Natural Flavonoids as Potential Angiotensin-Converting Enzyme 2 Inhibitors for Anti-SARS-CoV-2

 ,
,  ,
,
Abstract
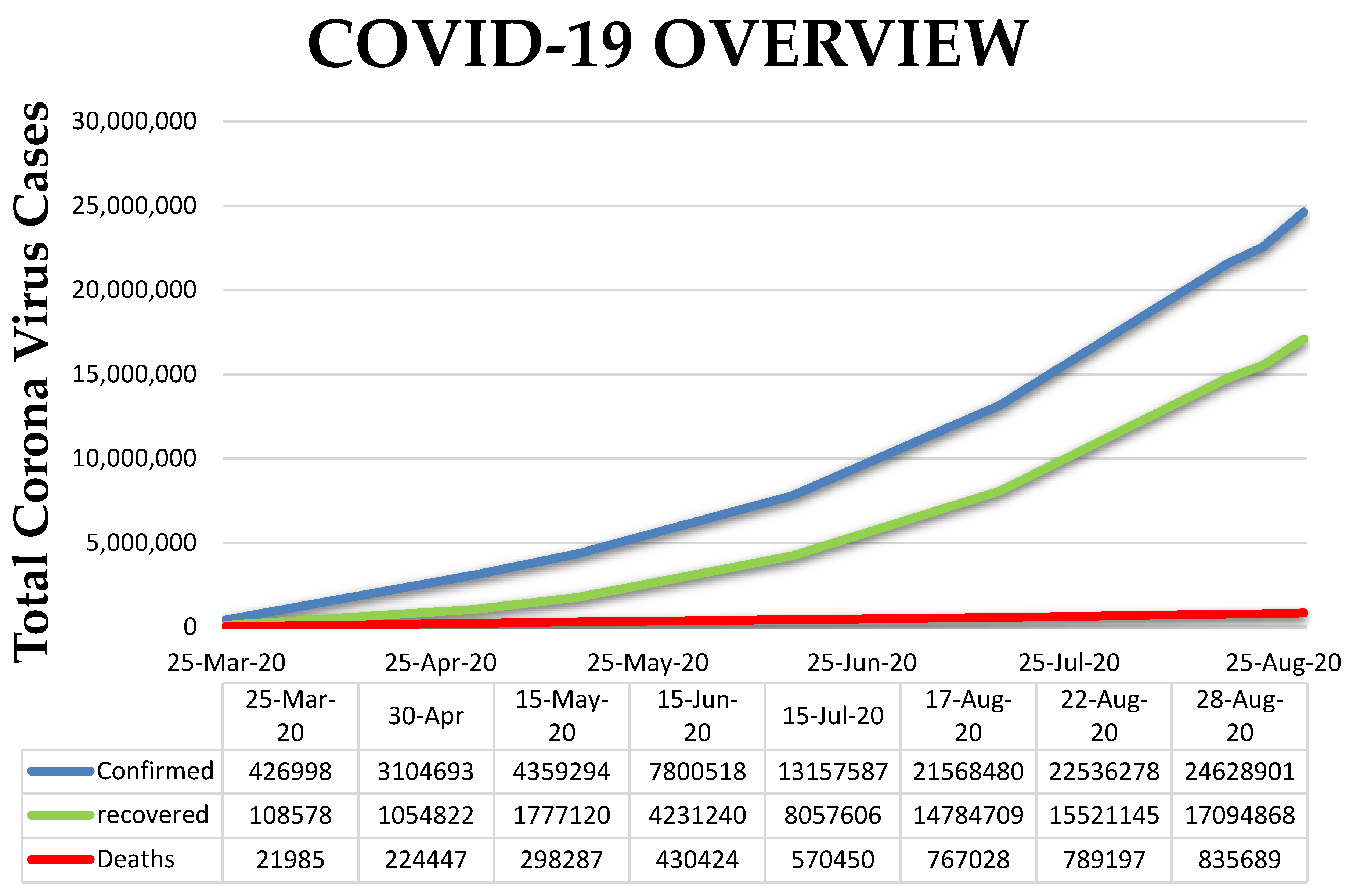
:1. Introduction
2. Methods
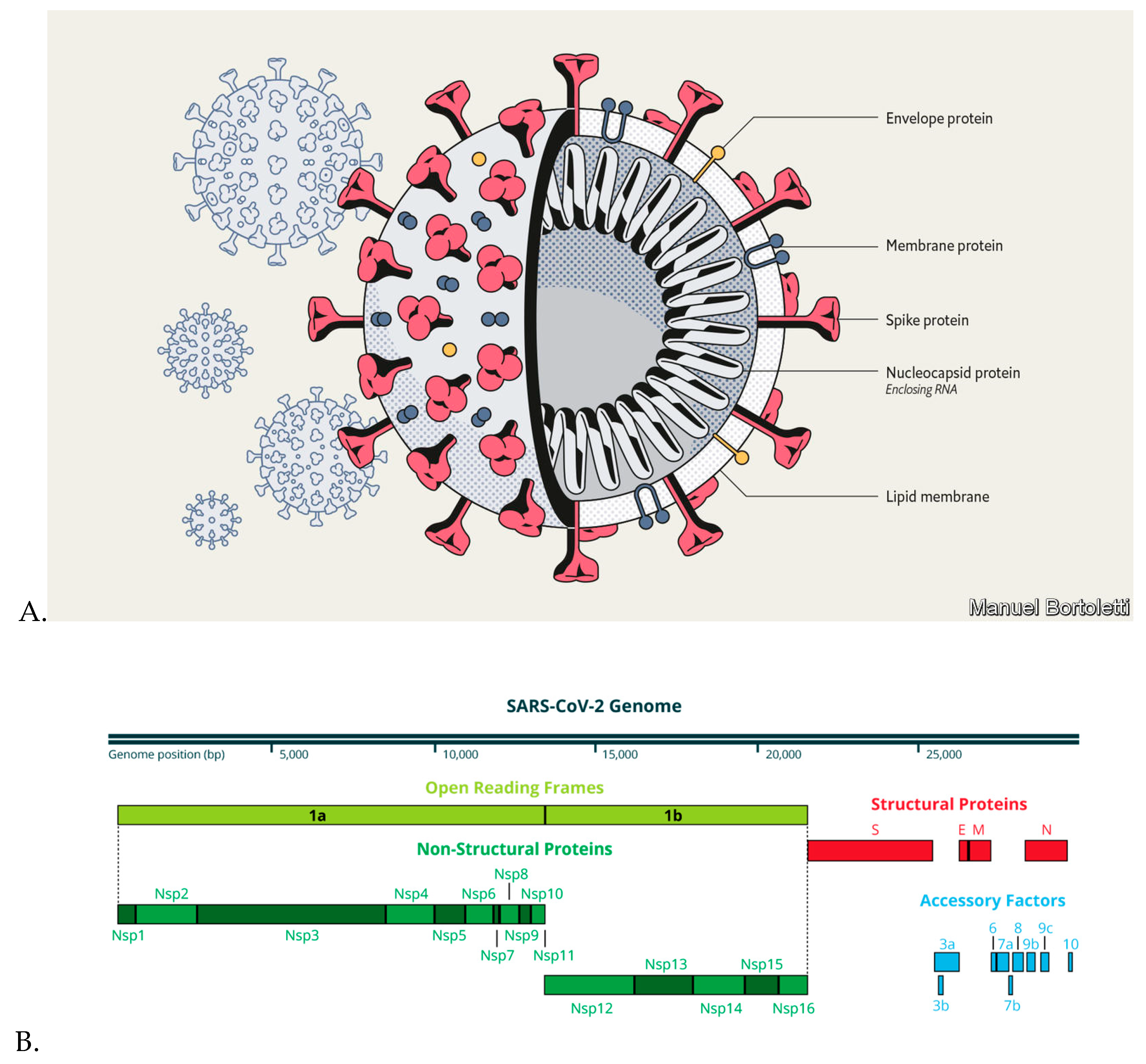
3. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2)
4. Angiotensin-Converting Enzyme 2 (ACE2)
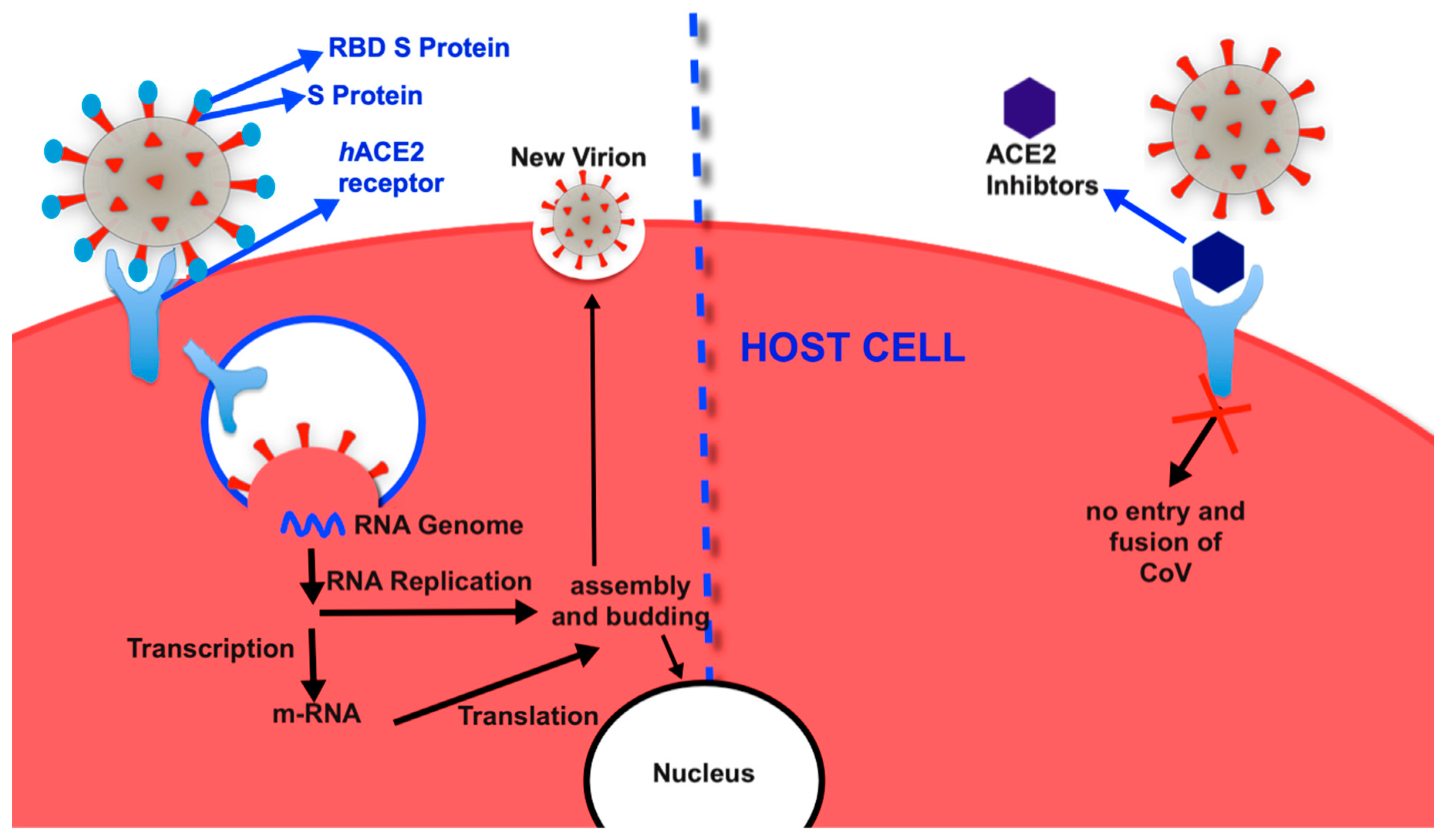
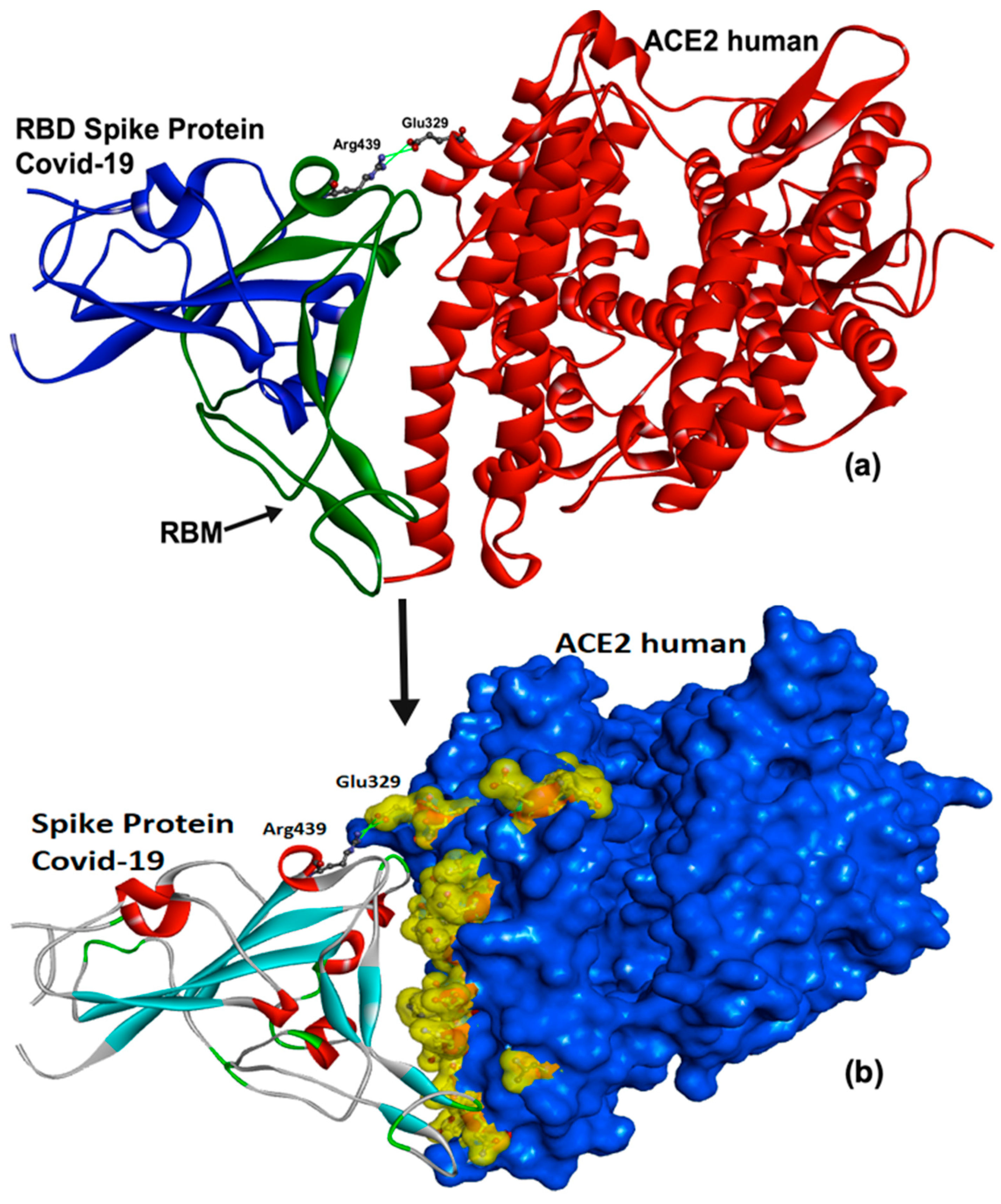
The Active Site of hACE2 as the Therapeutic Target of COVID-19
5. Inhibitors of ACE2
5.1. Synthetic Compounds of ACE2 Inhibitors
5.2. Natural Compounds Inhibiting ACE1 and ACE2 Receptors
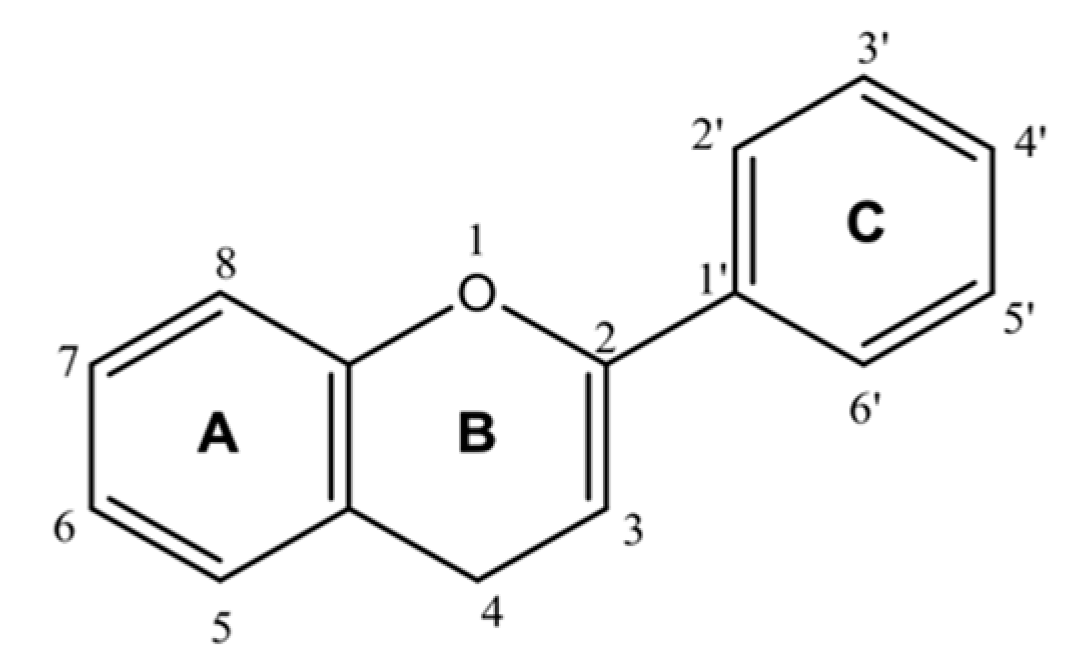
6. Flavonoids as ACE2 Inhibitors
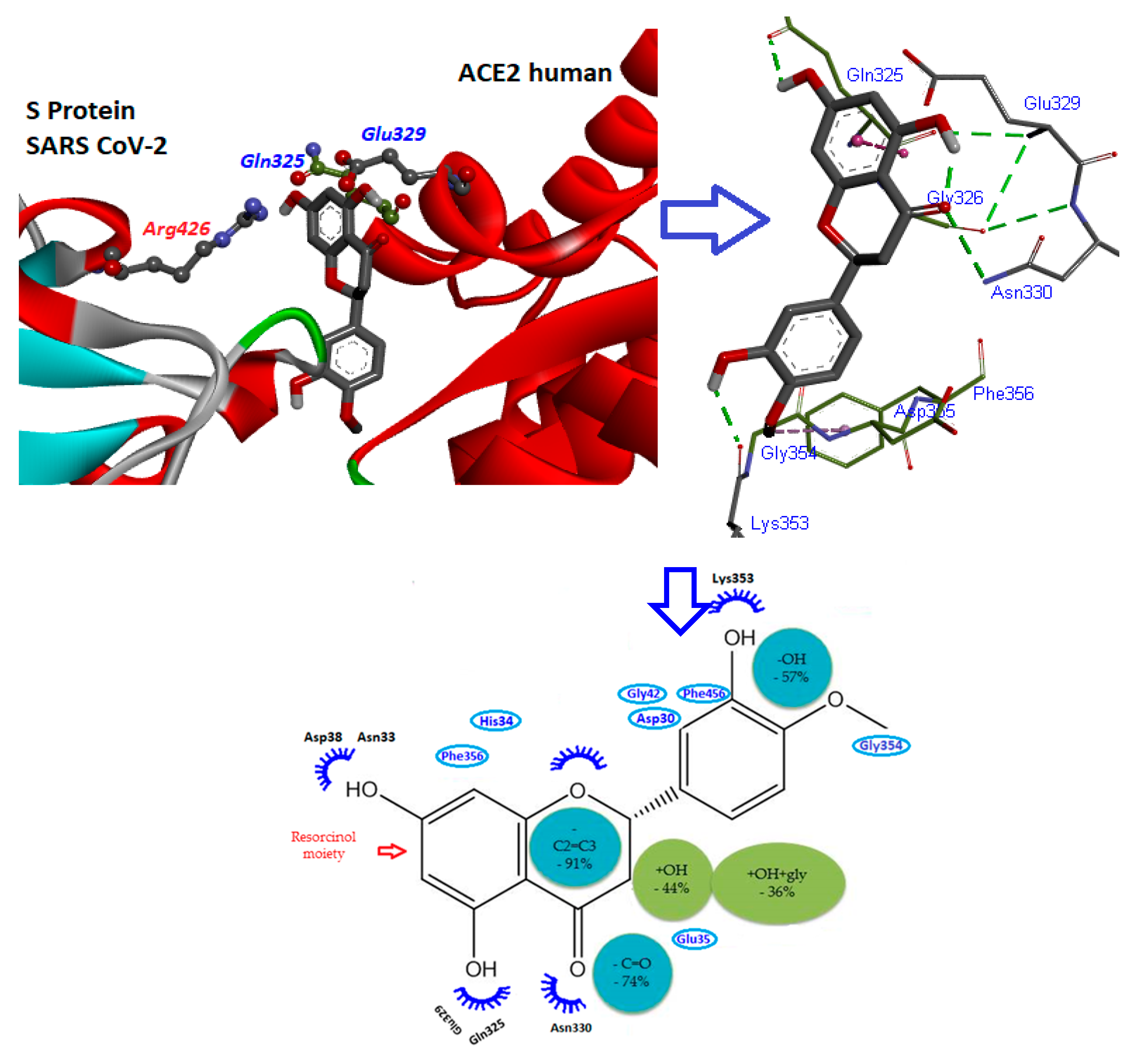
7. Mode of Action of Flavonoids
8. Perspectives and Overall Conclusion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Chhikara, B.S.; Rathi, B.; Singh, J.; Poonam, F. Corona virus SARS-CoV-2 disease COVID-19: Infection, prevention and clinical advances of the prospective chemical drug therapeutics. Chem. Biol. Lett. 2020, 7, 63–72. [Google Scholar]
- World Health Organization. Coronavirus Disease 2019 (COVID-19) Situation Report—70. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports/ (accessed on 2 May 2020).
- World Health Organization. WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19—11 March 2020. Available online: https://www.who.int/dg/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020 (accessed on 23 April 2020).
- Allcott, H.; Boxell, L.; Conway, J.; Gentzkow, M.; Thaler, M.; Yang, D.Y. Polarization and public health: Partisan differences in social distancing during the Coronavirus pandemic. NBER Work. Pap. 2020, 20, 1–52. [Google Scholar]
- Welt, F.G.; Shah, P.B.; Aronow, H.D.; Bortnick, A.E.; Henry, T.D.; Sherwood, M.W.; Young, M.N.; Davidson, L.J.; Kadavath, S.; Mahmud, E. Catheterization laboratory considerations during the coronavirus (COVID-19) pandemic: From ACC’s Interventional Council and SCAI. J. Am. Coll Cardiol. 2020, 75, 2372–2375. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Hao, X.; Lau, E.H.Y.; Wong, J.Y.; Leung, K.S.M.; Wu, J.T.; Cowling, B.J.; Leung, G.M. Real-time tentative assessment of the epidemiological characteristics of novel coronavirus infections in Wuhan, China, as at 22 January 2020. Eur. Commun. Dis. Bull. 2020, 25, 1–6. [Google Scholar] [CrossRef]
- Nadeem, S. Coronavirus COVID-19: Available free literature provided by various companies, journals and organizations around the world. J. Ongoing Chem. Res. 2020, 5, 7–13. [Google Scholar]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of coronavirus cell entry mediated by the viral spike protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Song, W.; Huang, H.; Sun, Q. Pharmacological therapeutics targeting RNA-dependent RNA polymerase, proteinase and spike protein: From mechanistic studies to clinical trials for COVID-19. J. Clin. Med. 2020, 9, 1131. [Google Scholar] [CrossRef] [Green Version]
- Chang, G.-G. Quaternary Structure of the SARS Coronavirus Main Protease. In Molecular Biology of the SARS-Coronavirus; Sunil, K.L., Ed.; Springer: New Delhi, India, 2009; Volume 1, pp. 115–128. [Google Scholar]
- Gheblawi, M.; Wang, K.; Viveiros, A.; Nguyen, Q.; Zhong, J.-C.; Turner, A.J.; Raizada, M.K.; Grant, M.B.; Oudit, G.Y. Angiotensin-converting enzyme 2: SARS-CoV-2 receptor and regulator of the renin-angiotensin system: Celebrating the 20th anniversary of the discovery of ACE2. Circ. Res. 2020, 126, 1456–1474. [Google Scholar] [CrossRef]
- Bahbah, E.I.; Negida, A.; Nabet, M.S. Purposing Saikosaponins for the treatment of COVID-19. Med. Hypoth. 2020, 140, 109782. [Google Scholar] [CrossRef]
- Jiang, S. Expert consensus on chloroquine phosphate for the treatment of novel coronavirus pneumonia (in Chinnese). Chin. J. Tuberc. Respir. Dis. 2020, 43, 185–188. [Google Scholar] [CrossRef]
- Gao, J.; Tian, Z.; Yang, X. Breakthrough: Chloroquine phosphate has shown apparent efficacy in treatment of COVID-19 associated pneumonia in clinical studies. Biosci. Trends 2020, 14, 72–73. [Google Scholar] [CrossRef] [Green Version]
- Gautret, P.; Lagier, J.-C.; Parola, P.; Hoang, V.T.; Meddeb, L.; Mailhe, M.; Doudier, B.; Courjon, J.; Giordanengo, V.; Vieira, V.E.; et al. Hydroxychloroquine and azithromycin as a treatment of COVID-19: Results of an open-label non-randomized clinical trial. Int. J. Antimicrob. Agents 2020, 56, 105949. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; LIU, D.; LIU, L.; LIU, P.; XU, Q.; XIA, L.; LING, Y.; HUANG, D.; SONG, S.; ZHANG, D. A pilot study of hydroxychloroquine in treatment of patients with common coronavirus disease-19 (COVID-19) (in Chinese). J. Zhejiang Univ. (Med. Sci.) 2020, 49, 215–219. [Google Scholar] [CrossRef]
- Cao, Y.-c.; Deng, Q.-x.; Dai, S.-x. Remdesivir for severe acute respiratory syndrome coronavirus 2 causing COVID-19: An evaluation of the evidence. Travel Med. Infect. Dis. 2020, 101647. [Google Scholar] [CrossRef] [PubMed]
- Grein, J.; Ohmagari, N.; Shin, D.; Diaz, G.; Asperges, E.; Castagna, A.; Feldt, T.; Green, G.; Green, M.L.; Lescure, F.-X. Compassionate use of remdesivir for patients with severe Covid-19. N. Engl. J. Med. 2020, 382, 2327–2336. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Yang, M.; Liu, D.; Chen, J.; Shu, D.; Xia, J.; Liao, X.; Gu, Y.; Cai, Q.; Yang, Y. Experimental treatment with favipiravir for COVID-19: An open-label control study. Engineering 2020, in press. [Google Scholar]
- Jean, S.-S.; Lee, P.-I.; Hsueh, P.-R. Treatment options for COVID-19: The reality and challenges. J. Microbiol. Immunol. Infect. 2020, 53, 436–443. [Google Scholar] [CrossRef]
- Rosa, S.G.V.; Santos, W.C. Clinical trials on drug repositioning for COVID-19 treatment. Rev. Panam Salud Publica 2020, 44, e40. [Google Scholar] [CrossRef]
- Carnero Contentti, E.; Correa, J. Immunosuppression during the COVID-19 pandemic in neuromyelitis optica spectrum disorders patients: A new challenge. Mult. Scler. Relat. Disord. 2020, 41, 102097. [Google Scholar] [CrossRef]
- Islam, M.T.; Sarkar, C.; El-Kersh, D.M.; Jamaddar, S.; Uddin, S.J.; Shilpi, J.A.; Mubarak, M.S. Natural products and their derivatives against coronavirus: A review of the non-clinical and pre-clinical data. Phytother. Res. 2020, 1–22. [Google Scholar] [CrossRef]
- Perrone, F.; Piccirillo, M.C.; Ascierto, P.A.; Salvarani, C.; Parrella, R.; Marata, A.M.; Popoli, P.; Ferraris, L.; Trischitta, M.M.M.; Ripamonti, D. Tocilizumab for patients with COVID-19 pneumonia. The TOCIVID-19 phase 2 trial. medRxiv 2020, 6, 1–24. [Google Scholar]
- Ascierto, P.A.; Fox, B.A.; Urba, W.J.; Anderson, A.C.; Atkins, M.B.; Borden, E.C.; Brahmer, J.R.; Butterfield, L.H.; Cesano, A.; Chen, D.S. Insights from immuno-oncology: The Society for Immunotherapy of Cancer Statement on access to IL-6-targeting therapies for COVID-19. J. Immunol. Ther. Cancer 2020, 8, e000878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, D.; Du, G.; Du, R.; Zhao, J.; Jin, Y.; Fu, S.; Gao, L.; Cheng, Z.; Lu, Q. Remdesivir in adults with severe COVID-19: A randomised, double-blind, placebo-controlled, multicentre trial. Lancet 2020, 395, 1569–1578. [Google Scholar] [CrossRef]
- Cavalcanti, A.B.; Zampieri, F.G.; Rosa, R.G.; Azevedo, L.C.P.; Veiga, V.C.; Avezum, A.; Damiani, L.P.; Marcadenti, A.; Kawano-Dourado, L.; Lisboa, T.; et al. Hydroxychloroquine with or without Azithromycin in Mild-to-Moderate Covid-19. N. Engl. J. Med. 2020, 1–12. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [Green Version]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, e125124. [Google Scholar] [CrossRef]
- Gorlach, S.; Fichna, J.; Lewandowska, U. Polyphenols as mitochondria-targeted anticancer drugs. Cancer Lett. 2015, 366, 141–149. [Google Scholar] [CrossRef]
- Brunetti, C.; Di Ferdinando, M.; Fini, A.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants and developmental regulators: Relative significance in plants and humans. Int. J. Mol. Sci. 2013, 14, 3540–3555. [Google Scholar] [CrossRef] [Green Version]
- Nabavi, S.F.; Braidy, N.; Habtemariam, S.; Orhan, I.E.; Daglia, M.; Manayi, A.; Gortzi, O.; Nabavi, S.M. Neuroprotective effects of chrysin: From chemistry to medicine. Neurochem. Int. 2015, 90, 224–231. [Google Scholar] [CrossRef]
- Moghrovyan, A.; Sahakyan, N.; Babayan, A.; Chichoyan, N.; Petrosyan, M.; Trchounian, A. Essential oil and ethanol extract of oregano (Origanum vulgare L.) from Armenian flora as a natural source of terpenes, flavonoids and other phytochemicals with antiradical, antioxidant, metal chelating, tyrosinase inhibitory and antibacterial activity. Curr. Pharm. Des. 2019, 25, 1809–1816. [Google Scholar] [CrossRef]
- Xie, Y.; Yang, W.; Tang, F.; Chen, X.; Ren, L. Antibacterial activities of flavonoids: Structure-activity relationship and mechanism. Curr. Med. Chem. 2015, 22, 132–149. [Google Scholar] [CrossRef] [PubMed]
- Kaul, T.N.; Middleton, E., Jr.; Ogra, P.L. Antiviral effect of flavonoids on human viruses. J. Med. Virol. 1985, 15, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Malik, Y.S.; Sircar, S.; Bhat, S.; Sharun, K.; Dhama, K.; Dadar, M.; Tiwari, R.; Chaicumpa, W. Emerging novel coronavirus (2019-nCoV)—Current scenario, evolutionary perspective based on genome analysis and recent developments. Vet. Q. 2020, 40, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J. SARS-CoV-2: An emerging coronavirus that causes a global threat. Int. J. Biol. Sci. 2020, 16, 1678. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Kulcsar, K.; Misra, V.; Frieman, M.; Mossman, K. Bats and coronaviruses. Viruses 2019, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Prompetchara, E.; Ketloy, C.; Palaga, T. Immune responses in COVID-19 and potential vaccines: Lessons learned from SARS and MERS epidemic. Asian Pac. J. Allergy Immunol. 2020, 38, 1–9. [Google Scholar]
- Yang, Y.; Xiao, Z.; Ye, K.; He, X.; Sun, B.; Qin, Z.; Yu, J.; Yao, J.; Wu, Q.; Bao, Z.; et al. SARS-CoV-2: Characteristics and current advances in research. Virol. J. 2020, 17, 117–134. [Google Scholar] [CrossRef]
- Lai, M.M. The making of infectious viral RNA: No size limit in sight. Proc. Natl. Acad. Sci. USA 2000, 97, 5025–5027. [Google Scholar] [CrossRef] [Green Version]
- Phan, M.V.T.; Ngo Tri, T.; Hong Anh, P.; Baker, S.; Kellam, P.; Cotten, M. Identification and characterization of Coronaviridae genomes from Vietnamese bats and rats based on conserved protein domains. Virus Evol. 2018, 4, vey035. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, X.; Li, T.; Zhang, S.; Wang, L.; Wu, X.; Liu, J. The genetic sequence, origin, and diagnosis of SARS-CoV-2. Eur. J. Clin. Microbio. Infect. Dis. 2020, 39, 1629–1635. [Google Scholar] [CrossRef]
- Chan, J.F.-W.; Kok, K.-H.; Zhu, Z.; Chu, H.; To, K.K.-W.; Yuan, S.; Yuen, K.-Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abduljalil, J.M.; Abduljalil, B.M. Epidemiology, genome, and clinical features of the pandemic SARS-CoV-2: A recent view. New Microbes New Infect. 2020, 35, 100672–100680. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Lorusso, A. Novel human coronavirus (SARS-CoV-2): A lesson from animal coronaviruses. Vet. Microb. 2020, 108693. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.P.; Look, D.C.; Shi, L.; Hickey, M.; Pewe, L.; Netland, J.; Farzan, M.; Wohlford-Lenane, C.; Perlman, S.; McCray, P.B. ACE2 receptor expression and severe acute respiratory syndrome coronavirus infection depend on differentiation of human airway epithelia. J. Virol. 2005, 79, 14614–14621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.F.-W.; Zhang, A.J.; Yuan, S.; Poon, V.K.-M.; Chan, C.C.-S.; Lee, A.C.-Y.; Chan, W.-M.; Fan, Z.; Tsoi, H.-W.; Wen, L. Simulation of the clinical and pathological manifestations of Coronavirus Disease 2019 (COVID-19) in golden Syrian hamster model: Implications for disease pathogenesis and transmissibility. Clin. Infect. Dis. 2020, 325, 1–55. [Google Scholar] [CrossRef] [Green Version]
- Bavishi, C.; Maddox, T.M.; Messerli, F.H. Coronavirus Disease 2019 (COVID-19) Infection and Renin Angiotensin System Blockers. JAMA Cardiol. 2020, 5, 745–747. [Google Scholar] [CrossRef] [Green Version]
- Wysocki, J.; Garcia-Halpin, L.; Ye, M.; Maier, C.; Sowers, K.; Burns, K.D.; Batlle, D. Regulation of urinary ACE2 in diabetic mice. Am. J. Physiol.-Ren. Physiol. 2013, 305, F600–F611. [Google Scholar] [CrossRef] [Green Version]
- Xiao, F.; Hiremath, S.; Knoll, G.; Zimpelmann, J.; Srivaratharajah, K.; Jadhav, D.; Fergusson, D.; Kennedy, C.R.; Burns, K.D. Increased urinary angiotensin-converting enzyme 2 in renal transplant patients with diabetes. PLoS ONE 2012, 7, e37649. [Google Scholar] [CrossRef] [Green Version]
- Becker, L.K.; Totou, N.L.; Oliveira, M.F.; Coelho, D.B.; de Oliveira, E.C.; Motta-Santos, D.; Silami-Garcia, E.; Campagnole-Santos, M.J.; Santos, R.A.S. Lifetime overproduction of circulating angiotensin-(1-7) in rats attenuates the increase in skeletal muscle damage biomarkers after exhaustive exercise. Chin. J. Physiol. 2019, 62, 226. [Google Scholar]
- Zeinalian, M.; Salari-Jazi, A.; Jannesari, A.; Khanahmad, H. A potential protective role of losartan against coronavirus-induced lung damage. Infect. Control Hosp. Epidemiol. 2020, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, C.; Kaushal, S.; Yeo, D. Enteric involvement of coronaviruses: Is faecal–oral transmission of SARS-CoV-2 possible? Lancet Gastroenterol. Hepatol. 2020, 5, 335–337. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Yu, J.; Wang, K.; Chen, Z.; Wen, C.; Xu, Z. The spatial and cell-type distribution of SARS-CoV-2 receptor ACE2 in human and mouse brain. BioRxiv 2020, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 182, 271–280. [Google Scholar] [CrossRef]
- Imai, Y.; Kuba, K.; Penninger, J.M. The discovery of angiotensin-converting enzyme 2 and its role in acute lung injury in mice. Exp. Physiol. 2008, 93, 543–548. [Google Scholar] [CrossRef]
- Othman, H.; Bouslama, Z.; Brandenburg, J.-T.; Da Rocha, J.; Hamdi, Y.; Ghedira, K.; Abid, N.-S.; Hazelhurst, S. In silico study of the spike protein from SARS-CoV-2 interaction with ACE2: Similarity with SARS-CoV, hot-spot analysis and effect of the receptor polymorphism. BioRxiv 2020, 527, 702–708. [Google Scholar] [CrossRef]
- Xu, Y.; Lou, Z.; Liu, Y.; Pang, H.; Tien, P.; Gao, G.F.; Rao, Z. Crystal structure of severe acute respiratory syndrome coronavirus spike protein fusion core. J. Biol. Chem. 2004, 279, 49414–49419. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Zhu, Y.; Liu, M.; Lan, Q.; Xu, W.; Wu, Y.; Ying, T.; Liu, S.; Shi, Z.; Jiang, S.; et al. Fusion mechanism of 2019-nCoV and fusion inhibitors targeting HR1 domain in spike protein. Cell Mol. Immunol. 2020, 17, 765–767. [Google Scholar] [CrossRef]
- Su, Q.-d.; Yi, Y.; Zou, Y.-n.; Jia, Z.-y.; Qiu, F.; Wang, F.; Yin, W.-j.; Zhou, W.-t.; Zhang, S.; Yu, P.-c.; et al. The biological characteristics of SARS-CoV-2 Spike protein Pro330-Leu650. Vaccine 2020, 38, 5071–5075. [Google Scholar] [CrossRef]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, A.; Sarkar, A.; Maulik, U. Computational approach for the design of potential spike protein binding natural compounds in SARS-CoV2. OSF Preprints 2020, 1–22. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, S.; Teng, T.; Abdalla, A.E.; Zhu, W.; Xie, L.; Wang, Y.; Guo, X. Systematic Comparison of Two Animal-to-Human Transmitted Human Coronaviruses: SARS-CoV-2 and SARS-CoV. Viruses 2020, 12, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, D.L.; Sternberg, A.; Stange, U.; Laufer, S.; Naujokat, C. Candidate drugs against SARS-CoV-2 and COVID-19. Pharmacol. Res. 2020, 157, 104859. [Google Scholar] [CrossRef] [PubMed]
- Magrone, T.; Magrone, M.; Jirillo, E. Focus on Receptors for Coronaviruses with Special Reference to Angiotensin-converting Enzyme 2 as a Potential Drug Target—A Perspective. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 807–811. [Google Scholar] [CrossRef]
- Samavati, L.; Uhal, B.D. ACE2, Much More Than Just a Receptor for SARS-COV-2. Front. Cell. Infect. Microbiol. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Han, D.P.; Penn-Nicholson, A.; Cho, M.W. Identification of critical determinants on ACE2 for SARS-CoV entry and development of a potent entry inhibitor. Virology 2006, 350, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.-w.; Wang, L.; Bao, L.-d. Exploring the Active Compounds of Traditional Mongolian Medicine in Intervention of Novel Coronavirus (COVID-19) Based on Molecular Docking Method. J. Func. Foods 2020, 71, 104016. [Google Scholar] [CrossRef]
- Zhai, X.; Sun, J.; Yan, Z.; Zhang, J.; Zhao, J.; Zhao, Z.; Gao, Q.; He, W.T.; Veit, M.; Su, S. Comparison of SARS-CoV-2 spike protein binding to ACE2 receptors from human, pets, farm animals, and putative intermediate hosts. J. Virol. 2020, 94, 1–16. [Google Scholar] [CrossRef]
- Guy, J.L.; Jackson, R.M.; Acharya, K.R.; Sturrock, E.D.; Hooper, N.M.; Turner, A.J. Angiotensin-converting enzyme-2 (ACE2): Comparative modeling of the active site, specificity requirements, and chloride dependence. Biochemistry 2003, 42, 13185–13192. [Google Scholar] [CrossRef]
- Kuster, G.M.; Pfister, O.; Burkard, T.; Zhou, Q.; Twerenbold, R.; Haaf, P.; Widmer, A.F.; Osswald, S. SARS-CoV2: Should inhibitors of the renin–angiotensin system be withdrawn in patients with COVID-19? Eur. Heart J. 2020, 41, 1801–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Zhu, L.; Cai, J.; Lei, F.; Qin, J.-J.; Xie, J.; Liu, Y.-M.; Zhao, Y.-C.; Huang, X.; Lin, L. Association of inpatient use of angiotensin converting enzyme inhibitors and angiotensin II receptor blockers with mortality among patients with hypertension hospitalized with COVID-19. Circ. Res. 2020, 126, 1671–1681. [Google Scholar] [CrossRef] [PubMed]
- Schumacher, H.; Mancia, G. The safety profile of telmisartan as monotherapy or combined with hydrochlorothiazide: A retrospective analysis of 50 studies. Blood Press. 2008, 17, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Rothlin, R.P.; Vetulli, H.M.; Duarte, M.; Pelorosso, F.G. Telmisartan as tentative angiotensin receptor blocker therapeutic for COVID-19. Drug Dev. Res. 2020, 1–3. [Google Scholar] [CrossRef]
- Trask, A.J.; Groban, L.; Westwood, B.M.; Varagic, J.; Ganten, D.; Gallagher, P.E.; Chappell, M.C.; Ferrario, C.M. Inhibition of Angiotensin-Converting Enzyme 2 Exacerbates Cardiac Hypertrophy and Fibrosis in Ren-2 Hypertensive Rats. Am. J. Hypert. 2010, 23, 687–693. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.; Balasubramanian, N.; Vasam, G.; Jarajapu, Y.P. Angiotensin converting enzyme versus angiotensin converting enzyme-2 selectivity of MLN-4760 and DX600 in human and murine bone marrow-derived cells. Eur. J. Pharmacol. 2016, 774, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Towler, P.; Staker, B.; Prasad, S.G.; Menon, S.; Tang, J.; Parsons, T.; Ryan, D.; Fisher, M.; Williams, D.; Dales, N.A.; et al. ACE2 X-ray structures reveal a large hinge-bending motion important for inhibitor binding and catalysis. J. Biol. Chem. 2004, 279, 17996–18007. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Yu, X.; Cheng, L.; Miao, L.Y.; Li, H.X.; Han, L.H.; Jiang, W.P. Effects of enalapril on the expression of cardiac angiotensin-converting enzyme and angiotensin-converting enzyme 2 in spontaneously hypertensive rats. Arch. Cardiovasc. Dis. 2013, 106, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, K.; Zimmerman, M.A.; Sullivan, J.C. Sex Differences in Angiotensin-Converting Enzyme Modulation of Ang (1–7) Levels in Normotensive WKY Rats. Am. J. Hypertens. 2013, 26, 591–598. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.; Wysocki, J.; Gonzalez-Pacheco, F.R.; Salem, M.; Evora, K.; Garcia-Halpin, L.; Poglitsch, M.; Schuster, M.; Batlle, D. Murine recombinant angiotensin-converting enzyme 2: Effect on angiotensin II-dependent hypertension and distinctive angiotensin-converting enzyme 2 inhibitor characteristics on rodent and human angiotensin-converting enzyme 2. Hypertension 2012, 60, 730–740. [Google Scholar] [CrossRef] [Green Version]
- Haga, S.; Nagata, N.; Okamura, T.; Yamamoto, N.; Sata, T.; Yamamoto, N.; Sasazuki, T.; Ishizaka, Y. TACE antagonists blocking ACE2 shedding caused by the spike protein of SARS-CoV are candidate antiviral compounds. Antivir. Res. 2010, 85, 551–555. [Google Scholar] [CrossRef]
- Furuhashi, M.; Moniwa, N.; Mita, T.; Fuseya, T.; Ishimura, S.; Ohno, K.; Shibata, S.; Tanaka, M.; Watanabe, Y.; Akasaka, H.; et al. Urinary angiotensin-converting enzyme 2 in hypertensive patients may be increased by olmesartan, an angiotensin II receptor blocker. Am. J. Hypertens. 2015, 28, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malek, V.; Sharma, N.; Sankrityayan, H.; Gaikwad, A.B. Concurrent neprilysin inhibition and renin-angiotensin system modulations prevented diabetic nephropathy. Life Sci. 2019, 221, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Vincent, M.J.; Bergeron, E.; Benjannet, S.; Erickson, B.R.; Rollin, P.E.; Ksiazek, T.G.; Seidah, N.G.; Nichol, S.T. Chloroquine is a potent inhibitor of SARS coronavirus infection and spread. Virol. J. 2005, 2, 69. [Google Scholar] [CrossRef] [Green Version]
- Goru, S.K.; Kadakol, A.; Malek, V.; Pandey, A.; Sharma, N.; Gaikwad, A.B. Diminazene aceturate prevents nephropathy by increasing glomerular ACE2 and AT2 receptor expression in a rat model of type1 diabetes. Br. J. Pharmacol. 2017, 174, 3118–3130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huentelman, M.J.; Zubcevic, J.; Prada, J.A.H.; Xiao, X.; Dimitrov, D.S.; Raizada, M.K.; Ostrov, D.A. Structure-Based Discovery of a Novel Angiotensin-Converting Enzyme 2 Inhibitor. Hypertension 2004, 44, 903–906. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.L.; Yang, J.M.; Sun, Y.P.; Su, G.H. Inhibitors of RAS Might Be a Good Choice for the Therapy of COVID-19 Pneumonia. Chin. J. Tuberc. Respir. Dis. 2020, 43, E014. [Google Scholar] [CrossRef]
- Xie, X.; Chen, J.; Wang, X.; Zhang, F.; Liu, Y. Age- and gender-related difference of ACE2 expression in rat lung. Life Sci. 2006, 78, 2166–2171. [Google Scholar] [CrossRef] [PubMed]
- Gullo, V.P.; McAlpine, J.; Lam, K.S.; Baker, D.; Petersen, F. Drug discovery from natural products. J. Ind. Microbiol. Biotechnol. 2006, 33, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z. The modification of natural products for medical use. Acta Pharm. Sin. B 2017, 7, 119–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Ganjhu, R.K.; Mudgal, P.P.; Maity, H.; Dowarha, D.; Devadiga, S.; Nag, S.; Arunkumar, G. Herbal plants and plant preparations as remedial approach for viral diseases. Virusdisease 2015, 26, 225–236. [Google Scholar] [CrossRef]
- Utomo, R.Y.; Ikawati, M.; Meiyanto, E. Revealing the Potency of Citrus and Galangal Constituents to Halt SARS-CoV-2 Infection. Preprints 2020, 214, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Loizzo, M.R.; Said, A.; Tundis, R.; Rashed, K.; Statti, G.A.; Hufner, A.; Menichini, F. Inhibition of angiotensin converting enzyme (ACE) by flavonoids isolated from Ailanthus excelsa (Roxb) (Simaroubaceae). Phytother. Res. 2007, 21, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.-Y.; Wu, S.-L.; Chen, J.-C.; Li, C.-C.; Hsiang, C.-Y. Emodin blocks the SARS coronavirus spike protein and angiotensin-converting enzyme 2 interaction. Antivir. Res. 2007, 74, 92–101. [Google Scholar] [CrossRef]
- Ojeda, D.; Jimenez-Ferrer, E.; Zamilpa, A.; Herrera-Arellano, A.; Tortoriello, J.; Alvarez, L. Inhibition of angiotensin convertin enzyme (ACE) activity by the anthocyanins delphinidin- and cyanidin-3-O-sambubiosides from Hibiscus sabdariffa. J. Ethnopharmacol. 2010, 127, 7–10. [Google Scholar] [CrossRef]
- Sui, H.; Yu, Q.; Zhi, Y.; Geng, G.; Liu, H.; Xu, H. Effects of apigenin on the expression of angiotensin-converting enzyme 2 in kidney in spontaneously hypertensive rats. J. Hyg. Res. 2010, 39, 693–696. [Google Scholar]
- Guerrero, L.; Castillo, J.; Quinones, M.; Garcia-Vallve, S.; Arola, L.; Pujadas, G.; Muguerza, B. Inhibition of angiotensin-converting enzyme activity by flavonoids: Structure-activity relationship studies. PLoS ONE 2012, 7, e49493. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, S.; Yoshiya, T.; Yoshizawa-Kumagaye, K.; Sugiyama, T. Nicotianamine is a novel angiotensin-converting enzyme 2 inhibitor in soybean. Biomed. Res. (Tokyo Japan) 2015, 36, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Hettihewa, S.K.; Hemar, Y.; Rupasinghe, H.P.V. Flavonoid-Rich Extract of Actinidia macrosperma (A Wild Kiwifruit) Inhibits Angiotensin-Converting Enzyme In Vitro. Foods 2018, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, R.S.; Jagdale, S.S.; Bansode, S.B.; Shankar, S.S.; Tellis, M.B.; Pandya, V.K.; Chugh, A.; Giri, A.P.; Kulkarni, M.J. Discovery of Potential Multi-Target-Directed Ligands by Targeting Host-specific SARS-CoV-2 Structurally Conserved Main Protease($). J. Biomol. Struct. Dyn. 2020, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.F.; Aluko, R.E.; Jin, Q.; Zhang, Y.; Yuan, L.J. Inhibitory activities of baicalin against renin and angiotensin-converting enzyme. Pharm. Biol. 2012, 50, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Du, Q. Potential natural compounds for preventing 2019-nCoV infection. Preprints 2020, 358, 1–17. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [Green Version]
- Durairaj, R.B. Resorcinol Structure and Physical Properties. Resorcinol. Chem. Technol. Appl. 2005, 1–9. [Google Scholar]
- Bhowmik, D.; Nandi, R.; Kumar, D. Evaluation of Flavonoids as 2019-nCoV Cell Entry Inhibitor Through Molecular Docking and Pharmacological Analysis. ChemRxiv 2020, 6, 1–27. [Google Scholar] [CrossRef]
- Yu, M.S.; Lee, J.; Lee, J.M.; Kim, Y.; Chin, Y.W.; Jee, J.G.; Keum, Y.S.; Jeong, Y.J. Identification of myricetin and scutellarein as novel chemical inhibitors of the SARS coronavirus helicase, nsP13. Bioorg. Med. Chem. Lett. 2012, 22, 4049–4054. [Google Scholar] [CrossRef]
- Schwarz, S.; Sauter, D.; Wang, K.; Zhang, R.; Sun, B.; Karioti, A.; Bilia, A.R.; Efferth, T.; Schwarz, W. Kaempferol derivatives as antiviral drugs against the 3a channel protein of coronavirus. Planta Med. 2014, 80, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Yi, L.; Li, Z.; Yuan, K.; Qu, X.; Chen, J.; Wang, G.; Zhang, H.; Luo, H.; Zhu, L.; Jiang, P.; et al. Small molecules blocking the entry of severe acute respiratory syndrome coronavirus into host cells. J. Virol. 2004, 78, 11334–11339. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.H.; Woo, H.-J.; Kang, H.-K.; Nguyen, V.D.; Kim, Y.-M.; Kim, D.-W.; Ahn, S.-A.; Xia, Y.; Kim, D. Flavonoid-mediated inhibition of SARS coronavirus 3C-like protease expressed in Pichia pastoris. Biotechnol. Lett. 2012, 34, 831–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, S.; Kim, H.; Kim, S.; Shin, D.H.; Kim, M.-S. Characteristics of flavonoids as potent MERS-CoV 3C-like protease inhibitors. Chem. Biol. Drug Des. 2019, 94, 2023–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-Y.; Yuk, H.J.; Ryu, H.W.; Lim, S.H.; Kim, K.S.; Park, K.H.; Ryu, Y.B.; Lee, W.S. Evaluation of polyphenols from Broussonetia papyrifera as coronavirus protease inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm. Sin. B 2020. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Bi, J.; Li, F.; Wang, S.; Huang, X.; Meng, X.; Sun, B.; Wang, D.; Kong, W.; Jiang, C. Antiviral Efficacy of Flavonoids against Enterovirus 71 Infection in Vitro and in Newborn Mice. Viruses 2019, 11, 625. [Google Scholar] [CrossRef] [Green Version]
- Zakaryan, H.; Arabyan, E.; Oo, A.; Zandi, K. Flavonoids: Promising natural compounds against viral infections. Arch. Virol. 2017, 162, 2539–2551. [Google Scholar] [CrossRef]
- Cheng, L.; Zheng, W.; Li, M.; Huang, J.; Bao, S.; Xu, Q.; Ma, Z. Citrus fruits are rich in flavonoids for immunoregulation and potential targeting ACE2. Preprints 2020, 2020020313. [Google Scholar]
- Kim, V.Y.; Batty, A.; Li, J.; Kirk, S.G.; Crowell, S.A.; Jin, Y.; Tang, J.; Zhang, J.; Rogers, L.K.; Deng, H.-X. Glutathione Reductase Promotes Fungal Clearance and Suppresses Inflammation during Systemic Candida albicans Infection in Mice. J. Immunol. 2019, 203, 2239–2251. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Brewster, R.M.; Manthey, J.A. Flavonoid Compositions and Uses Thereof. Google Patents WO2015152648A1, 8 October 2015. [Google Scholar]
- Wang, T.-Y.; Li, Q.; Bi, K.-S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Muchtaridi, M.; Sugijanto, M.; Mohd Gazzali, A.; Wahab, H.A. Anti-Neuraminidase Bioactives from Manggis Hutan (Garcinia celebica L.) Leaves: Partial Purification and Molecular Characterization. Molecules 2020, 25, 821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hošek, J.; Šmejkal, K. Flavonoids as Anti-inflammatory Agents. In Encyclopedia of Inflammatory Diseases; Parnham, M., Ed.; Springer: Basel, Switzerland, 2015; pp. 1–17. [Google Scholar]
- Treml, J.; Šmejkal, K. Flavonoids as Potent Scavengers of Hydroxyl Radicals. Compr. Rev. Food Sci. Food Saf. 2016, 15, 720–738. [Google Scholar] [CrossRef]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Chelombitko, M.A. Role of Reactive Oxygen Species in Inflammation: A Minireview. Mosc. Univ. Biol. Sci. Bull. 2018, 73, 199–202. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.-P.-P.; Sulaiman Rahman, H. Antioxidant and Oxidative Stress: A Mutual Interplay in Age-Related Diseases. Front. Pharmacol. 2018, 9, 1162–1190. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Rahaman, S. Flavonoids: A vital resource in healthcare and medicine. Pharm. Pharmacol. Int. J. 2020, 8, 91–104. [Google Scholar]
- Codorniu-Hernández, E.; Rolo-Naranjo, A.; Montero-Cabrera, L.A. Theoretical affinity order among flavonoids and amino acid residues: An approach to understand flavonoid–protein interactions. J. Mol. Struct. THEOCHEM 2007, 819, 121–129. [Google Scholar] [CrossRef]
- Tikellis, C.; Thomas, M.C. Angiotensin-Converting Enzyme 2 (ACE2) Is a Key Modulator of the Renin Angiotensin System in Health and Disease. Int. J. Pept. 2012, 2012, 256294. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Inhibitors | Derivates | Plants | Methods | Years | Source |
|---|---|---|---|---|---|---|
| 1. | Luteolin | Flavonoid | Ailanthus excelsa | In vitro using ACE2 via Elbl and Wagner methods | 2007 | [99] |
| 2. | Kaempferol | |||||
| 3. | Apigenin | |||||
| 4. | Quercetin | |||||
| 5. | Luteolin | |||||
| 6. | Emodin | Anthraquinone | Rheum officinale Polygonum multiflorum | In vitro using ACE2 | 2007 | [100] |
| 7. | Chrysin | Flavonoid | ||||
| 8. | Rhein | Flavonoid | ||||
| 9. | Delphinidin | Flavonoid | Hibiscus sabdariffa | In vitro ACE Inhibition assay | 2010 | [101] |
| 10. | Cyanidin | Flavonoid | ||||
| 11. | Apigenin | Flavonoid | Apium graveolens | In vitro using ACE2 isolated from kidney | 2010 | [102] |
| 12. | Rhoifolin | Flavonoid | Rhus succedanea | ACE activity was measured by a fluorometric assay | 2012 | [103] |
| 13. | Rutin and Quercetine | Fagopyrum tataricum | ||||
| 14. | Nicotianamine | Peptide | Glycine max | In vitro using internally quenched fluorogenic (IQF) substrate for ACE2 | 2015 | [104] |
| 15. | Quercetin | Flavonoid | Actinidia macrosperma | In vitro using a fluorescence-based biochemical assay against ACE enzyme | 2018 | [103,105] |
| 16. | Catechin | |||||
| 17. | Quercetin | |||||
| 18. | Epigallocatechin | |||||
| 19. | Epigallocatechin gallate | |||||
| 20. | Ferulic acid | Phenolic acid | ||||
| 21. | Chlorogenic acid | |||||
| 22. | Isoferulic acid | |||||
| 23. | Caffeic acid | |||||
| 24. | δ-Viniferin | Flavonoid | Vitis vinifera | Virtual screening against ACE2 using Autodock Vina | 2020 | [106] |
| 25. | Myritilin | Flavonoid | ||||
| 26. | Myricitrin | Flavonoid | ||||
| 27. | Taiwanhomoflavone A | Flavonoid | Cephalotaxus wilsoniana | |||
| 28. | Lactucopicrin 15-oxalate | Sesquiterpene lactone | Lactuca virosa | |||
| 29. | Nympholide A | Flavonoid | Nymphaea lotus | |||
| 30. | Afzelin | Flavonoid | Cornus macrophylla | |||
| 31. | Biorobin | Flavonoid | Acalypha indica | |||
| 32. | Phyllaemblicin B | sesquiterpenoid | Phyllanthus emblica | |||
| 33. | Baicalin | Flavonoid | Scutellaria baicalensis | Using spectroscopy method to determine renin and ACE activities | [107] | |
| 34. | Hesperetin | Flavonoid | Citrus aurantium | Virtual Screening against ACE2 using molecular docking | 2020 | [108] |
| 35. | Baicalin | Flavonoid | Scutellaria baicalensis | |||
| 36. | Scutellarin | Flavonoid | ||||
| 37. | Glycyrrhizin | Sesquiterpene | Glycyrrhiza radix | |||
| 38. | Curcumin | Curcuminoids | Curcuma xanthoriza | Virtual Screening against ACE2 using MOE molecular docking | 2020 | [98] |
| 39. | Tangeretin | Flavonoid | Citrus aurantifolia | |||
| 40. | Nobiletin | |||||
| 41. | Naringenin | |||||
| 42. | Brazilein | Flavonoid | Caesalpinia sappan | |||
| 43. | Brazilin | |||||
| 44. | Galangin | Flavonoid | Alpinia galanga | |||
| 45. | Acetoxychavicol acetate (ACA) | ACA derivatives |
| Name | Inhibition Approach | Effective Compound | Inhibition Potential (IC50/EC50) * | ADME | Reference |
|---|---|---|---|---|---|
| Rheum officinale (rhubarb) | Viral spike protein and human ACE2 receptor inhibitor | Emodin | 1–10 µM/mL | HIA: 85.74 Caco2: 20.30 PPB: 88.75 BBB: 0.37 | [119] |
| Reynoutria multiflora tuber | Viral spike protein and human ACE2 receptor inhibitor | Emodin | 1–10 µM/mL | HIA: 85.74 Caco2: 20.30 PPB: 88.75 BBB: 0.37 | [119] |
| Citrus accumulate | Viral spike protein and human ACE2 receptor inhibitor | Naringenin | Not yet reported | HIA: 87.31 Caco2: 10.52 PPB: 100 BBB: 0.59 | [100] |
| Citrus aurantium and Citri Reticulatae Pericarpium | Viral spike protein and human ACE2 receptor inhibitor | Hesperetin | Not yet reported | HIA: 87.19 Caco2: 7.003 PPB: 96.79 BBB: 0.22 | [121] |
| Scutellaria baicalensis Georgi | Viral spike protein and human ACE2 receptor inhibitor | Baicalin | 2.24 mM | HIA: 32.42 Caco2: 11.55 PPB: 75.69 BBB: 0.02 | [108] |
| Citrus | Viral spike protein and human ACE2 receptor inhibitor | Neohesperidin | Not yet reported | HIA: 8.80 Caco2: 7.07 PPB: 44.05 BBB: 0.02 | [100] |
| Citrus | Viral spike protein and human ACE2 receptor inhibitor | Nobiletin | Not yet reported | HIA: 98.89 Caco2: 54.05 PPB: 85.16 BBB: 0.044 | [100] |
| Erigeron breviscapus (Vant.) | Viral spike protein and human ACE2 receptor inhibitor | Scutellarin | Not yet reported | HIA: 13.45 Caco2: 10.13 PPB: 72.90 BBB: 0.029 | [121] |
| Soya bean (Glycine max) | Viral spike protein and human ACE2 receptor inhibitor | Nicotinamine | 84 nM | HIA: 92.94 Caco2: 20.36 PPB: 2.02 BBB: 0.33 | [122] |
| Licorice root (Glycyrrhiza radix) | Viral spike protein and human ACE2 receptor inhibitor | Glycyrrhizin (saponin) | Not yet reported | HIA: 38.22 Caco2: 20.37 PPB: 88.72 BBB: 0.055 | [121] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muchtaridi, M.; Fauzi, M.; Khairul Ikram, N.K.; Mohd Gazzali, A.; Wahab, H.A. Natural Flavonoids as Potential Angiotensin-Converting Enzyme 2 Inhibitors for Anti-SARS-CoV-2. Molecules 2020, 25, 3980. https://doi.org/10.3390/molecules25173980
Muchtaridi M, Fauzi M, Khairul Ikram NK, Mohd Gazzali A, Wahab HA. Natural Flavonoids as Potential Angiotensin-Converting Enzyme 2 Inhibitors for Anti-SARS-CoV-2. Molecules. 2020; 25(17):3980. https://doi.org/10.3390/molecules25173980
Chicago/Turabian StyleMuchtaridi, Muchtaridi, M. Fauzi, Nur Kusaira Khairul Ikram, Amirah Mohd Gazzali, and Habibah A. Wahab. 2020. "Natural Flavonoids as Potential Angiotensin-Converting Enzyme 2 Inhibitors for Anti-SARS-CoV-2" Molecules 25, no. 17: 3980. https://doi.org/10.3390/molecules25173980
APA StyleMuchtaridi, M., Fauzi, M., Khairul Ikram, N. K., Mohd Gazzali, A., & Wahab, H. A. (2020). Natural Flavonoids as Potential Angiotensin-Converting Enzyme 2 Inhibitors for Anti-SARS-CoV-2. Molecules, 25(17), 3980. https://doi.org/10.3390/molecules25173980