
Characterization and Antibacterial Activity of 7S and 11S Globulins Isolated from Cowpea Seed Protein

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
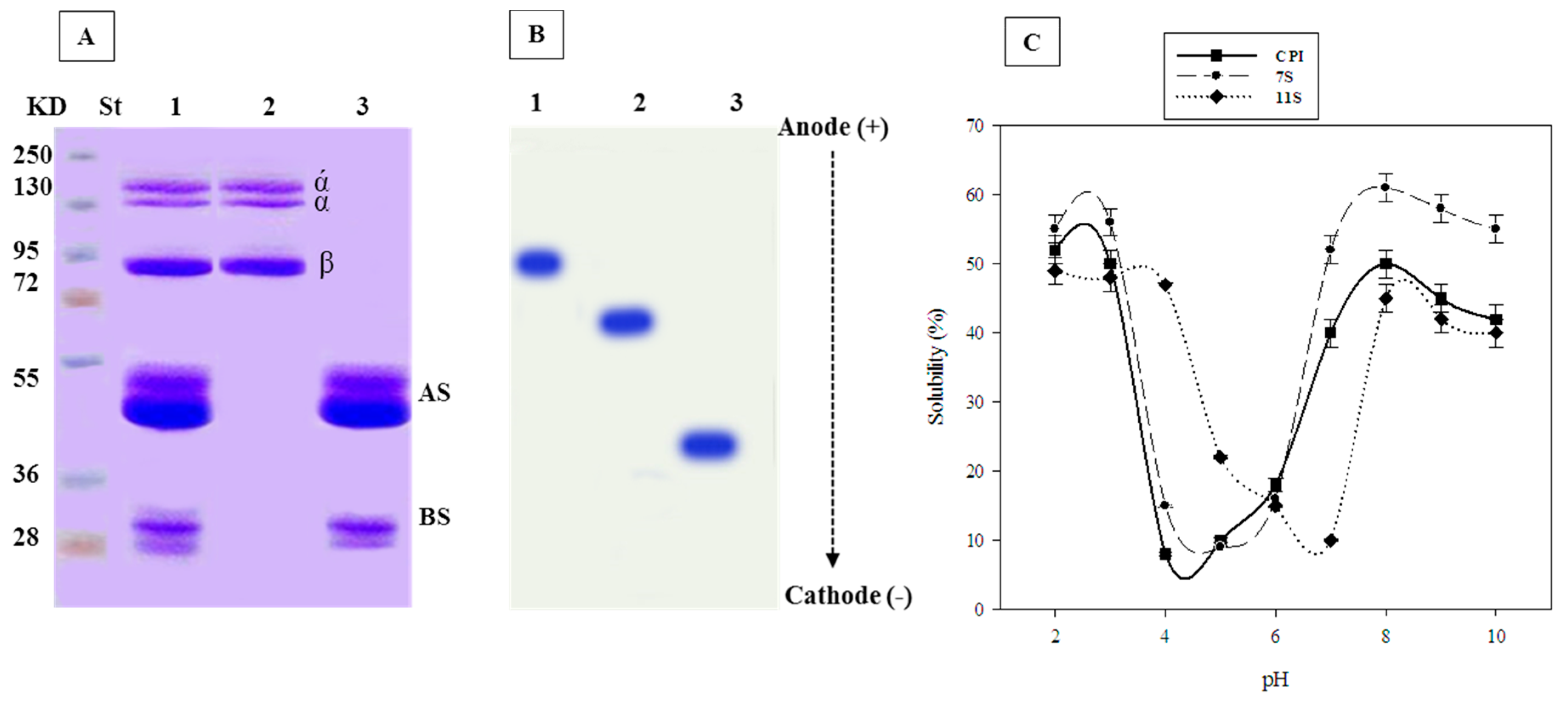
2.1. SDS-PAGE of 7S and 11S Globulins
2.2. Urea-PAGE of 7S and 11S Globulins
2.3. pH-Solubility Curve of 7S and 11S Globulins:
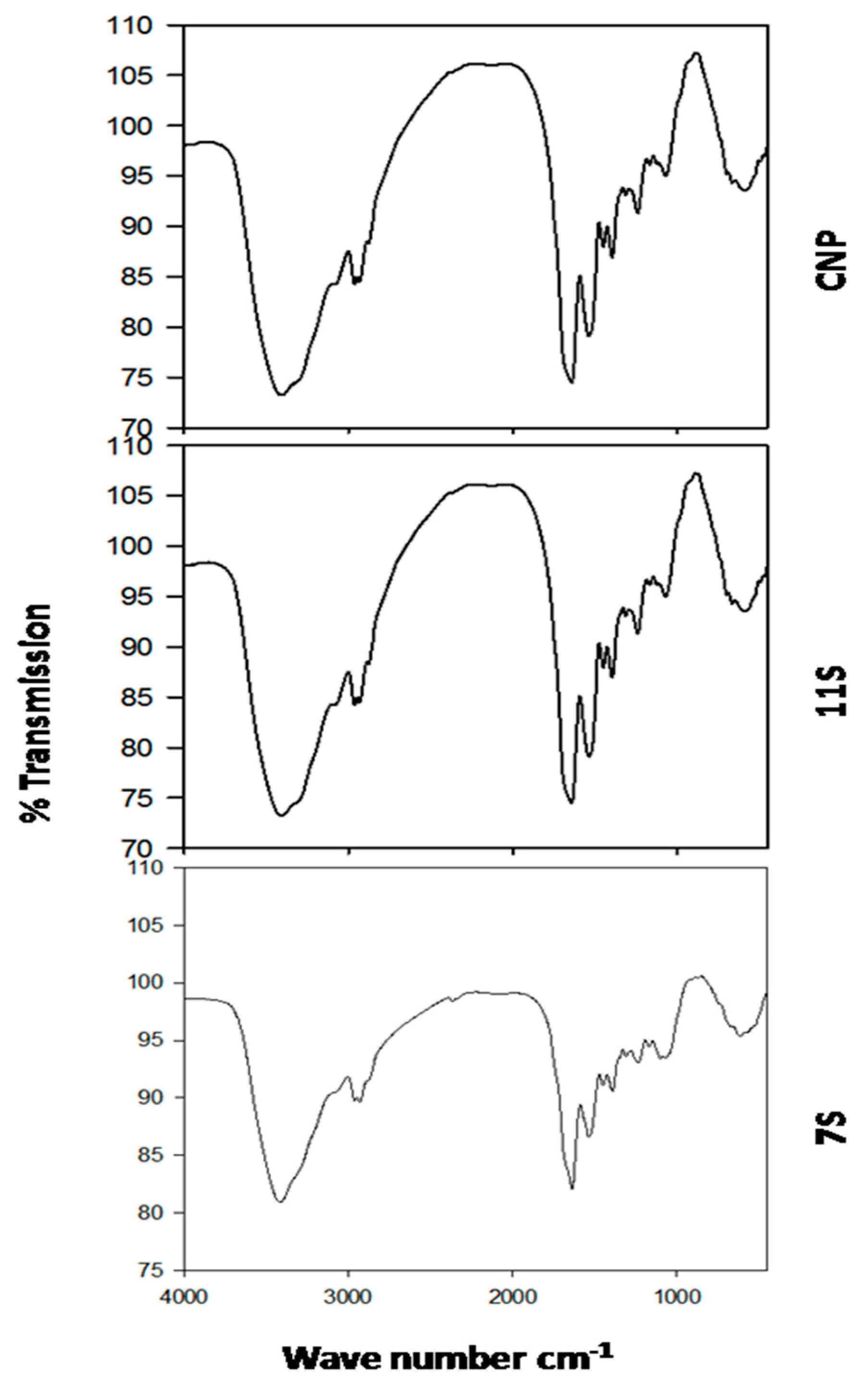
2.4. Fourier Transform Infrared (FT-IR) Spectroscopy of 7S and 11S Globulins
2.5. Antibacterial Activity and MIC of 7S and 11S Globulins
2.6. Combined Effect of Cip-7S and Cip-11S against the Tested Bacteria
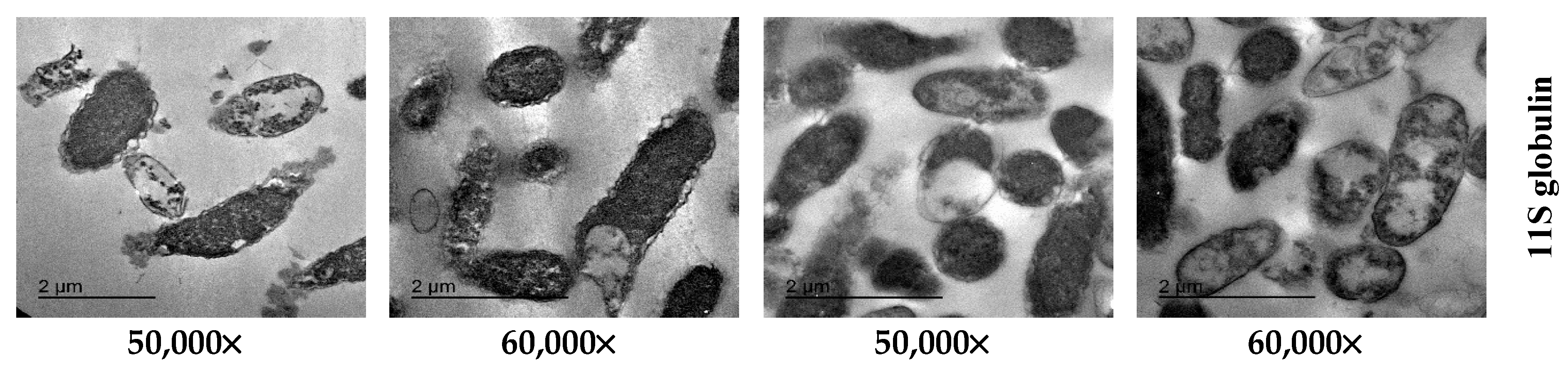
2.7. TEM Image Analysis
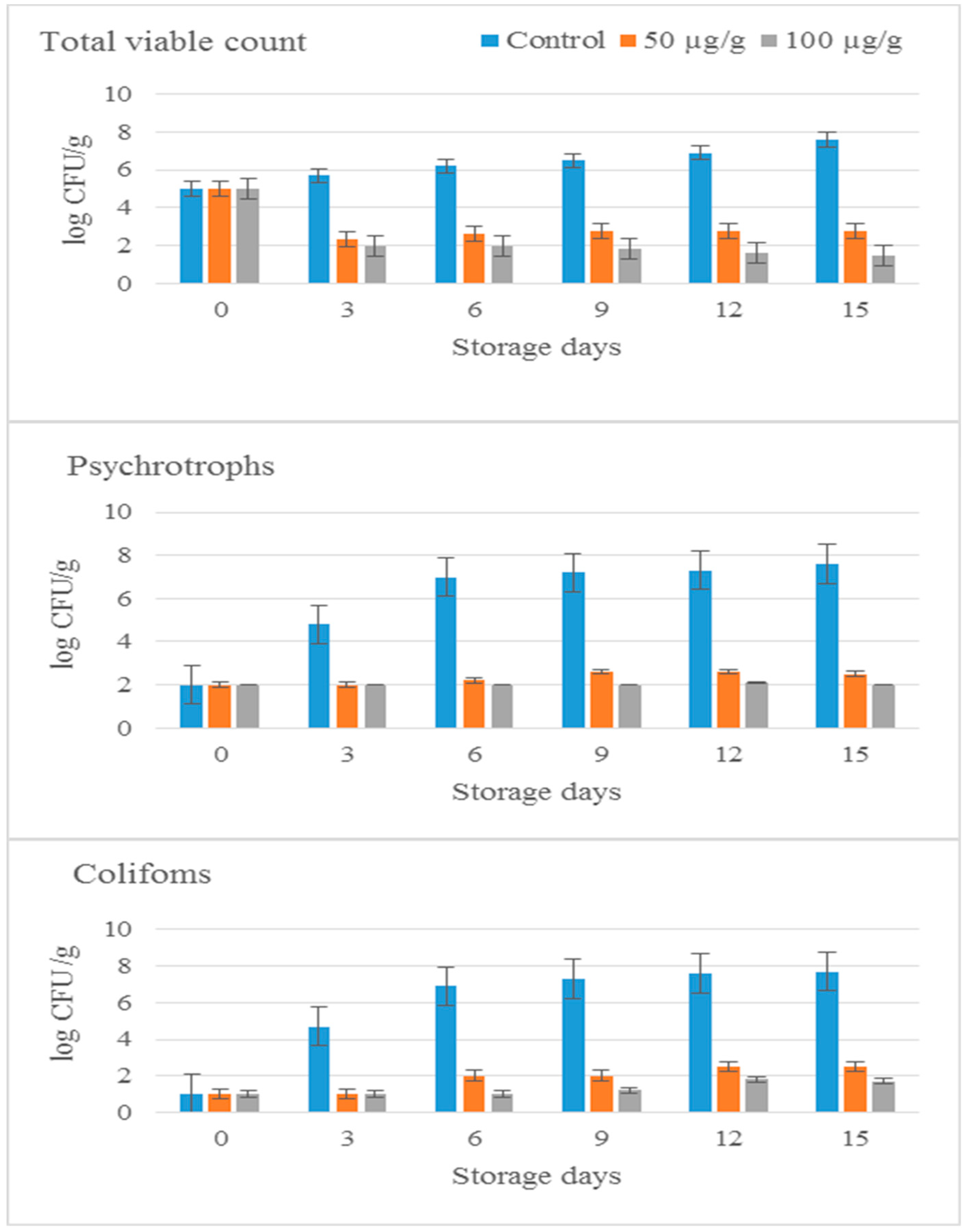
2.8. Storage of Minced Beef with 11S Globulin from Cowpea at 4 °C
Microbial Analysis
2.9. Physicochemical Analysis of Meat Sample
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Microorganisms
4.3. Isolation and Purification of Globulins from Cowpea (Vigna unguiculata) Seed
4.3.1. Fractionation of Papain Hydrolysed Cowpea
4.3.2. Fractionation of PHC by Size Exclusion Chromatography (SEC)
4.4. Characterization of 7S and 11S Globulins
4.4.1. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.4.2. Urea-PAGE
4.4.3. pH-Solubility Curve
4.4.4. Fourier Transform Infrared (FT-IR) Spectroscopy
4.5. Antibacterial Activity of 7S and 11S Globulins
4.6. Determination of MIC Values of both 7S and 11S Globulins:
4.7. Antibacterial Activity of Combinations between the Ciprofloxacin Antibiotic and the Globulins
4.8. Transmission Electron Microscopy (TEM)
4.9. Storage of Minced Beef with 11S Globulin Isolated from Cowpea at 4 °C
4.9.1. Meat Samples
4.9.2. Experimental Design
4.9.3. Microbial Analysis
4.10. Physicochemical Analysis of Meat Sample
4.10.1. pH Determination
4.10.2. Metmyoglobin (MetMb) Analysis
4.10.3. Lipid Peroxidation Assay
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Enan, G. Nature and phenotypic characterization of plantaricin UG1 resistance in Listeria monocytogenes LMG10470. J. Food Agric. Environ. 2006, 4, 105–108. [Google Scholar]
- Enan, G. Control of the regrowing bacteriocin resistant variants of Listeria monocytogenes LMG10470 in vitro and in food by nisin-plantaricin UG1 mixture. Biotechnology 2006, 5, 143–147. [Google Scholar]
- Abdel-Shafi, S.; Ouda, S.M.; El-Balaty, I.; Enan, G. Characterization and identification of multidrug resistant bacteria from Egyptian patient. Biotechnology 2013, 12, 65–73. [Google Scholar] [CrossRef]
- Kapoor, G.; Saurabh Saigal1, S.; Elongavan, A. Action and resistance mechanisms of antibiotics: A guide for Clinicians. J. Anaesthesiol. Clin. Pharmacol. 2017, 33, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hamid, M.; Goda, H.A.; De Gobba, C.; Jenssen, H.; Osman, A. Antibacterial activity of papain hydrolysed camel whey and its fractions. Int. Dairy J. 2016, 61, 91–98. [Google Scholar] [CrossRef]
- Osman, A.; Goda, H.A.; Abdel-Hamid, M.; Badran, S.M.; Otte, J. Antibacterial peptides generated by Alcalase hydrolysis of goat whey. LWT-Food Sci. Technol. 2016, 65, 480–486. [Google Scholar] [CrossRef]
- Abdel-Shafi, S.; Osman, A.; Enan, G.; El-Nemer, M.; Sitohy, M. Antibacterial activity of methylated egg white proteins against pathogenic G+ and G− bacteria matching antibiotics. SpringerPlus 2016, 5, 983. [Google Scholar] [CrossRef]
- Osman, A.; El-Araby, G.M.; Taha, H. Potential use as a bio-preservative from lupin protein hydrolysate generated by alcalase in food system. J. Appl. Biol. Biotechnol. 2016, 4, 076–081. [Google Scholar]
- Sitohy, M.; Mahgoub, S.; Osman, A.; El-Masry, R.; Al-Gaby, A. Extent and mode of action of cationic legume proteins against Listeria monocytogenes and Salmonella enteritidis. Probiot. Antimicrob. Proteins 2013, 5, 195–205. [Google Scholar] [CrossRef]
- Sitohy, M.; Osman, A. Antimicrobial activity of native and esterified legume proteins against Gram-negative and Gram-positive bacteria. Food Chem. 2010, 120, 66–73. [Google Scholar] [CrossRef]
- Sitohy, M.; Osman, A.; Ghany, A.G.A.; Salama, A. Antibacterial phycocyanin from Anabaena oryzae SOS13. Int. J. Appl. Res. Nat. Prod. 2015, 8, 27–36. [Google Scholar]
- Mahgoub, S.; Osman, A.; Sitohy, M. Inhibition of growth of pathogenic bacteria in raw milk by legume protein esters. J. Food Prot. 2011, 74, 1475–1481. [Google Scholar] [CrossRef]
- Mahgoub, S.A.; Osman, A.O.; Sitohy, M.Z. Impeding Bacillus spore germination in vitro and in milk by soy glycinin during long cold storage. J. Gen. Appl. Microbiol. 2016, 62, 52–59. [Google Scholar] [CrossRef]
- Mahgoub, S.A.; Sitohy, M.Z.; Osman, A.O. Counteracting recontamination of pasteurized milk by methylated soybean protein. Food Bioprocess Technol. 2013, 6, 101–109. [Google Scholar] [CrossRef]
- Osman, A.; Daidamony, G.; Sitohy, M.; Khalifa, M.; Enan, G. Soybean glycinin basic subunit inhibits methicillin resistant-vancomycin intermediate Staphylococcus aureus (MRSA-VISA) in vitro. Int. J. Appl. Res. Nat. Prod. 2016, 9, 17–26. [Google Scholar]
- Osman, A.; Mahgoub, S.; El-Masry, R.; Al-Gaby, A.; Sitohy, M. Extending the technological validity of raw buffalo milk at room temperature by esterified legume proteins. J. Food Process. Preserv. 2014, 38, 223–231. [Google Scholar] [CrossRef]
- Osman, A.; Mahgoub, S.; Sitohy, M. Hindering milk quality storage deterioration by mild thermization combined with methylated chickpea protein. Int. Food Res. J. 2014, 21, 693–701. [Google Scholar]
- Osman, A.O.; Mahgoub, S.A.; Sitohy, M.Z. Preservative action of 11S (glycinin) and 7S (β-conglycinin) soy globulin on bovine raw milk stored either at 4 or 25 C. J. Dairy Res. 2013, 80, 174–183. [Google Scholar] [CrossRef]
- Sitohy, M.; Osman, A. Bioactive Compounds in Soybean Proteins and Its Applications in Food Systems. In The Handbook of Environmental Chemistry; Springer: Cham, Switzerland, 2018; Volume 76. [Google Scholar]
- Sitohy, M.Z.; Osman, A.O. Enhancing milk preservation with esterified legume proteins. Probiot. Antimicrob. Proteins 2011, 3, 48–56. [Google Scholar] [CrossRef]
- Ehlers, J.; Hall, A. Cowpea (Vigna unguiculata L. Walp.). Field Crop. Res. 1997, 53, 187–204. [Google Scholar] [CrossRef]
- Liu, K. Soybeans: Chemistry, Technology, and Utilization; Springer: New York, NY, USA, 2012. [Google Scholar]
- Freitas, R.L.; Teixeira, A.R.; Ferreira, R.B. Characterization of the proteins from Vigna unguiculata seeds. J. Agric. Food Chem. 2004, 52, 1682–1687. [Google Scholar] [CrossRef]
- Sitohy, M.Z.; Mahgoub, S.A.; Osman, A.O. In vitro and in situ antimicrobial action and mechanism of glycinin and its basic subunit. Int. J. Food Microbiol. 2012, 154, 19–29. [Google Scholar] [CrossRef]
- Li, Y.-Q.; Sun, X.X.; Feng, J.L.; Mo, H.Z. Antibacterial activities and membrane permeability actions of glycinin basic peptide against Escherichia coli. Innov. Food. Sci. Emerg. Technol. 2015, 31, 170–176. [Google Scholar] [CrossRef]
- Yang, J.; Sun, G.J.; Li, Y.Q.; Cui, K.Y.; Mo, H.Z. Antibacterial characteristics of glycinin basic polypeptide against Staphylococcus aureus. Food Sci. Biotechnol. 2016, 25, 1477–1483. [Google Scholar] [CrossRef]
- Zhao, G.P.; Li, Y.Q.; Sun, G.J.; Mo, H.Z. Antibacterial actions of glycinin basic peptide against Escherichia coli. J. Agric. Food Chem. 2017, 65, 5173–5180. [Google Scholar] [CrossRef]
- Barker, R.D.; Derbyshire, E.; Yarwood, A.; Boulter, D. Purification and characterization of the major storage proteins of Phaseolus vulgaris seeds, and their intracellular and cotyledonary distribution. Phytochemistry 1976, 15, 751–757. [Google Scholar] [CrossRef]
- Lombardi, L.; Maisetta, G.; Batoni, G.; Tavanti, A. Insights into the antimicrobial properties of hepcidins: Advantages and drawbacks as potential therapeutic agents. Molecules 2015, 20, 6319–6341. [Google Scholar] [CrossRef]
- Wang, J.X.; Zhao, X.F.; Liang, Y.L.; Li, L.; Zhang, W.; Ren, Q.; Wang, L.C.; Wang, L.Y. Molecular characterization and expression of the antimicrobial peptide defensin from the housefly (Musca domestica). Cell. Mol. Life Sci. 2006, 63, 3072–3082. [Google Scholar] [CrossRef]
- Barari, L.; Mosavi, N.; Asgharpour, F. Antibacterial and antifungal effect of chickpea (cicer arietinum) aqueous seed extract. World J. Pharm. Pharm. Sci. 2015, 4, 336–346. [Google Scholar]
- Chan, M.; Chan, L.; Ng, S.; Liu, Q. Heavy metal contents and magnetic properties of street dust in two districts of different traffic density in Hong Kong. Geophys. Solut. Environ. Eng. 2006, 1, 807–811. [Google Scholar]
- Nielsen, N.C. The structure and complexity of the 11S polypeptides in soybeans. J. Am. Oil. Chem. Soc. 1985, 62, 1680–1686. [Google Scholar] [CrossRef]
- Osman, A.; Abbas, E.; Mahgoub, S.; Sitohy, M. Inhibition of Penicillium digitatum in vitro and in postharvest orange fruit by a soy protein fraction containing mainly β-conglycinin. J. Gen. Plant Pathol. 2016, 82, 293–301. [Google Scholar] [CrossRef]
- Sitohy, M.; Doheim, M.; Badr, H. Isolation and characterization of a lectin with antifungal activity from Egyptian Pisum sativum seeds. Food Chem. 2007, 104, 971–979. [Google Scholar] [CrossRef]
- Amaral, A.L.; Ferreira, E.S.; Neves, V.A.; Demonte, A. Legumin from chickpea: Hypolipidemic effect in the liver of hypercholesterolemic rats. Nutr. Food Sci. 2014, 44, 378–388. [Google Scholar] [CrossRef]
- Osman, A.; Goda, H.A.; Sitohy, M. Storage stability of minced beef supplemented with chickpea legumin at 4° C as a potential substitute for nisin. LWT-Food Sci. Technol. 2018, 93, 434–441. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, L.; Nie, S.; Qi, X.; Zhang, Q. Secondary structure of insulin encapsulated within liposomes. Acta Pharm. Sin. Yao Xue Xue Bao 2003, 38, 863–866. [Google Scholar]
- Hancock, R.E.; Rozek, A. Role of membranes in the activities of antimicrobial cationic peptides. FEMS Microbiol. Lett. 2002, 206, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, H.R.; Thomas, U.; Pellegrini, A. A helix-loop-helix peptide at the upper lip of the active site cleft of lysozyme confers potent antimicrobial activity with membrane permeabilization action. J. Biol. Chem. 2001, 276, 43767–43774. [Google Scholar] [CrossRef]
- Bals, R.; Wilson, J. Cathelicidins-a family of multifunctional antimicrobial peptides. Cell. Mol. Life Sci. 2003, 60, 711–720. [Google Scholar] [CrossRef]
- Mahgoub, S.A.M.; Osman, A.; Ramadan, M.F. Inhibitory effect of Nigella sativa oil against Listeria monocytogenes and Salmonella enteritidis inoculated in minced beef meat. J. Food Meas. Charact. 2017, 11, 2043–2051. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.W.; Williams, J. The electrophoresis of transferrins in urea/polyacrylamide gels. Biochem. J. 1980, 189, 541–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chobert, J.-M.; Touati, A.; Bertrand-Harb, C.; Dalgalarrondo, M.; Nicolas, M.-G.; Haertle, T. In vitro proteolysis and functional properties of reductively alkylated β-casein derivatives. J. Dairy Res. 1991, 58, 285–298. [Google Scholar] [CrossRef]
- Souillac, P.O.; Middaugh, C.R.; Rytting, J.H. Investigation of protein/carbohydrate interactions in the dried state. 2. Diffuse reflectance FTIR studies. Int. J. Pharm. 2002, 235, 207–218. [Google Scholar] [CrossRef]
- Surewicz, W.K.; Mantsch, H.H. New insight into protein secondary structure from resolution-enhanced infrared spectra. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1988, 952, 115–130. [Google Scholar] [CrossRef]
- Bauer, A.; Kirby, W.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Karnovsky, M.J. A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron microscopy. J. Cell Biol. 1965, 27, 1A–149A. [Google Scholar]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208. [Google Scholar] [CrossRef]
- Association, A.P.H. Compendium of Methods for the Microbiological Examination of Foods, 3rd ed.; APHA Inc.: Washington, DC, USA, 1992; Volume 27, p. 2013. [Google Scholar]
- Goldman, E.; Green, L.H. Practical Handbook of Microbiology; CRC Press: Baco Raton, FL, USA, 2015. [Google Scholar]
- Özyurt, G.; Kuley, E.; Balikçi, E.; Kaçar, Ç.; Gökdogan, S.; Etyemez, M.; Özogul, F. Effect of the icing with rosemary extract on the oxidative stability and biogenic amine formation in sardine (Sardinella aurita) during chilled storage. Food Bioprocess Technol. 2012, 5, 2777–2786. [Google Scholar] [CrossRef]
- Badr, H.M. Antioxidative activity of carnosine in gamma irradiated ground beef and beef patties. Food Chem. 2007, 104, 665–679. [Google Scholar] [CrossRef]
- Krzywicki, K. The determination of haem pigments in meat. Meat Sci. 1982, 7, 29–36. [Google Scholar] [CrossRef]
- Niehaus, W., Jr.; Samuelsson, B. Formation of malonaldehyde from phospholipid arachidonate during microsomal lipid peroxidation. Eur. J. Biochem. 1968, 6, 126–130. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | Protein Subunit Concentration (µg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| 10 | 25 | 50 | 100 | 200 | 400 | 800 | 1000 | |
| Inhibition Zone Diameter (mm) | ||||||||
| 7S globulin | ||||||||
| S. typhimurium | - | - | - | 10 ± 0.58 | 10.3 ± 0.75 | 10.6 ± 0.57 | 13 ± 1.15 | 19 ± 0.15 |
| K. pneumoniae | - | - | - | - | 9 ± 0.57 | 10.3 ± 0.58 | 10.6 ± 0.57 | 11.6 ± 0.57 |
| S. pyogenes | - | - | - | 10.3 ± 0.75 | 11.3 ± 0.74 | 13.3 ± 0.75 | 11.6 ± 0.75 | 13.6 ± 0.92 |
| L.monocytogenes | 13.6 ± 0.35 | 15 ± 1.15 | 15 ± 1.15 | 16 ± 1.15 | 19.3 ± 1.15 | 20 ± 1.15 | 20 ± 1.14 | 23 ± 1.73 |
| L. ivanovii | - | - | - | 9 ± 0.58 | 10.6 ± 0.92 | 11 ± 0.57 | 13 ± 0.88 | 18 ± 1.15 |
| P. aeruginosa | - | - | 10.3 ± 0.58 | 11 ± 0.86 | 12.6 ± 1.50 | 17 ± 1.15 | 18 ± 0.86 | 19 ± 0.86 |
| S. aureus | - | - | - | 0.0 ± 0.0 | 9 ± 0.56 | 10.6 ± 0.58 | 14.6 ± 0.57 | 15 ± 0.58 |
| E. coli | - | - | - | - | - | - | - | - |
| 11S globulin | ||||||||
| S. typhi | - | - | - | - | 12.6 ± 0.92 | 13.6 ± 0.92 | 13 ± 1.15 | 23 ± 1.45 |
| k. pneumonia | - | - | - | - | 0.0 ± 0.0 | 9.3 ± 0.58 | 9.6 ± 0.57 | 14 ± 0.86 |
| St. pyogenes | - | - | - | - | 8 ± 0.57 | 10 ± 0.57 | 11 ± 0.86 | 13.6 ± 0.58 |
| L. monocytogens | - | - | - | - | 7.6 ± 0.35 | 9.3 ± 0.56 | 9.6 ± 0.57 | 13.6 ± 0.58 |
| L. ivanovii | - | 8.7 ± 0.40 | 11 ± 0.86 | 12.3 ± 0.75 | 15.3 ± 0.75 | 23 ± 1.73 | 19 ± 1.15 | 25 ± 1.73 |
| P. auriginosa | - | 9 ± 0.57 | 10.3 ± 0.58 | 11 ± 0.57 | 11.6 ± 0.58 | 13 ± 0.86 | 13.3 ± 0.75 | 15 ± 0.86 |
| S. aureus | - | - | 14.6 ± 1.15 | 15.3 ± 1.15 | 0.0 ± 0.0 | 24 ± 1.15 | 24 ± 1.15 | 25 ± 1.73 |
| E. coli | - | - | 9 ± 0.57 | 9.3 ± 0.57 | 10 ± 0.57 | 10.3 ± 0.58 | 10 ± 0.57 | 17.3 ± 1.15 |
| Concentration (µg/mL) | Inhibition Zone Diameter (mm) | ||
|---|---|---|---|
| 7S | 11S | ||
| Against | |||
| Cip | Protein | L. monocytogenes | S. aureus |
| 100 | 0 | 15.3 ± 0.58 | 11.3 ± 0.57 |
| 80 | 20 | 17.0 ± 0.69 | 17.6 ± 0.93 |
| 60 | 40 | 17.3 ± 0.75 | 20.0 ± 1.15 |
| 40 | 60 | 19.4 ± 0.31 | 28.0 ± 1.15 |
| 20 | 80 | 20.8 ± 0.51 | 30.0 ± 1.44 |
| 0 | 100 | 24.5 ± 0.52 | 32.6 ± 1.50 |
| Storage Time (day) | Control | 11S (µg/g) | ||
|---|---|---|---|---|
| 50 | 100 | |||
| pH Values | ||||
| 0 | 5.6 | 5.6 | 5.6 | |
| 3 | 5.9 | 5.7 | 5.7 | |
| 6 | 6.5 | 5.8 | 5.75 | |
| 9 | 6.6 | 6.3 | 5.8 | |
| 12 | 6.9 | 6.4 | 6.1 | |
| 15 | 7.2 | 6.55 | 6.3 | |
| Metmyoglobin (%) | ||||
| 0 | 11 | 11 | 11 | |
| 3 | 34.6 | 24 | 20 | |
| 6 | 50.6 | 30 | 25 | |
| 9 | 55 | 39 | 30 | |
| 12 | 63 | 45 | 32 | |
| 15 | 69 | 50 | 45 | |
| Lipid Oxidation Inhibition (%) | ||||
| 0 | 25 | 25 | 25 | |
| 3 | 23 | 24 | 24 | |
| 6 | 22 | 23 | 24 | |
| 9 | 19 | 22 | 23 | |
| 12 | 13 | 18 | 20 | |
| 15 | 10 | 16 | 17 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Shafi, S.; Al-Mohammadi, A.-R.; Osman, A.; Enan, G.; Abdel-Hameid, S.; Sitohy, M. Characterization and Antibacterial Activity of 7S and 11S Globulins Isolated from Cowpea Seed Protein. Molecules 2019, 24, 1082. https://doi.org/10.3390/molecules24061082
Abdel-Shafi S, Al-Mohammadi A-R, Osman A, Enan G, Abdel-Hameid S, Sitohy M. Characterization and Antibacterial Activity of 7S and 11S Globulins Isolated from Cowpea Seed Protein. Molecules. 2019; 24(6):1082. https://doi.org/10.3390/molecules24061082
Chicago/Turabian StyleAbdel-Shafi, Seham, Abdul-Raouf Al-Mohammadi, Ali Osman, Gamal Enan, Samar Abdel-Hameid, and Mahmoud Sitohy. 2019. "Characterization and Antibacterial Activity of 7S and 11S Globulins Isolated from Cowpea Seed Protein" Molecules 24, no. 6: 1082. https://doi.org/10.3390/molecules24061082
APA StyleAbdel-Shafi, S., Al-Mohammadi, A. -R., Osman, A., Enan, G., Abdel-Hameid, S., & Sitohy, M. (2019). Characterization and Antibacterial Activity of 7S and 11S Globulins Isolated from Cowpea Seed Protein. Molecules, 24(6), 1082. https://doi.org/10.3390/molecules24061082