
4-Cyanoindole-2′-deoxyribonucleoside as a Dual Fluorescence and Infrared Probe of DNA Structure and Dynamics

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
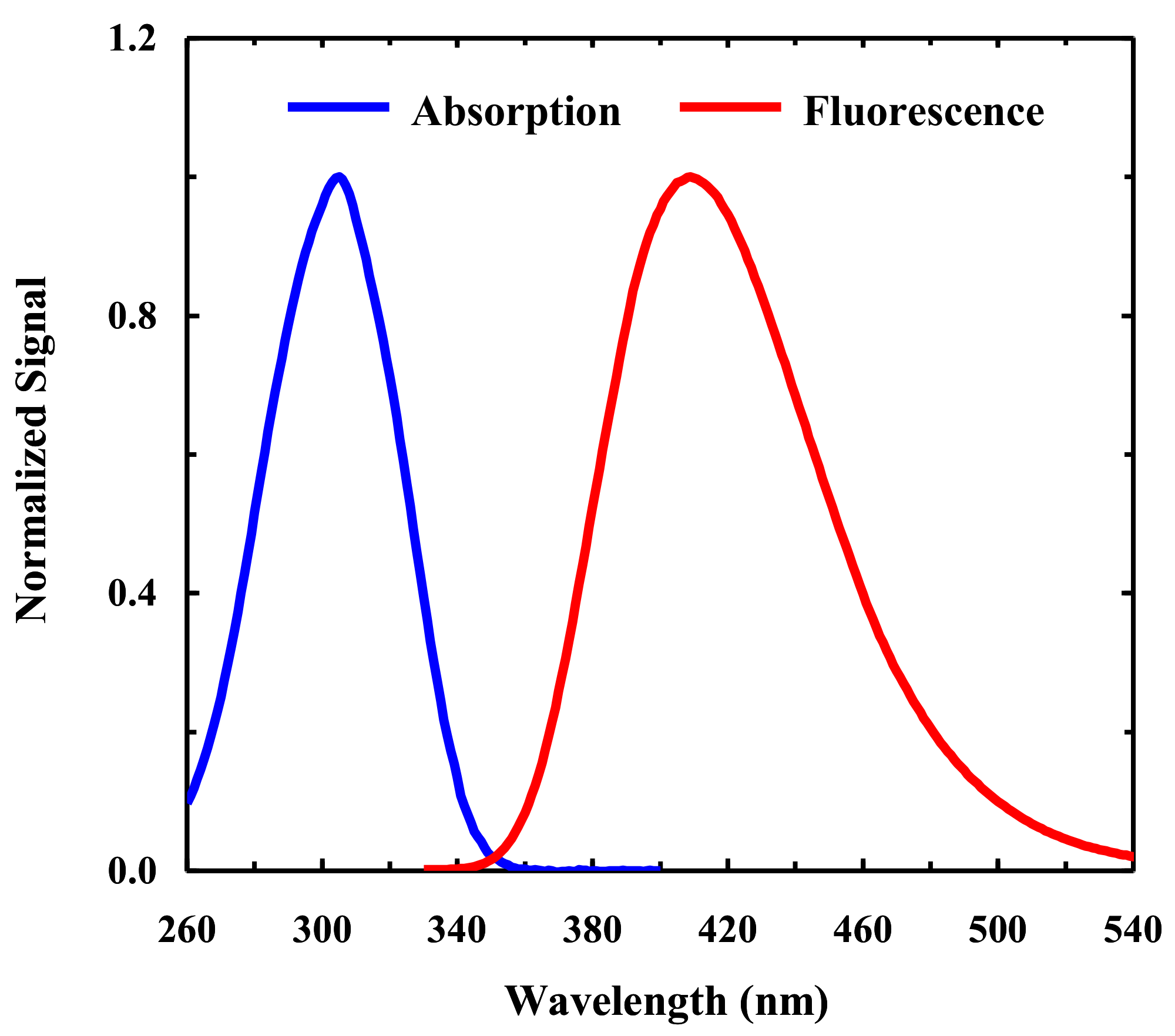
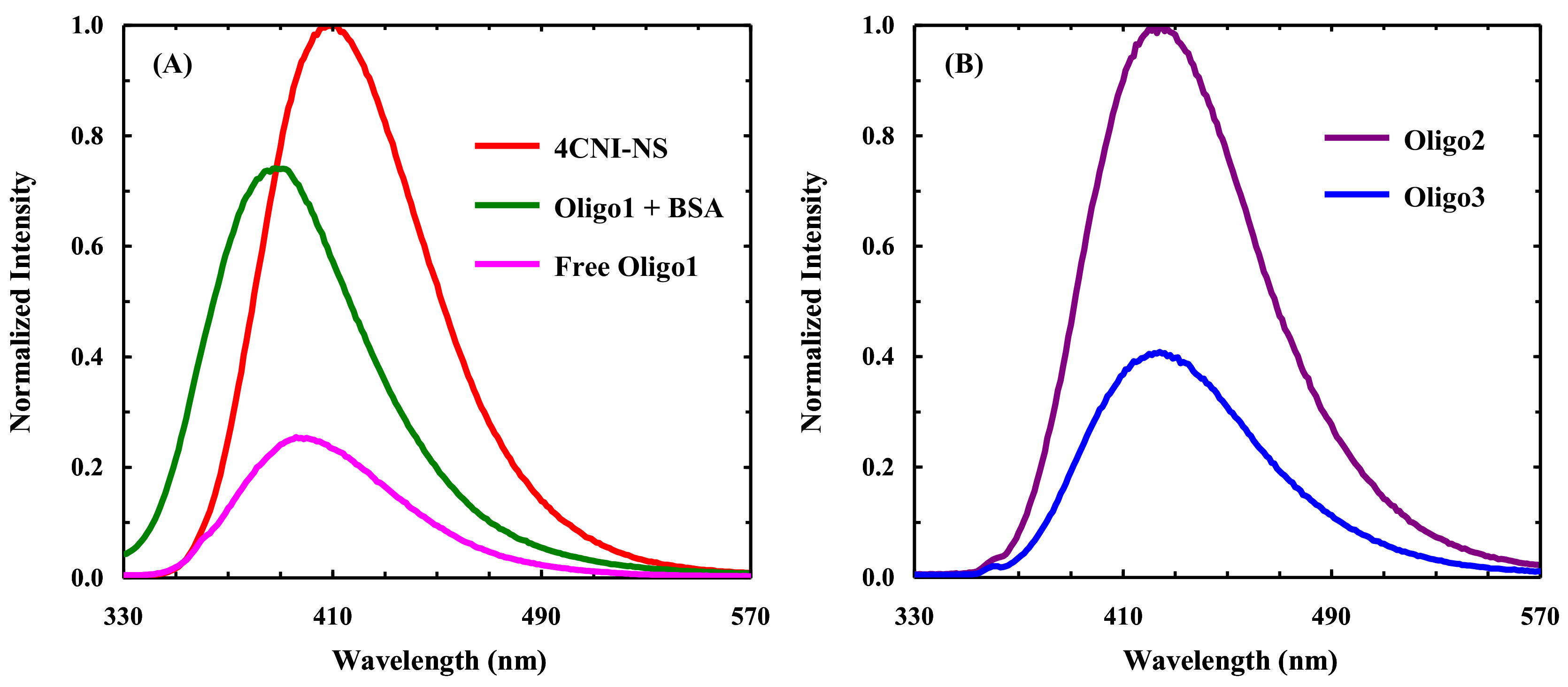
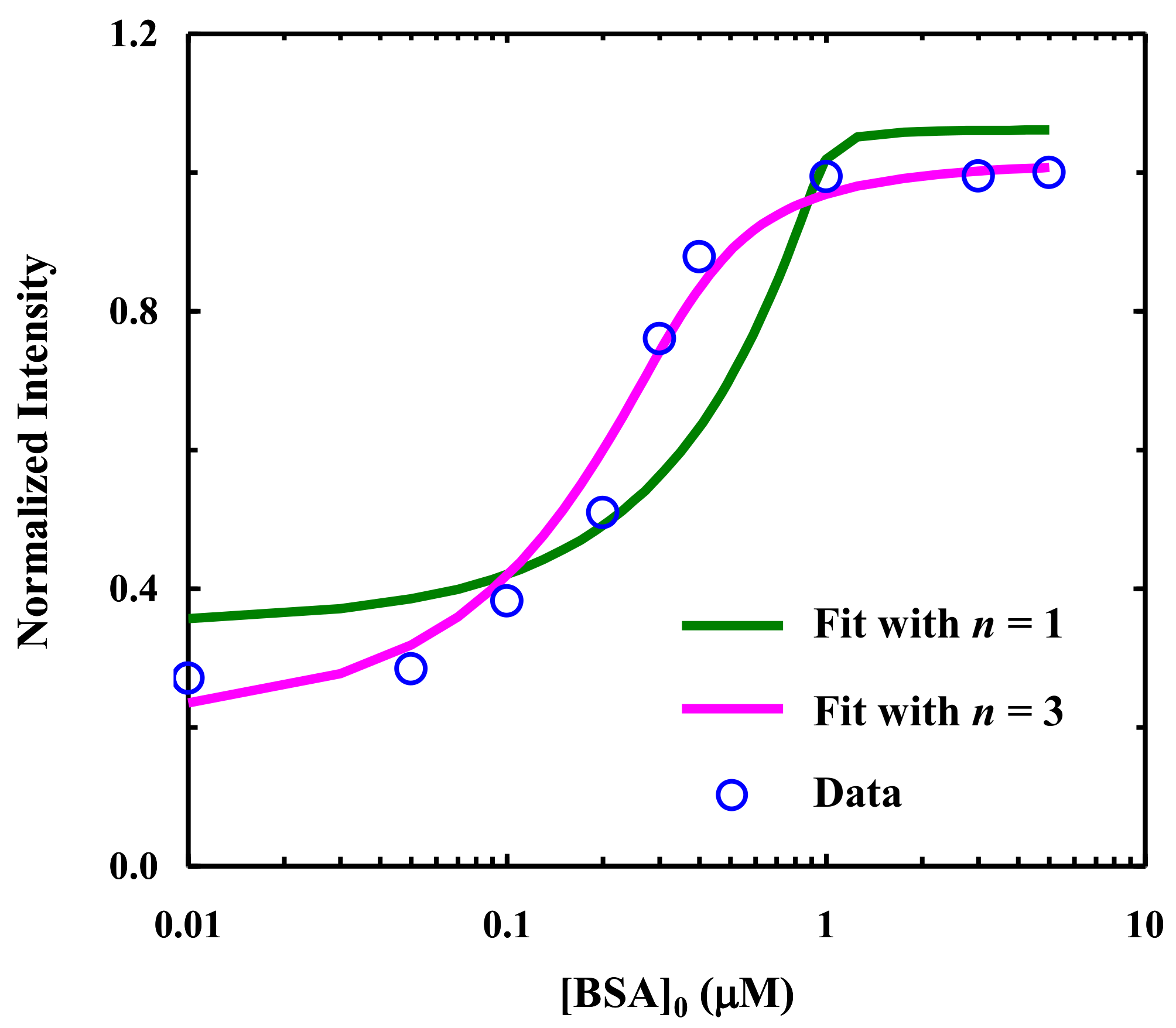
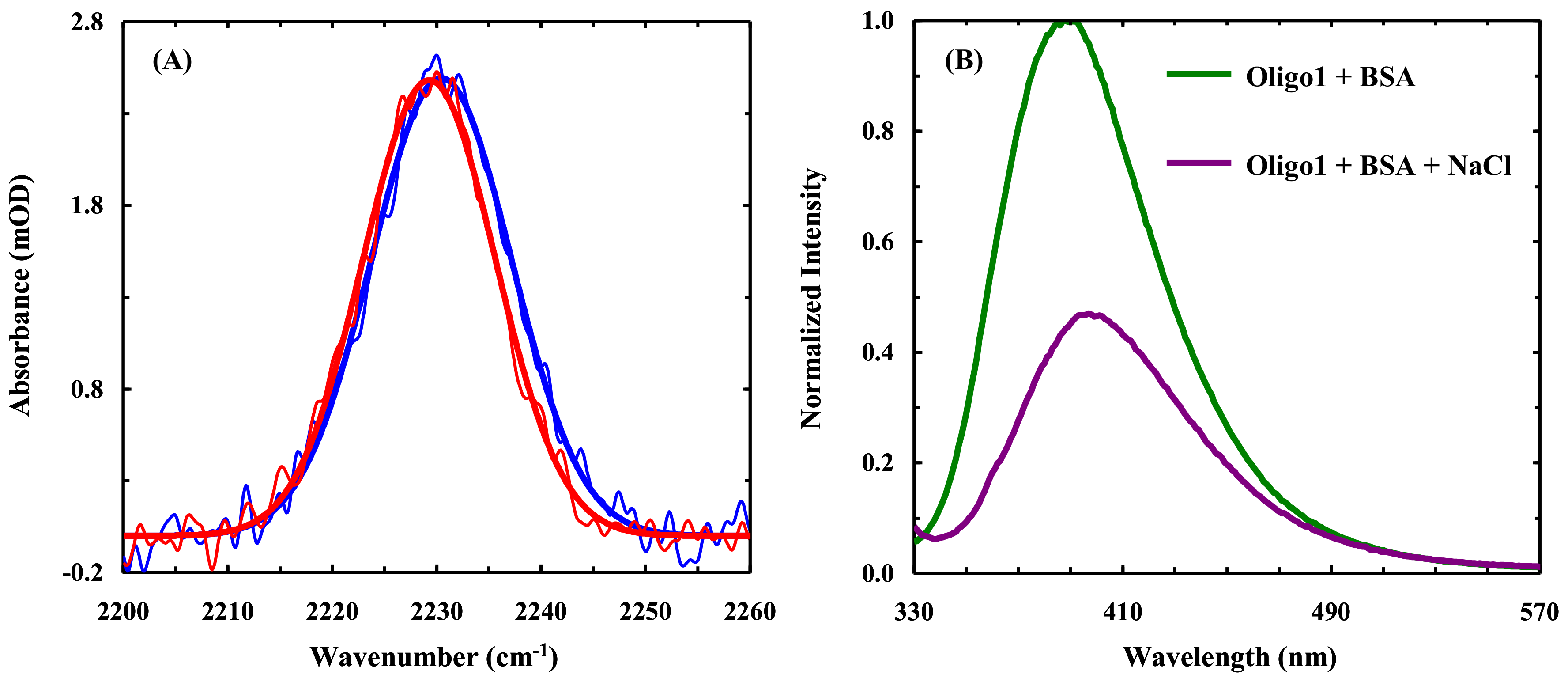
2.1. Fluorescence Study
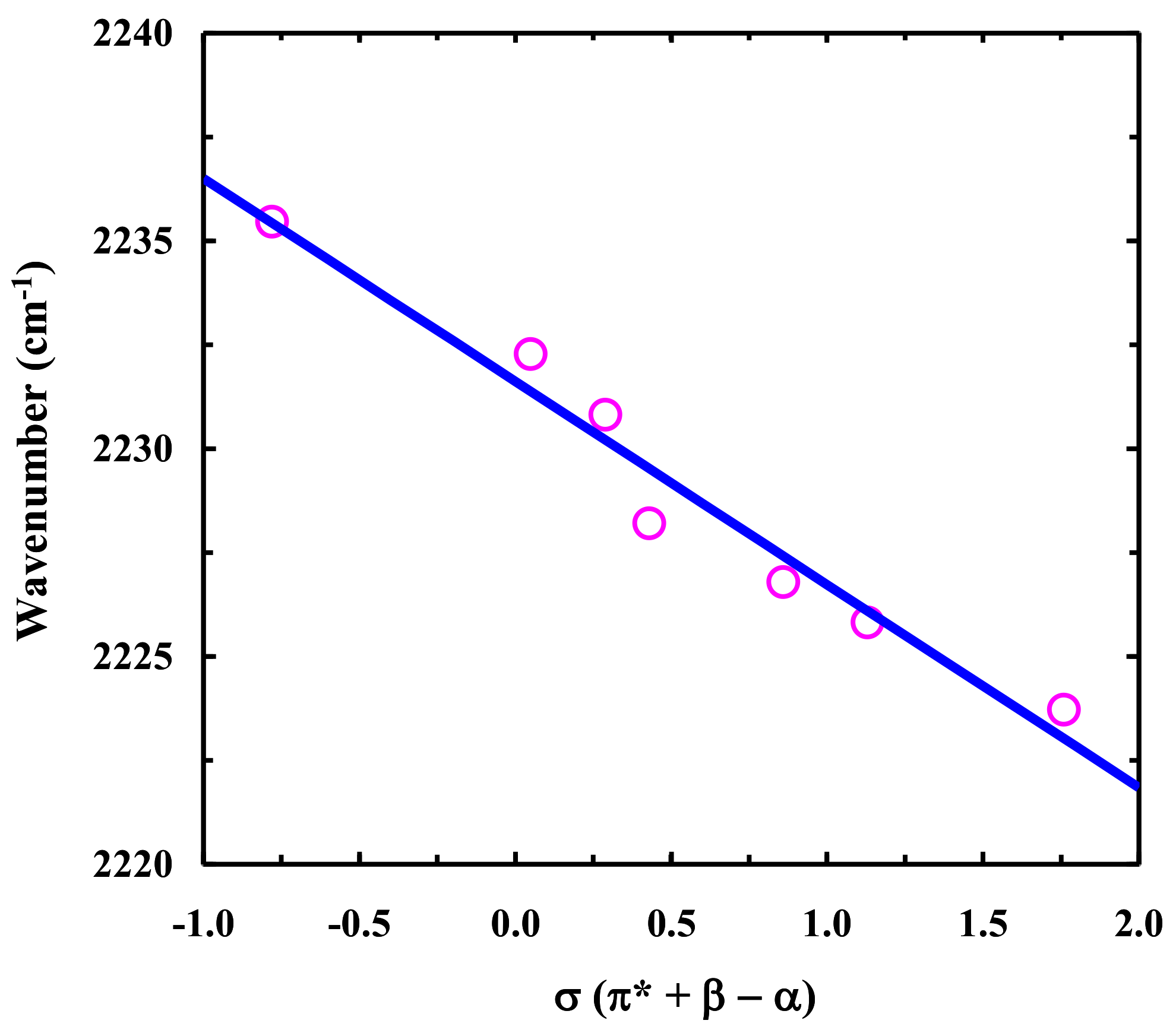
2.2. FTIR Study
3. Materials and Methods
3.1. Sample Preparation
3.2. Fluorescence Measurement
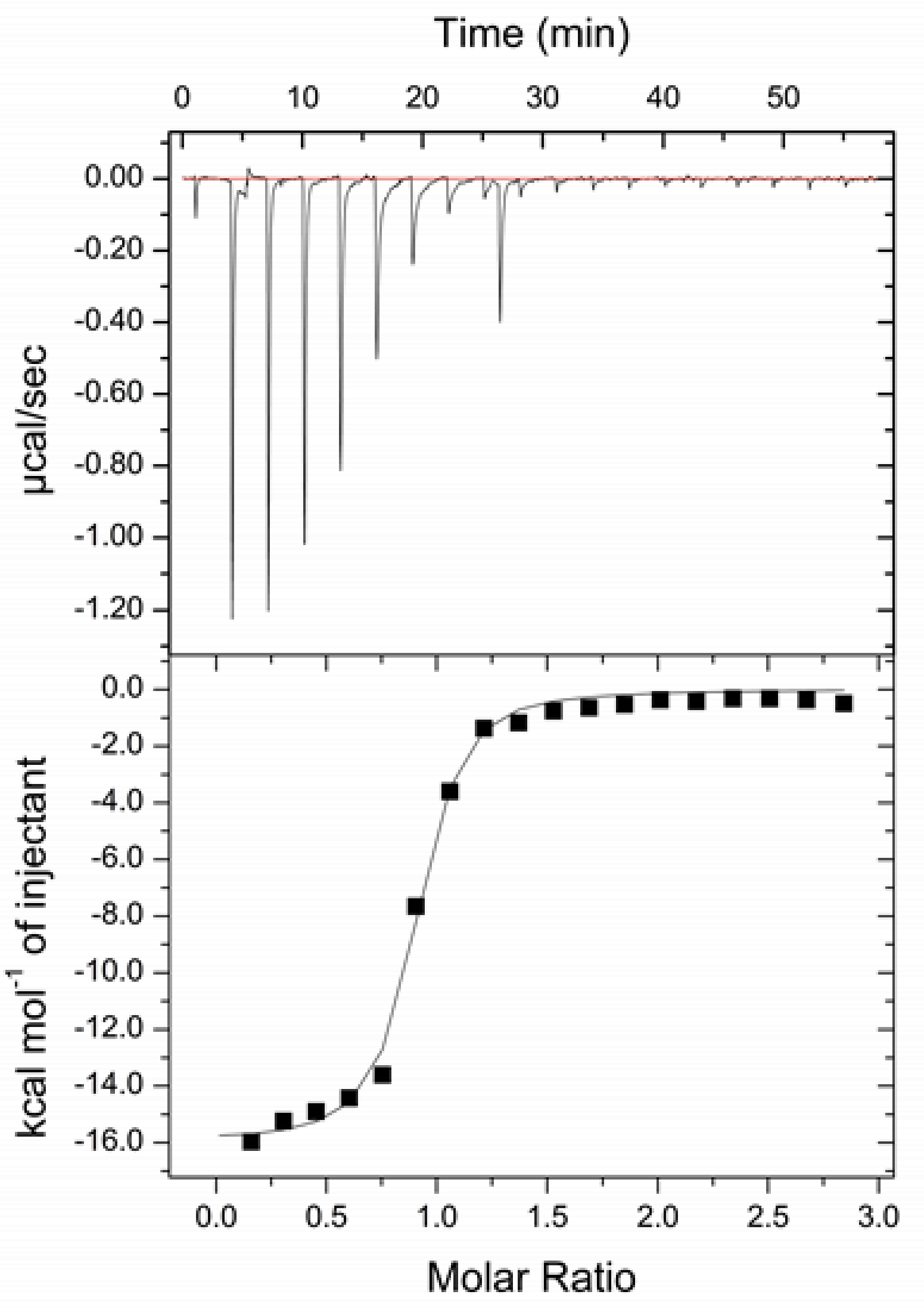
3.4. Isothermal Titration Calorimetry Measurement
3.5. FTIR Measurement
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Serrano, A.L.; Waegele, M.M.; Gai, F. Spectroscopic studies of protein folding: Linear and nonlinear methods. Protein Sci. 2012, 21, 157–170. [Google Scholar] [CrossRef]
- Ma, J.; Pazos, I.M.; Zhang, W.; Culik, R.M.; Gai, F. Site-specific infrared probes of proteins. Annu. Rev. Phys. Chem. 2015, 66, 357–377. [Google Scholar] [PubMed]
- Haney, C.M.; Wissner, R.F.; Petersson, E.J. Multiply labeling proteins for studies of folding and stability. Curr. Opin. Chem. Biol. 2015, 28, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Hanoian, P.; Liu, C.T.; Hammes-Schiffer, S.; Benkovic, S. Perspectives on electrostatics and conformational motions in enzyme catalysis. Acc. Chem. Res. 2015, 48, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Boxer, S.G. Stark realities. J. Phys. Chem. B 2009, 113, 2972–2983. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Ostrander, J.S.; Zanni, M.T. Watching proteins wiggle: Mapping structures with two-dimensional infrared spectroscopy. Chem. Rev. 2017, 117, 10726–10759. [Google Scholar] [CrossRef]
- Błasiak, B.; Londergan, C.H.; Webb, L.J.; Cho, M. Vibrational probes: From small molecule solvatochromism theory and experiments to applications in complex systems. Acc. Chem. Res. 2017, 50, 968–976. [Google Scholar] [CrossRef]
- Adhikary, R.; Zimmermann, J.; Romesberg, F.E. Transparent window vibrational probes for the characterization of proteins with high structural and temporal resolution. Chem. Rev. 2017, 117, 1927–1969. [Google Scholar] [CrossRef]
- Loakes, D.; Brown, D.M. 5-Nitroindole as an universal base analogue. Nucleic Acids Res. 1994, 22, 4039–4043. [Google Scholar] [CrossRef]
- Passow, K.T.; Harki, D.A. 4–Cyanoindole-2′-deoxyribonucleoside (4CIN): A universal fluorescent nucleoside analogue. Org. Lett. 2018, 20, 4310–4313. [Google Scholar] [CrossRef]
- Hilaire, M.R.; Mukherjee, D.; Troxler, T.; Gai, F. Solvent dependence of cyanoindole fluorescence lifetime. Chem. Phys. Lett. 2017, 685, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Hilaire, M.R.; Ahmed, I.A.; Lin, C.-W.; Jo, H.; DeGrado, W.F.; Gai, F. Blue fluorescent amino acid for biological spectroscopy and microscopy. Proc. Natl. Acad. Sci. USA 2017, 114, 6005–6009. [Google Scholar] [CrossRef] [PubMed]
- Van Wilderen, L.J.G.W.; Brunst, H.; Gustmann, H.; Wachtveitl, J.; Broos, J.; Bredenbeck, J. Cyano-tryptophans as dual infrared and fluorescence spectroscopic labels to assess structural dynamics in proteins. Phys. Chem. Chem. Phys. 2018, 20, 19906–19915. [Google Scholar]
- Schmitz, A.J.; Hogle, D.G.; Gai, X.S.; Fenlon, E.E.; Brewer, S.H.; Tucker, M.J. Two-dimensional infrared study of vibrational coupling between azide and nitrile reporters in a RNA nucleoside. J. Phys. Chem. B 2016, 120, 9387–9394. [Google Scholar] [CrossRef] [PubMed]
- Getahun, Z.; Huang, C.Y.; Wang, T.; De León, B.; DeGrado, W.F.; Gai, F. Using nitrile-derivatized amino acids as infrared probes of local environment. J. Am. Chem. Soc. 2003, 125, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Fafarman, A.T.; Webb, L.J.; Chuang, J.I.; Boxer, S.G. Site-specific conversion of cysteine thiols into thiocyanate creates an IR probe for electric fields in proteins. J. Am. Chem. Soc. 2006, 128, 13356–13357. [Google Scholar] [CrossRef] [PubMed]
- Waegele, M.M.; Tucker, M.J.; Gai, F. 5-Cyanotryptophan as an infrared probe of local hydration status of proteins. Chem. Phys. Lett. 2009, 478, 249–253. [Google Scholar] [CrossRef]
- Bagchi, S.; Fried, S.D.; Boxer, S.G. A solvatochromic model calibrates nitriles’ vibrational frequencies to electrostatic fields. J. Am. Chem. Soc. 2012, 134, 10373–10376. [Google Scholar] [CrossRef]
- Ding, B.; Hilaire, M.R.; Gai, F. Infrared and fluorescence assessment of protein dynamics: From folding to function. J. Phys. Chem. B 2016, 120, 5103–5113. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectrscopy, 2nd ed.; Springer: New York, NY, USA, 1999. [Google Scholar]
- Seidel, C.A.M.; Schulz, A.; Sauer, M.H.M. Nucleobase-specific quenching of fluorescent dyes. 1. Nucleobase one-electron redox potentials and their correlation with static and dynamic quenching efficiencies. J. Phys. Chem. 1996, 100, 5541–5553. [Google Scholar] [CrossRef]
- Heinlein, T.; Knemeyer, J.-P.; Piestert, O.; Sauer, M. Photoinduced electron transfer between fluorescent dyes and guanosine residues in DNA-hairpins. J. Phys. Chem. B 2003, 107, 7957–7964. [Google Scholar] [CrossRef]
- Jing, M.; Bowser, M.T. Methods for measuring aptamer-protein equilibria: A review. Anal. Chim. Acta 2011, 686, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Peters, T.J. Serum albumin: Advances in protein chemistry, 1st ed.; Academic Press: New York, NY, USA, 1985. [Google Scholar]
- Cattan, D.; Bourgoin, D.; Joly, M. Change of conformation of DNA by association with proteins. Eur. J. Biochem. 1969, 8, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Taskent-Sezgin, H.; Marek, P.; Thomas, R.; Goldberg, D.; Chung, J.; Carrico, I.; Raleigh, D.P. Modulation of p-cyanophenylalanine fluorescence by amino acid side chains and rational design of fluorescence probes of α-helix formation. Biochemistry 2010, 49, 6290–6295. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, J.M.; Speight, L.C.; Fegley, M.W.; Petersson, E.J. Minimalist probes for studying protein dynamics: Thioamide quenching of selectively excitable fluorescent amino acids. J. Am. Chem. Soc. 2012, 134, 6088–6091. [Google Scholar] [CrossRef] [PubMed]
- Mintzer, M.R.; Troxler, T.; Gai, F. p-Cyanophenylalanine and selenomethionine constitute a useful fluorophore-quencher pair for short distance measurements: Application to polyproline peptides. Phys. Chem. Chem. Phys. 2015, 17, 7881–7887. [Google Scholar] [PubMed]
- Peters, T.J. All About Albumin: Biochemistry, Genetics and Medical Applications; Academic Press: Cambridge, MA, USA, 1996. [Google Scholar]
- Clarkson, J.; Smith, W.E. A DFT analysis of the vibrational spectra of nitrobenzene. J. Mol. Struct. 2003, 655, 413–422. [Google Scholar] [CrossRef]
- Seuvre, A.M.; Mathlouthi, M. F.T.-I.R. spectra of oligo- and poly-nucleotides. Carbohydr. Res. 1987, 169, 83–103. [Google Scholar] [CrossRef]
- Wood, B.R. The importance of hydration and DNA conformation in interpreting infrared spectra of cells and tissues. Chem. Soc. Rev. 2016, 45, 1980–1998. [Google Scholar] [CrossRef]
- Zhang, W.; Markiewicz, B.N.; Doerksen, R.S.; Smith, A.B.; Gai, F. C≡N stretching vibration of 5-cyanotryptophan as an infrared probe of protein local environment: What determines its frequency? Phys. Chem. Chem. Phys. 2016, 18, 7027–7034. [Google Scholar]
- Kamlet, M.J.; Abboud, J.L.M.; Abraham, M.H.; Taft, R.W. Linear solvation energy relationships. 23. A comprehensive collection of the solvatochromic parameters, π*, α, and β, and some methods for simplifying the generalized solvatochromic equation. J. Org. Chem. 1983, 48, 2877–2887. [Google Scholar] [CrossRef]
- Weber, R.J.; Liang, S.I.; Selden, N.S.; Desai, T.A.; Gartner, Z.J. Efficient targeting of fatty-acid modified oligonucleotides to live cell membranes through stepwise assembly. Biomacromolecules 2014, 15, 4621–4626. [Google Scholar] [CrossRef] [PubMed]
- Tucker, M.J.; Oyola, R.; Gai, F. Conformational distribution of a 14-residue peptide in solution: A fluorescence resonance energy transfer study. J. Phys. Chem. B 2005, 109, 4788–4795. [Google Scholar] [CrossRef] [PubMed]
- Holz, B.; Klimasauskasm, S.; Serva, S.; Weinhold, E. 2-Aminopurine as a fluorescent probe for DNA base flipping by methyltransferases. Nucleic Acids Res. 1998, 26, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds within this manuscript are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solvent | ω0, cm−1 | FWHM, cm−1 | π* | β | α | σ |
|---|---|---|---|---|---|---|
| Water (H2O) | 2232.3 | 11.1 | 1.09 | 0.47 | 1.51 | 0.05 |
| Methanol (MeOH) | 2230.8 | 9.6 | 0.6 | 0.62 | 0.93 | 0.29 |
| Dimethyl sulfoxide (DMSO) | 2223.7 | 7.4 | 1 | 0.76 | 0 | 1.76 |
| Acetonitrile (ACN) | 2226.8 | 10.1 | 0.75 | 0.31 | 0.19 | 0.87 |
| Tetrahydrofuran (THF) | 2225.8 | 7.17 | 0.58 | 0.55 | 0 | 1.13 |
| Trifluoroethanol (TFE) | 2235.4 | 22.6 | 0.73 | 0 | 1.51 | −0.78 |
| Chloroform (CHCl3) | 2228.2 | 11.2 | 0.53 | 0.1 | 0.2 | 0.43 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, I.A.; Acharyya, A.; Eng, C.M.; Rodgers, J.M.; DeGrado, W.F.; Jo, H.; Gai, F. 4-Cyanoindole-2′-deoxyribonucleoside as a Dual Fluorescence and Infrared Probe of DNA Structure and Dynamics. Molecules 2019, 24, 602. https://doi.org/10.3390/molecules24030602
Ahmed IA, Acharyya A, Eng CM, Rodgers JM, DeGrado WF, Jo H, Gai F. 4-Cyanoindole-2′-deoxyribonucleoside as a Dual Fluorescence and Infrared Probe of DNA Structure and Dynamics. Molecules. 2019; 24(3):602. https://doi.org/10.3390/molecules24030602
Chicago/Turabian StyleAhmed, Ismail A., Arusha Acharyya, Christina M. Eng, Jeffrey M. Rodgers, William F. DeGrado, Hyunil Jo, and Feng Gai. 2019. "4-Cyanoindole-2′-deoxyribonucleoside as a Dual Fluorescence and Infrared Probe of DNA Structure and Dynamics" Molecules 24, no. 3: 602. https://doi.org/10.3390/molecules24030602
APA StyleAhmed, I. A., Acharyya, A., Eng, C. M., Rodgers, J. M., DeGrado, W. F., Jo, H., & Gai, F. (2019). 4-Cyanoindole-2′-deoxyribonucleoside as a Dual Fluorescence and Infrared Probe of DNA Structure and Dynamics. Molecules, 24(3), 602. https://doi.org/10.3390/molecules24030602