CrossRef]
- James, D.; Schmidt, A.-M.; Wall, E.; Green, M.; Masri, S. Reliable Detection and Identification of Genetically Modified Maize, Soybean, and Canola by Multiplex PCR Analysis. J. Agric. Food Chem. 2003, 51, 5829–5834. [Google Scholar] [CrossRef]
- Zeitler, R.; Pietsch, K.; Waiblinger, H.-U. Validation of real-time PCR methods for the quantification of transgenic contaminations in rape seed. Eur. Food Res. Technol. 2002, 214, 346–351. [Google Scholar] [CrossRef]
- Weng, H.; Yang, L.; Liu, Z.; Ding, J.; Pan, A.; Zhang, D. Novel Reference Gene, High-mobility-group protein I/Y, Used in Qualitative and Real-Time Quantitative Polymerase Chain Reaction Detection of Transgenic Rapeseed Cultivars. J. AOAC Int. 2005, 88, 577–584. [Google Scholar]
- Demeke, T.; Ratnayaka, I. Multiplex qualitative PCR assay for identification of genetically modified canola events and real-time event-specific PCR assay for quantification of the GT73 canola event. Food Control 2008, 19, 893–897. [Google Scholar] [CrossRef]
- Hernández, M.; Río, A.; Esteve, T.; Prat, S.; Pla, M. A Rapeseed-Specific Gene, Acetyl-CoA Carboxylase, Can Be Used as a Reference for Qualitative and Real-Time Quantitative PCR Detection of Transgenes from Mixed Food Samples. J. Agric. Food Chem. 2001, 49, 3622–3627. [Google Scholar] [CrossRef]
- Yoshimura, T.; Kuribara, H.; Matsuoka, T.; Kodama, T.; Iida, M.; Watanabe, T.; Akiyama, H.; Maitani, T.; Furui, S.; Hino, A. Applicability of the Quantification of Genetically Modified Organisms to Foods Processed from Maize and Soy. J. Agric. Food Chem. 2005, 53, 2052–2059. [Google Scholar] [CrossRef]
- Van Duijn, G.; Biert, R.V.; Bleeker-Marcelis, H.; Peppelman, H.; Hessing, M. Detection methods for genetically modified crops. Food Control 1999, 10, 375–378. [Google Scholar] [CrossRef]
- Chaouachi, M.; Giancola, S.; Romaniuk, M.; Laval, V.; Bertheau, Y.; Brunel, D. A Strategy for Designing Multi-Taxa Specific Reference Gene Systems. Example of Application––ppi Phosphofructokinase (ppi-PPF) Used for the Detection and Quantification of Three Taxa: Maize (Zea mays), Cotton (Gossypium hirsutum) and Rice (Oryza sativa). J. Agric. Food Chem. 2007, 55, 8003–8010. [Google Scholar] [CrossRef]
- Lee, S.-H.; Kim, J.-K.; Yi, B.-Y. Detection Methods for Biotech Cotton MON 15985 and MON 88913 by PCR. J. Agric. Food Chem. 2007, 55, 3351–3357. [Google Scholar] [CrossRef]
- Yang, L.; Chen, J.; Huang, C.; Liu, Y.; Jia, S.; Pan, L.; Zhang, D. Validation of a cotton-specific gene, Sad1, used as an endogenous reference gene in qualitative and real-time quantitative PCR detection of transgenic cottons. Plant Cell Rep. 2005, 24, 237–245. [Google Scholar] [CrossRef]
- Baeumler, S.; Wulff, D.; Tagliani, L.; Song, P. A Real-Time Quantitative PCR Detection Method Specific to Widestrike Transgenic Cotton (Event 281-24-236/3006-210-23). J. Agric. Food Chem. 2006, 54, 6527–6534. [Google Scholar] [CrossRef]
- Zimmermann, A.; Hemmer, W.; Liniger, M.; Lüthy, J.; Pauli, U. A Sensitive Detection Method For Genetically Modified MaisGardTMCorn Using a Nested PCR-system. LWT-Food Sci. Technol. 1998, 31, 664–667. [Google Scholar] [CrossRef]
- Pardigol, A.; Guillet, S.; Pöpping, B. A simple procedure for quantification of genetically modified organisms using hybrid amplicon standards. Eur. Food Res. Technol. 2003, 216, 412–420. [Google Scholar] [CrossRef]
- Hernández, M.; Duplan, M.-N.; Berthier, G.; Vaïtilingom, M.; Hauser, W.; Freyer, R.; Pla, M.; Bertheau, Y. Development and Comparison of Four Real-Time Polymerase Chain Reaction Systems for Specific Detection and Quantification of Zea mays L. J. Agric. Food Chem. 2004, 52, 4632–4637. [Google Scholar] [CrossRef]
- Vaïtilingom, M.; Pijnenburg, H.; Gendre, F.; Brignon, P. Real-Time Quantitative PCR Detection of Genetically Modified Maximizer Maize and Roundup Ready Soybean in Some Representative Foods. J. Agric. Food Chem. 1999, 47, 5261–5266. [Google Scholar] [CrossRef] [PubMed]
- Höhne, M.; Santisi, C.; Meyer, R. Real-time multiplex PCR: An accurate method for the detection and quantification of 35S-CaMV promoter in genetically modified maize-containing food. Eur. Food Res. Technol. 2002, 215, 59–64. [Google Scholar] [CrossRef]
- Matsuoka, T.; Kuribara, H.; Takubo, K.; Akiyama, H.; Miura, H.; Goda, Y.; Kusakabe, Y.; Isshiki, K.; Toyoda, M.; Hino, A. Detection of Recombinant DNA Segments Introduced to Genetically Modified Maize (Zea mays). J. Agric. Food Chem. 2002, 50, 2100–2109. [Google Scholar] [CrossRef] [PubMed]
- Brodmann, P.D.; Ilg, E.C.; Berthoud, H.; Herrmann, A. Real-Time Quantitative Polymerase Chain Reaction Methods for Four Genetically Modified Maize Varieties and Maize DNA Content in Food. J. AOAC Int. 2002, 85, 646–653. [Google Scholar] [PubMed]
- Rønning, S.B.; Vaïtilingom, M.; Berdal, K.G.; Holst-Jensen, A. Event specific real-time quantitative PCR for genetically modified Bt11 maize (Zea mays). Eur. Food Res. Technol. 2003, 216, 347–354. [Google Scholar] [CrossRef]
- Kuribara, H.; Shindo, Y.; Matsuoka, T.; Takubo, K.; Futo, S.; Aoki, N.; Hirao, T.; Akiyama, H.; Goda, Y.; Toyada, M.; et al. Novel Reference Molecules for Quantitation of Genetically Modified Maize and Soybean. J. AOAC Int. 2002, 85, 1077–1089. [Google Scholar]
- Lee, S.-H.; Min, D.-M.; Kim, J.-K. Qualitative and Quantitative Polymerase Chain Reaction Analysis for Genetically Modified Maize MON863. J. Agric. Food Chem. 2006, 54, 1124–1129. [Google Scholar] [CrossRef]
- Hernández, M.; Esteve, T.; Pla, M. Real-Time Polymerase Chain Reaction Based Assays for Quantitative Detection of Barley, Rice, Sunflower, and Wheat. J. Agric. Food Chem. 2005, 53, 7003–7009. [Google Scholar] [CrossRef]
- Jeong, S.-C.; Pack, I.S.; Cho, E.-Y.; Youk, E.S.; Park, S.; Yoon, W.K.; Kim, C.-G.; Choi, Y.D.; Kim, J.-K.; Kim, H.M. Molecular analysis and quantitative detection of a transgenic rice line expressing a bifunctional fusion TPSP. Food Control 2007, 18, 1434–1442. [Google Scholar] [CrossRef]
- Ding, J.; Jia, J.; Yang, L.; Wen, H.; Zhang, C.; Liu, W.; Zhang, D. Validation of a Rice Specific Gene, Sucrose Phosphate Synthase, Used as the Endogenous Reference Gene for Qualitative and Real-Time Quantitative PCR Detection of Transgenes. J. Agric. Food Chem. 2004, 52, 3372–3377. [Google Scholar] [CrossRef]
- Liu, Q.; Qi, X.; Yan, H.; Huang, L.; Nie, G.; Zhang, X. Reference Gene Selection for Quantitative Real-Time Reverse-Transcriptase PCR in Annual Ryegrass (Lolium multiflorum) Subjected to Various Abiotic Stresses. Molecules 2018, 23, 172. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Shao, B.; Chen, W.; Li, S.; Wang, Z.; Zhun, X.; Liao, J. Study on the Comparison of three Genomic DNA Extraction Methods for 15 Familiar Edulis and Medicincal Fungi. Food Sci. 2004, 25, 36–40. [Google Scholar]
- González-Mendoza, D.; Argumedo-Delira, R.; Morales-Trejo, A.; Pulido-Herrera, A.; Cervantes-Díaz, L.; Grimaldo-Juarez, O.; Alarcón, A. A rapid method for isolation of total DNA from pathogenic filamentous plant fungi. Genet. Mol. Res. 2010, 9, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Murata, H.; Yamada, A.; Babasaki, K. Identification of Repetitive Sequences Containing Motifs of Retrotransposons in the Ectomycorrhizal Basidiomycete Tricholoma matsutake. Mycologia 1999, 91, 766–775. [Google Scholar] [CrossRef]
- Murata, H.; Yamada, A. marY1, a Member of the gypsy Group of Long Terminal Repeat Retroelements from the Ectomycorrhizal Basidiomycete Tricholoma matsutake. Appl. Environ. Microbiol. 2000, 66, 3642–3645. [Google Scholar] [CrossRef] [Green Version]
- Nan, Z.; Xu, W.; Bai, W.; Zhai, Z.; Luo, Y.; Yan, X.; Jing, H.; Huang, K. Event-specific qualitative and quantitative PCR detection of LY038 maize in mixed samples. Food Control 2011, 22, 1287–1295. [Google Scholar]
- Shang, Y.; Yan, Y.; Xu, W.; Tian, W.; Huang, K. Research on Gene Mobility and Gene Flow Between Genetically Modified Mon 15985 Cotton and Pleurotus Ostreatus. J. Food Saf. 2016, 36, 423–432. [Google Scholar] [CrossRef]
- Yan, Y.; Xu, W.; Su, C.; Luo, Y.; Wang, Y.; Gu, X.; Dai, Y.; Tian, H. Study on the methods of extraction of Pleurotus ostreatus genomic DNA. Sci. Technol. Food Ind. 2011, 190–193. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | OD260/280 | Concentration (ng/μL) |
|---|---|---|
| CTAB | 2.01 ± 0.02 a | 983.33 ± 2.89 a |
| SDS-CTAB | 1.97 ± 0.01 a | 1138.33 ± 2.89 b |
| High-concentration salt precipitation | 1.88 ± 0.02 b | 676.67 ± 0.01 c |
| Kit method | 1.66 ± 0.11 c | 163.33 ± 2.89 d |
| Primer Name | Primer Sequence (5′→3′) | Length | Product Size (bp) | Reference |
|---|---|---|---|---|
| 18S-F | CCTGAGAAACGGCTACCAT | 19 | 80 | This study |
| 18S-R | ATCTTCACTACCTCCCCATTCTG | 23 | ||
| pol-F | GACTCCCATACTGAAGCCAAT | 21 | 107 (from 69 to 175 bp) | |
| pol-R | ACTCCTTTCCATGCCCATAC | 20 | ||
| pol-probe | (FAM)-TGGCTCCTACTCCAAACACTGACAC-(TAMRA) | |||
| Southern blot-F | CGTGATGGATGGAATACCTGT | 21 | 775 (from 563 to 1157 bp) | |
| Southern blot-R | GTGTACCCCCCCTTAGACTGA | 21 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shan, L.; Wang, D.; Li, Y.; Zheng, S.; Xu, W.; Shang, Y. Identification of the Pol Gene as a Species-Specific Diagnostic Marker for Qualitative and Quantitative PCR Detection of Tricholoma matsutake. Molecules 2019, 24, 455. https://doi.org/10.3390/molecules24030455
Shan L, Wang D, Li Y, Zheng S, Xu W, Shang Y. Identification of the Pol Gene as a Species-Specific Diagnostic Marker for Qualitative and Quantitative PCR Detection of Tricholoma matsutake. Molecules. 2019; 24(3):455. https://doi.org/10.3390/molecules24030455
Chicago/Turabian StyleShan, Luying, Dazhou Wang, Yinjiao Li, Shi Zheng, Wentao Xu, and Ying Shang. 2019. "Identification of the Pol Gene as a Species-Specific Diagnostic Marker for Qualitative and Quantitative PCR Detection of Tricholoma matsutake" Molecules 24, no. 3: 455. https://doi.org/10.3390/molecules24030455
APA StyleShan, L., Wang, D., Li, Y., Zheng, S., Xu, W., & Shang, Y. (2019). Identification of the Pol Gene as a Species-Specific Diagnostic Marker for Qualitative and Quantitative PCR Detection of Tricholoma matsutake. Molecules, 24(3), 455. https://doi.org/10.3390/molecules24030455