Synergistic Effects of The Enhancements to Mitochondrial ROS, p53 Activation and Apoptosis Generated by Aspartame and Potassium Sorbate in HepG2 Cells

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. The Effect of AT and PS Inhibited the Proliferation in HepG2 Cells
Apoptosis induced by AT together with PS in HepG2 cells
2.2. AT Together with PS Induced the Changes of MMP, CMP and Protein Content of Cytochrome c in HepG2 Cells
2.3. AT Together with PS Induced the Release of ROS in HepG2 Cells
2.4. AT Together with PS Induced DNA Damage in HepG2 Cells
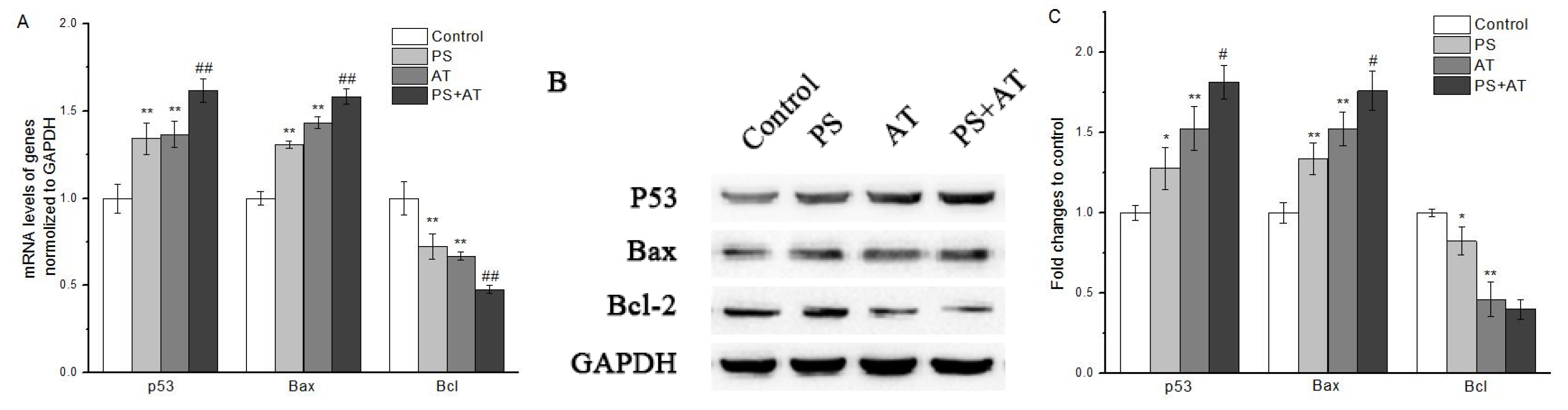
2.5. AT Together with PS Induced Apoptosis through Activation of p53, Bax and Bcl-2 in HepG2 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. Cell Viability Assay
4.3. Evaluation of Joint Effect
4.4. Cell Apoptosis Detection
4.5. Mitochondrial Health Detection
4.6. Measurement of Intracellular ROS Release
4.7. DNA Damage Detection
4.8. RNA Extraction and Quantitative Real-Time RT-PCR
4.9. Western Blot Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kaiser, K.L.E.; Palabrica, V.S. Photobacterium phosphoreum Toxicity Data Index. Water Qual. Res. J. 1991, 26, 361–431. [Google Scholar]
- Gören, A.C.; Bilsel, G.; Şimşek, A.; Bilsel, M.; Akçadağ, F.; Topal, K.; Ozgen, H. HPLC and LC-MS/MS methods for determination of sodium benzoate and potassium sorbate in food and beverages: Performances of local accredited laboratories via proficiency tests in Turkey. Food Chem. 2015, 175, 273–279. [Google Scholar] [CrossRef]
- Carocho, M.; Barreiro, M.F.; Morales, P.; Ferreira, I.C.F.R. Adding Molecules to Food, Pros and Cons: A Review on Synthetic and Natural Food Additives. Compr. Rev. Food Sci. Food Safety 2014, 13, 377–399. [Google Scholar] [CrossRef] [Green Version]
- Oyama, Y.; Sakai, H.; Arata, T.; Okano, Y.; Akaike, N.; Sakai, K.; Noda, K. Cytotoxic effects of methanol, formaldehyde, and formate on dissociated rat thymocytes: A possibility of aspartame toxicity. Cell Biol. Toxicol. 2002, 18, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Renwick, A.G. The intake of intense sweeteners - an update review. Food Addit. Contam. 2006, 23, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Zanella, F.; Lorens, J.B.; Link, W. High content screening: Seeing is believing. Trends Biotechnol. 2010, 28, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Persson, M.; Mow, T.; Hornberg, J.J. A high content screening assay to predict human drug-induced liver injury during drug discovery. J. Pharmacol. Toxicol. Methods 2013, 68, 302–313. [Google Scholar] [CrossRef]
- Tolosa, L.; Gómezlechón, M.J.; Donato, M.T. High-content screening technology for studying drug-induced hepatotoxicity in cell models. Arch. Toxicol. 2015, 89, 1007–1022. [Google Scholar] [CrossRef]
- Westerink, W.M.A.; Schirris, T.J.J.; Horbach, G.J.; Schoonen, W.G.E.J. Development and validation of a high-content screening in vitro micronucleus assay in CHO-k1 and HepG2 cells. Mutation Res. Genet. Toxicol. Environ. Mutagenesis 2011, 724, 7–21. [Google Scholar] [CrossRef]
- Tolosa, L.; Gómezlechón, M.J.; Pérezcataldo, G.; Castell, J.V.; Donato, M.T. HepG2 cells simultaneously expressing five P450 enzymes for the screening of hepatotoxicity: Identification of bioactivable drugs and the potential mechanism of toxicity involved. Arch. Toxicol. 2013, 87, 1115–1127. [Google Scholar] [CrossRef]
- Ma, J.; Feng, Y.; Yang, L.; Li, X. PUMA and survivin are involved in the apoptosis of HepG2 cells induced by microcystin-LR via mitochondria-mediated pathway. Chemosphere 2016, 157, 241–249. [Google Scholar] [CrossRef]
- Olson, H.; Betton, G.; Robinson, D.; Thomas, K.; Monro, A.; Kolaja, G.; Lilly, P.; Sanders, J.; Sipes, G.; Bracken, W. Concordance of the toxicity of pharmaceuticals in humans and in animals. Regul. Toxicol. Pharm. 2000, 32, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, Y.F.; Kawaguchi, S.; Kamaya, A.; Ohshita, M.; Kabasawa, K.; Iwama, K.; Taniguchi, K.; Tsuda, S. The comet assay with 8 mouse organs: Results with 39 currently used food additives. Mutation Res. Genet. Toxicol. Environ. Mutagenesis 2002, 519, 103–119. [Google Scholar] [CrossRef]
- Lau, K.; Mclean, W.G.; Williams, D.P.; Howard, C.V. Synergistic interactions between commonly used food additives in a developmental neurotoxicity test. Toxicol. Sci. 2006, 90, 178. [Google Scholar] [CrossRef] [PubMed]
- Münzner, R.; Guigas, C.; Renner, H.W. Re-examination of potassium sorbate and sodium sorbate for possible genotoxic potential. Food Chem. Toxicol. 1990, 28, 397–401. [Google Scholar] [CrossRef]
- Mamur, S.; Yüzbaşioğlu, D.; Unal, F.; Yilmaz, S. Does potassium sorbate induce genotoxic or mutagenic effects in lymphocytes? Toxicol. in Vitro 2010, 24, 790–794. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Giri, A.K.; Talukder, G.; Sharma, A. Sister chromatid exchanges and micronuclei formations induced by sorbic acid and sorbic acid-nitrite in vivo in mice. Toxicol. Lett. 1988, 42, 47–53. [Google Scholar] [CrossRef]
- Pandurangan, M.; Enkhtaivan, G.; Mistry, B.; Chandrasekaran, M.; Noorzai, R.; Kim, D.H. Investigation of role of aspartame on apoptosis process in HeLa cells. Saudi J. Biol. Sci. 2016, 23, 503–506. [Google Scholar] [CrossRef]
- Sarıkaya, R.; Cakır, S. Genotoxicity testing of four food preservatives and their combinations in the Drosophila wing spot test. Environ. Toxicol. Pharmacol. 2005, 20, 424–430. [Google Scholar] [CrossRef]
- Türkoğlu, S. Evaluation of genotoxic effects of sodium propionate, calcium propionate and potassium propionate on the root meristem cells of Allium cepa. Food Chem. Toxicol. 2008, 46, 2035–2041. [Google Scholar] [CrossRef]
- Ashkenazi, A. Targeting the extrinsic apoptosis pathway in cancer. Cytokine Growth Factor Rev. 2008, 19, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.B.; Liu, Y.Q.; Cui, Y.F. Pathways to caspase activation. Cell Biol. Int. 2013, 29, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Ly, J.D.; Grubb, D.R.; Lawen, A. The mitochondrial membrane potential (Δψm) in apoptosis; an update. Apoptosis 2003, 8, 115. [Google Scholar] [CrossRef] [PubMed]
- Paradies, G.; Petrosillo, G.; Pistolese, M.; Ruggiero, F.M. Reactive oxygen species affect mitochondrial electron transport complex I activity through oxidative cardiolipin damage. Gene 2002, 286, 135–141. [Google Scholar] [CrossRef]
- Wang, C.; Li, T.; Tang, S.; Zhao, D.; Zhang, C.; Zhang, S.; Deng, S.; Zhou, Y.; Xiao, X. Thapsigargin induces apoptosis when autophagy is inhibited in HepG2 cells and both processes are regulated by ROS-dependent pathway. Environ. Toxicol. Pharmacol. 2016, 41, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, J. Initiator caspases in apoptosis signaling pathways. Apoptosis 2002, 7, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Chipuk, J.E.; Moldoveanu, T.; Llambi, F.; Parsons, M.J.; Green, D.R. The BCL-2 Family Reunion. Molecular Cell 2010, 37, 299–310. [Google Scholar] [CrossRef]
- Rudner, J.; Elsaesser, S.J.; Müller, A.C.; Belka, C.; Jendrossek, V. Differential effects of anti-apoptotic Bcl-2 family members Mcl-1, Bcl-2, and Bcl-xL on celecoxib-induced apoptosis. Biochem. Pharmacol. 2010, 79, 10–20. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, J.; Liu, Y.; Gong, L.; Sun, B. Wheat bran feruloyl oligosaccharides ameliorate AAPH-induced oxidative stress in HepG2 cells via Nrf2 signalling. J. Funct. Foods 2016, 25, 333–340. [Google Scholar] [CrossRef]
- Zhang, B.; Peng, X.; Li, G.; Xu, Y.; Xia, X.; Wang, Q. Oxidative stress is involved in Patulin induced apoptosis in HEK293 cells. Toxicon 2015, 94, 1–7. [Google Scholar] [CrossRef]
- Chen, W.; Hou, J.; Yin, Y.; Jang, J.; Zheng, Z.; Fan, H.; Zou, G. Alpha-Bisabolol induces dose- and time-dependent apoptosis in HepG2 cells via a Fas- and mitochondrial-related pathway, involves p53 and NFkappaB. Biochem. Pharmacol. 2010, 80, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Kwak, G.H.; Kim, H.Y. MsrB3 deficiency induces cancer cell apoptosis through p53-independent and ER stress-dependent pathways. Arch. Biochem. Biophys. 2017, 621, 1. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhou, X.; Wang, J.; Zhang, W.; Zheng, H.; Lu, W.; Yuan, J. MEHP-induced oxidative DNA damage and apoptosis in HepG2 cells correlates with p53-mediated mitochondria-dependent signaling pathway. Food Chem. Toxicol. 2012, 50, 2424–2431. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qu, D.; Jiang, M.; Huang, D.; Zhang, H.; Feng, L.; Chen, Y.; Zhu, X.; Wang, S.; Han, J. Synergistic Effects of The Enhancements to Mitochondrial ROS, p53 Activation and Apoptosis Generated by Aspartame and Potassium Sorbate in HepG2 Cells. Molecules 2019, 24, 457. https://doi.org/10.3390/molecules24030457
Qu D, Jiang M, Huang D, Zhang H, Feng L, Chen Y, Zhu X, Wang S, Han J. Synergistic Effects of The Enhancements to Mitochondrial ROS, p53 Activation and Apoptosis Generated by Aspartame and Potassium Sorbate in HepG2 Cells. Molecules. 2019; 24(3):457. https://doi.org/10.3390/molecules24030457
Chicago/Turabian StyleQu, Daofeng, Mengxue Jiang, Dongping Huang, Hui Zhang, Lifang Feng, Yuewen Chen, Xuan Zhu, Suhua Wang, and Jianzhong Han. 2019. "Synergistic Effects of The Enhancements to Mitochondrial ROS, p53 Activation and Apoptosis Generated by Aspartame and Potassium Sorbate in HepG2 Cells" Molecules 24, no. 3: 457. https://doi.org/10.3390/molecules24030457
APA StyleQu, D., Jiang, M., Huang, D., Zhang, H., Feng, L., Chen, Y., Zhu, X., Wang, S., & Han, J. (2019). Synergistic Effects of The Enhancements to Mitochondrial ROS, p53 Activation and Apoptosis Generated by Aspartame and Potassium Sorbate in HepG2 Cells. Molecules, 24(3), 457. https://doi.org/10.3390/molecules24030457