
Identification of a Recombinant Human Interleukin-12 (rhIL-12) Fragment in Non-Reduced SDS-PAGE


Abstract
:1. Introduction
2. Results and Discussion
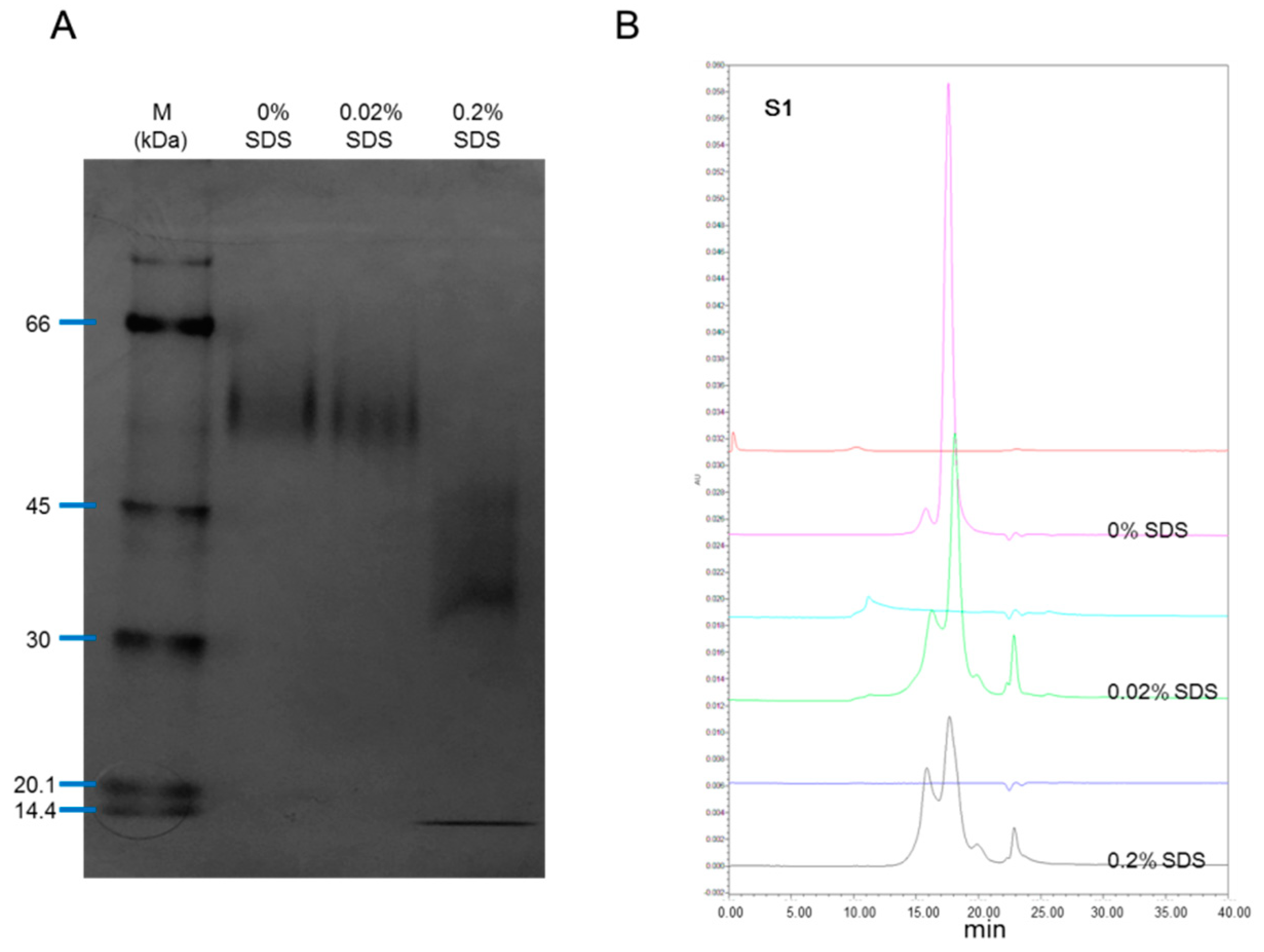
2.1. Purity Determination of rhIL-12 Samples by Non-Reduced SDS-PAGE and SEC-HPLC
2.2. Identification of rhIL-12 Fragment by UPLC/MS and N-Terminal Sequencing
2.3. Effect of Denaturant on rhIL-12 Pattern in Native PAGE and SEC-HPLC
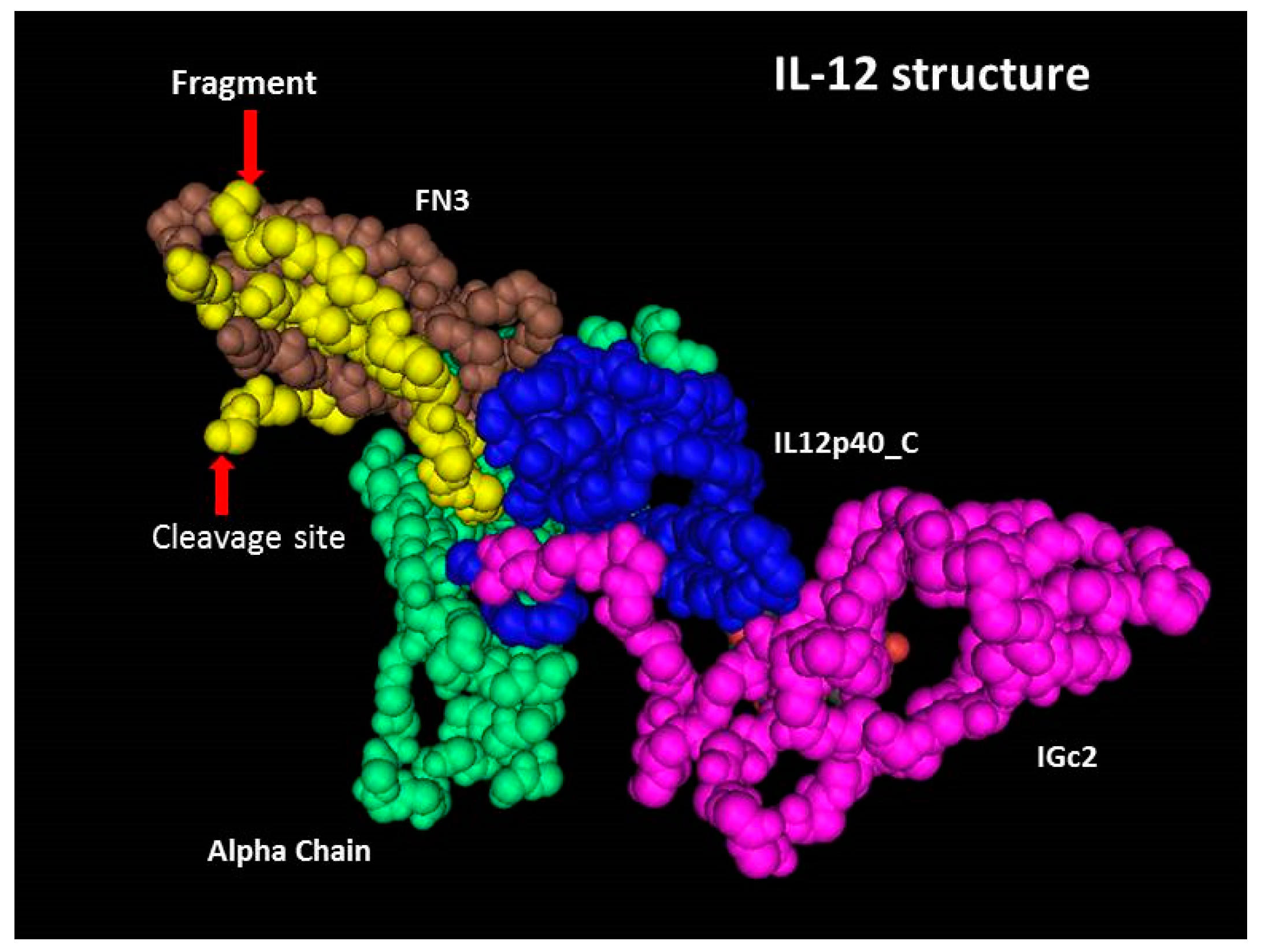
2.4. Cleavage Site in 3D Structure of rhIL-12
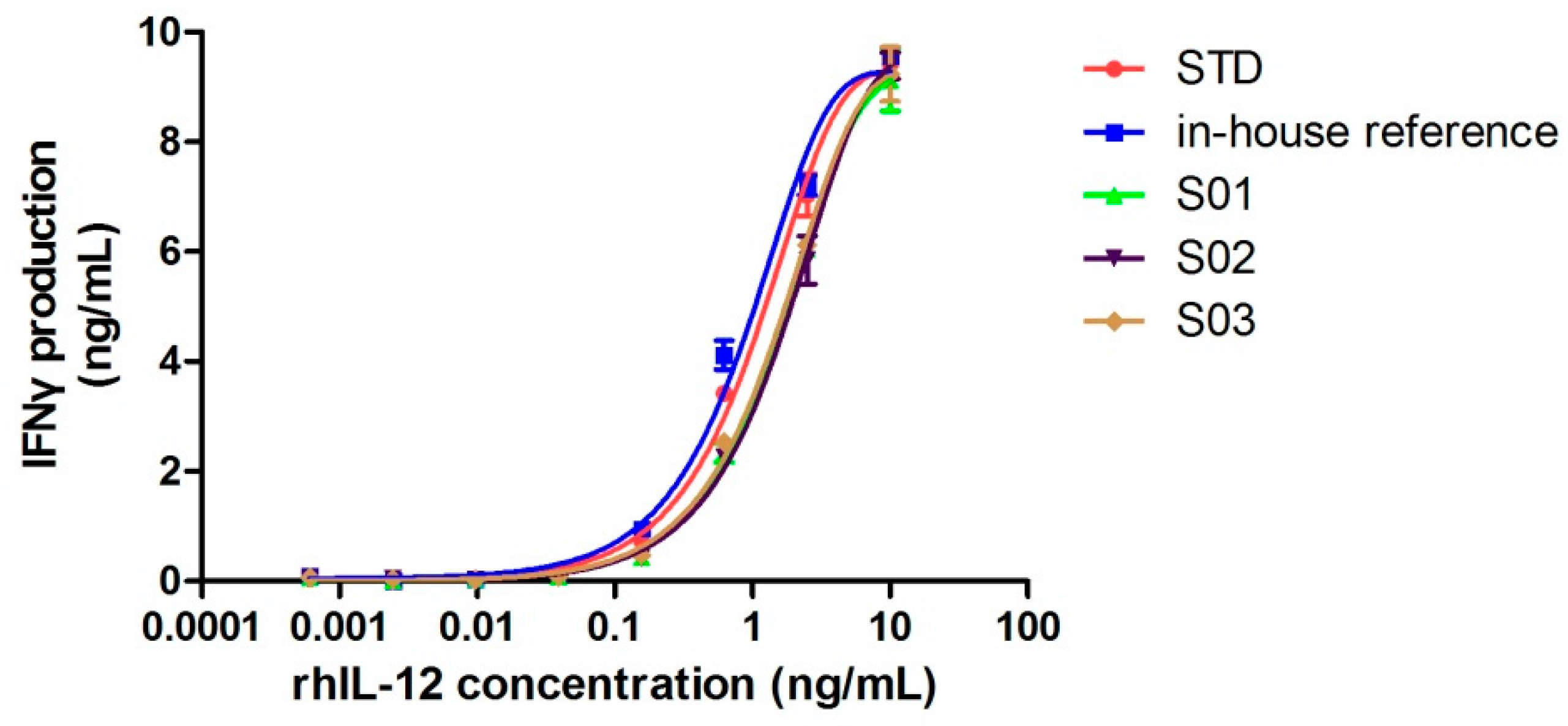
2.5. Influence of Cleavage on Bioactivity
3. Materials and Methods
3.1. Materials
3.2. Electrophoresis Analysis
3.3. Size-Exclusion Chromatography Analysis
3.4. UPLC/MS
3.5. N-Terminal Sequencing
3.6. Measurement of rhIL-12 Concentration and Bioactivity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hamza, T.; Barnett, J.B.; Li, B. Interleukin 12 a key immunoregulatory cytokine in infection applications. Int. J. Mol. Sci. 2010, 11, 789–806. [Google Scholar] [CrossRef]
- Sun, L.; He, C.; Nair, L.; Yeung, J.; Egwuagu, C.E. Interleukin 12 (IL-12) family cytokines: Role in immune pathogenesis and treatment of CNS autoimmune disease. Cytokine 2015, 75, 249–255. [Google Scholar] [CrossRef] [Green Version]
- Braun, M.; Ress, M.L.; Yoo, Y.E.; Scholz, C.J.; Eyrich, M.; Schlegel, P.G.; Wolfl, M. IL12-mediated sensitizing of T-cell receptor-dependent and -independent tumor cell killing. Oncoimmunology 2016, 5, e1188245. [Google Scholar] [CrossRef] [Green Version]
- Del Vecchio, M.; Bajetta, E.; Canova, S.; Lotze, M.T.; Wesa, A.; Parmiani, G.; Anichini, A. Interleukin-12: Biological properties and clinical application. Clin. Cancer Res. 2007, 13, 4677–4685. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.Y.; Zhan, M.Y.; Guo, M.Z.; Yi, Y.; Cong, Y.; Tian, R.G.; Zhang, W.Y.; Bi, S.L. Expression of human IL-12 in mammalian cell and study on its biological activities. Chin. J. Exp. Clin. Virol. 2007, 21, 235–237. [Google Scholar]
- Zundler, S.; Neurath, M.F. Interleukin-12: Functional activities and implications for disease. Cytokine Growth Factor Rev. 2015, 26, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Wang, W.Q.; Gong, X.J.; Gao, L.; Yang, L.R.; Yu, W.N.; Shen, H.Y.; Wan, L.Q.; Jia, X.F.; Wang, Y.S.; et al. Study of recombinant human interleukin-12 for treatment of complications after radiotherapy for tumor patients. World J. Clin. Oncol. 2017, 8, 158–167. [Google Scholar] [CrossRef]
- Berraondo, P.; Etxeberria, I.; Ponz-Sarvise, M.; Melero, I. Revisiting interleukin-12 as a caner immunotherapy agent. Clin. Cancer Res. 2018, 24, 2716–2718. [Google Scholar] [CrossRef]
- Tugues, S.; Burkhard, S.H.; Ohs, I.; Vrohlings, M.; Nussbaum, K.; Vom Berg, J.; Kulig, P.; Becher, B. New insights into IL-12-mediated tumor suppression. Cell Death Differ. 2015, 22, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Jayanthi, S.; Koppolu, B.P.; Smith, S.G.; Jalah, R.; Bear, J.; Rosati, M.; Pavlakis, G.N.; Felber, B.K.; Zaharoff, D.A.; Kumar, T.K. Efficient production and purification of recombinant human interleukin-12 (IL-12) overexpressed in mammalian cells without affinity tag. Protein Expr. Purif. 2014, 102, 76–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Li, X.; Wang, J.; Gao, D.; Li, Y.; Li, H.; Chu, Y.; Zhang, Z.; Liu, H.; Jiang, G.; et al. Re-designing Interleukin-12 to enhance its safety and potential as an anti-tumor immunotherapeutic agent. Nat. Commun. 2017, 8, 1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, K.M.; Lan, Y.; Lauder, S.; Zhang, J.; Brunkhorst, B.; Qin, G.; Verma, R.; Courtenay-Luck, N.; Gillies, S.D. huBC1-IL12, an immunocytokine which targets EDB-containing oncofetal fibronectin in tumors and tumor vasculature, shows potent anti-tumor activity in human tumor models. Cancer Immunol. Immunother. 2007, 56, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Sommavilla, R.; Pasche, N.; Trachsel, E.; Giovannoni, L.; Roesli, C.; Villa, A.; Neri, D.; Kaspar, M. Expression, engineering and characterization of the tumor-targeting heterodimeric immunocytokine F8-IL12. Protein Eng. Des. Sel. 2010, 23, 653–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, J.L.; Priya, T.A.; Hu, K.Y.; Yan, H.Y.; Shen, S.T.; Song, Y.L. Grouper interleukin-12, linked by an ancient disulfide-bond architecture, exhibits cytokine and chemokine activities. Fish Shellfish Immunol. 2014, 36, 27–37. [Google Scholar] [CrossRef]
- Yoon, C.; Johnston, S.C.; Tang, J.; Stahl, M.; Tobin, J.F.; Somers, W.S. Charged residues dominate a unique interlocking topography in the heterodimeric cytokine interleukin-12. Embo J. 2000, 19, 3530–3541. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, C.; Domingues, L. Guidelines to reach high-quality purified recombinant proteins. Appl. Microbiol. Biotechnol. 2018, 102, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Rogers, R.S.; Nightlinger, N.S.; Livingston, B.; Campbell, P.; Bailey, R.; Balland, A. Multi-attribute method for characterization, quality control testing and disposition of biologics. Mabs-Austin 2015, 7, 881–890. [Google Scholar] [CrossRef]
- Rogers, R.S.; Abernathy, M.; Richardson, D.D.; Rouse, J.C.; Sperry, J.B. A view on the importance of “multi-attribute method” for measuring purity of biopharmaceuticals and improving overall control strategy. AAPS J. 2017, 20, 7. [Google Scholar] [CrossRef]
- Reynolds, J.A.; Tanford, C. Binding of Dodecyl Sulfate to Proteins at High Binding Ratios. Possible Implications for the State of Proteins in Biological Membranes. Proc. Natl. Acad. Sci. USA 1970, 66, 1002–1007. [Google Scholar] [CrossRef] [Green Version]
- Madej, T.; Lanczycki, C.J.; Zhang, D.; Thiessen, P.A.; Geer, R.C.; Marchler-Bauer, A.; Bryant, S.H. MMDB and VAST+: Tracking structural similarities between macromolecular complexes. Nucleic Acids Res. 2014, 42, D297–D303. [Google Scholar] [CrossRef]
- Hosaka, M.; Nagahama, M.; Kim, W.S.; Watanabe, T.; Hatsuzawa, K.; Ikemizu, J.; Murakami, K.; Nakayama, K. Arg-X-Lys/Arg-Arg motif as a signal for precursor cleavage catalyzed by furin within the constitutive secretory pathway. J. Biol. Chem. 1991, 266, 12127–12130. [Google Scholar] [PubMed]
- Matos, J.L.; Fiori, C.S.; Silva-Filho, M.C.; Moura, D.S. A conserved dibasic site is essential for correct processing of the peptide hormone AtRALF1 in Arabidopsis thaliana. FEBS Lett. 2008, 582, 3343–3347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, K.; Pathak, D.C.; Saikia, D.P.; Debnath, A.; Ramakrishnan, S.; Dey, S.; Chellappa, M.M. Generation and evaluation of a recombinant Newcastle disease virus strain R2B with an altered fusion protein cleavage site as a vaccine candidate. Microb. Pathog. 2018, 118, 230–237. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of rhIL-12 are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Multimer (%) a | Monomer (%) a | Fragment (%) a | |||
|---|---|---|---|---|---|---|
| SDS-PAGE | SEC-HPLC | SDS-PAGE | SEC-HPLC | SDS-PAGE | SEC-HPLC | |
| S01 | 0.69 | 4.19 | 92.00 | 95.81 | 7.31 | –b |
| S02 | 0.71 | 2.96 | 91.32 | 97.04 | 7.97 | –b |
| S03 | 0.98 | 4.60 | 91.29 | 95.40 | 7.73 | –b |
| Subunit | Amino Acid Sequence | Theoretical MW (Da) | Measured MW (Da) | Error (Da) | Relative Error (ppm) |
|---|---|---|---|---|---|
| p40 | 1–260 | 29415.12 a | 29414.00 | 1.12 | 38 |
| 261–306 | 5300.93 | 5300.60 | 0.33 | 62 | |
| p35 | 1–197 | 22544.21 b | 22543.60 | 0.61 | 27 |
| 23200.80 c | 23199.00 | 1.80 | 78 | ||
| 23492.06 d | 23490.20 | 1.86 | 79 |
| Samples | Protein Content (mg/mL, Mean of Three Replicates) | Biological Activity (units/mL, Mean of Three Replicates) | Specific Activity (units/mg) |
|---|---|---|---|
| In-house reference | 1.78 | 1.57 × 107 | 8.81 × 106 |
| S01 | 0.35 | 1.56 × 106 | 4.45 × 106 |
| S02 | 0.34 | 1.46 × 106 | 4.31 × 106 |
| S03 | 0.35 | 1.57 × 106 | 4.48 × 106 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Li, Y.; Tao, L.; Jia, C.; Yao, W.; Rao, C.; Wang, J. Identification of a Recombinant Human Interleukin-12 (rhIL-12) Fragment in Non-Reduced SDS-PAGE. Molecules 2019, 24, 1210. https://doi.org/10.3390/molecules24071210
Yu L, Li Y, Tao L, Jia C, Yao W, Rao C, Wang J. Identification of a Recombinant Human Interleukin-12 (rhIL-12) Fragment in Non-Reduced SDS-PAGE. Molecules. 2019; 24(7):1210. https://doi.org/10.3390/molecules24071210
Chicago/Turabian StyleYu, Lei, Yonghong Li, Lei Tao, Chuncui Jia, Wenrong Yao, Chunming Rao, and Junzhi Wang. 2019. "Identification of a Recombinant Human Interleukin-12 (rhIL-12) Fragment in Non-Reduced SDS-PAGE" Molecules 24, no. 7: 1210. https://doi.org/10.3390/molecules24071210
APA StyleYu, L., Li, Y., Tao, L., Jia, C., Yao, W., Rao, C., & Wang, J. (2019). Identification of a Recombinant Human Interleukin-12 (rhIL-12) Fragment in Non-Reduced SDS-PAGE. Molecules, 24(7), 1210. https://doi.org/10.3390/molecules24071210