
Contents of Functionally Bioactive Peptides, Free Amino Acids, and Biogenic Amines in Dutch-Type Cheese Models Produced with Different Lactobacilli


Abstract
:1. Introduction
2. Results and Discussion
2.1. Contents of Free Amino Acids in Cheese Models Examined
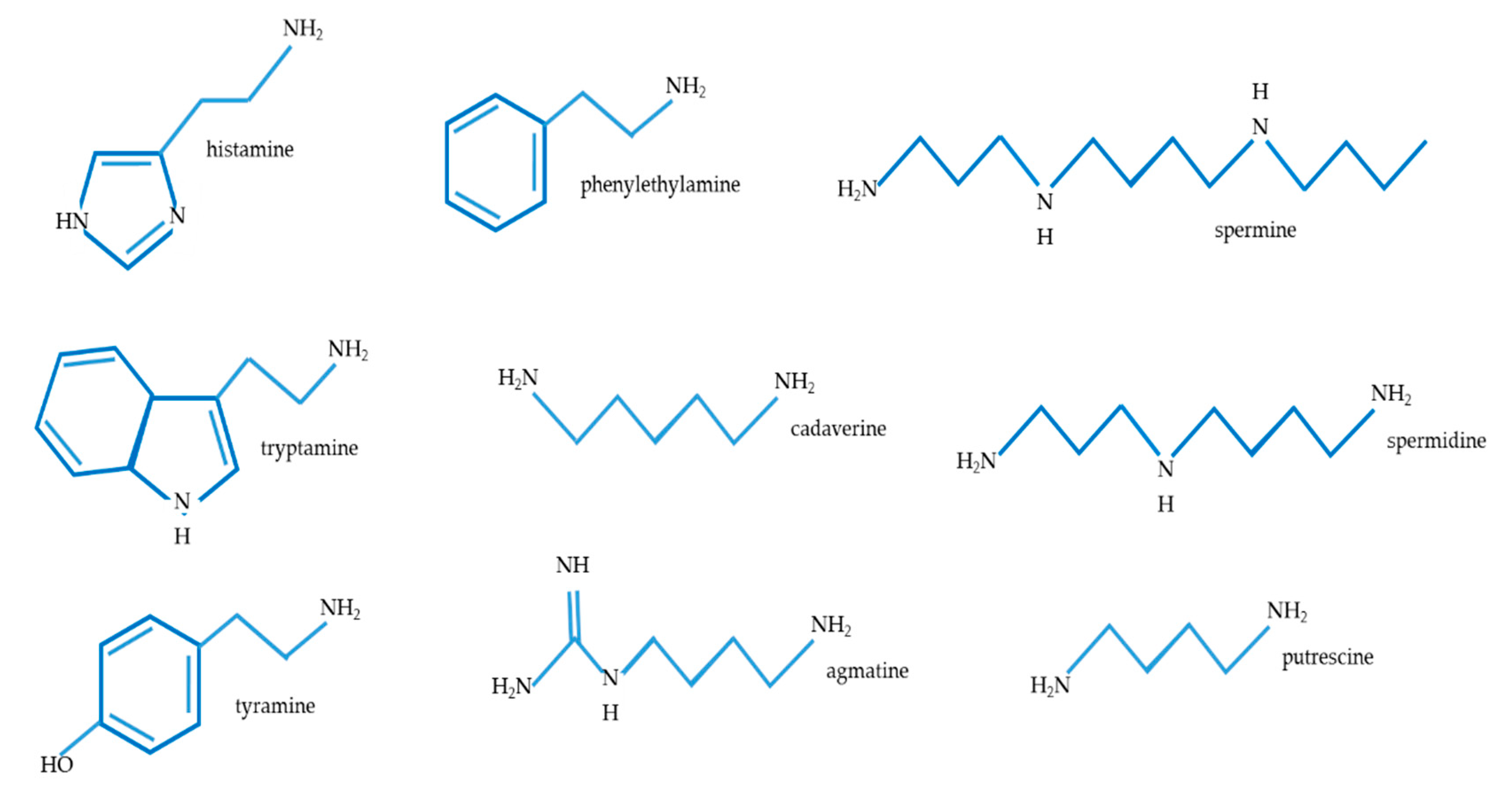
2.2. The Presence of Biogenic Amines in Cheese Models Examined
2.3. The Presence of Anserine and Carnosine in Cheese Models Examined
3. Material and Methods
3.1. Experimental Material
3.2. Preparation of Cheese Models
3.3. Determination of Contents of Free Amino Acids, Biogenic Amines, and Bioactive Dipeptides in Cheese Models Using Liquid Chromatography Coupled with Electrospray Ionization Ion Trap Tandem Mass Spectrometry (LC-ESI-IT-MS/MS)
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P.L. Biochemistry of cheese ripening. In Fundamentals of Cheese Science; Springer: New York, NY, USA, 2017; pp. 391–442. [Google Scholar]
- Ardö, Y.; McSweeney, P.L.; Magboul, A.A.; Upadhyay, V.K.; Fox, P.F. Biochemistry of cheese ripening: Proteolysis. In Cheese; Academic Press: Cambridge, MA, USA, 2017; pp. 445–482. [Google Scholar]
- Diana, M.; Rafecas, M.; Arco, C.; Quílez, J. Free amino acid profile of Spanish artisanal cheeses: Importance of gamma-aminobutyric acid (GABA) and ornithine content. J. Food Compos. Anal. 2014, 35, 94–100. [Google Scholar] [CrossRef]
- Garbowska, M.; Pluta, A.; Berthold-Pluta, A. Impact of Nisin-Producing Strains of Lactococcus lactis on the Contents of Bioactive Dipeptides, Free Amino Acids, and Biogenic Amines in Dutch-Type Cheese Models. Materials 2020, 13, 1835. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Iwaniak, A.; Minkiewicz, P. Proteins as the source of physiologically and functionally active peptides. Acta Sci. Pol. Technol. Aliment. 2007, 6, 5–15. [Google Scholar]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Fontenele, M.A.; do SR Bastos, M.; dos Santos, K.M.; Bemquerer, M.P.; do Egito, A.S. Peptide profile of Coalho cheese: A contribution for Protected Designation of Origin (PDO). Food Chem. 2017, 219, 382–390. [Google Scholar] [CrossRef]
- Pisanu, S.; Pagnozzi, D.; Pes, M.; Pirisi, A.; Roggio, T.; Uzzau, S.; Addis, M.F. Differences in the peptide profile of raw and pasteurised ovine milk cheese and implications for its bioactive potential. Int. Dairy J. 2015, 42, 26–33. [Google Scholar] [CrossRef]
- Sadat-Mekmene, L.; Richoux, R.; Aubert-Frogerais, L.; Madec, M.-N.; Corre, C.; Piot, M.; Gagnaire, V. Lactobacillus helveticus as a tool to change proteolysis and functionality in Swiss-type cheeses. J. Dairy Sci. 2013, 96, 1455–1470. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef]
- Corona, C.; Frazzini, V.; Silvestri, E.; Lattanzio, R.; La Sorda, R.; Piantelli, M.; Canzoniero, L.M.; Ciavardelli, D.; Rizzarelli, E.; Sensi, S.L. Effects of dietary supplementation of carnosine on mitochondrial dysfunction, amyloid pathology, and cognitive deficits in 3xTg-AD mice. PLoS ONE 2011, 6, 17971. [Google Scholar] [CrossRef] [Green Version]
- Mori, M.; Mizuno, D.; Konoha-Mizuno, K.; Sadakane, Y.; Kawahara, M. Quantitative analysis of carnosine and anserine in foods by performing high performance liquid chromatography. Biomed. Res. Trace Elem. 2015, 26, 147–152. [Google Scholar]
- Iovine, B.; Iannella, M.L.; Nocella, F.; Pricolo, M.R.; Bevilacqua, M.A. Carnosine inhibits KRAS-mediated HCT116 proliferation by affecting ATP and ROS production. Cancer Lett. 2012, 315, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Roberts, P.R.; Zaloga, G.P. Cardiovascular effects of carnosine. Biochemistry 2000, 65, 856–861. [Google Scholar] [PubMed]
- Buñková, L.; Buñka, F.; Mantlová, G.; Cablová, A.; Sedláce, I.; Svec, P.; Pachlová, V.; Kraácmar, S. The effect of ripening and storage conditions on the distribution of tyramine: Putrescine and cadaverine in Edam-cheese. Food Microbiol. 2010, 27, 880–888. [Google Scholar] [CrossRef]
- Ladero, V.; Calles-Enríquez, M.; Fernández, M.; Álvarez, M.A. Toxicological effects of dietary biogenic amines. Curr. Nutr. Food Sci. 2010, 6, 145–156. [Google Scholar] [CrossRef]
- Poveda, J.M.; Chicón, R.; Cabezas, L. Biogenic amine content and proteolysis in Manchego cheese manufactured with Lactobacillus paracasei subsp. paracasei as adjunct and other autochthonous strains as starters. Int. Dairy J. 2015, 47, 94–101. [Google Scholar] [CrossRef]
- Spizzirri, U.G.; Restuccia, D.; Curcio, M.; Parisi, O.I.; Iemma, F.; Picci, N. Determination of biogenic amines in different cheese samples by LC with evaporative light scattering detector. J. Food Compos. Anal. 2013, 29, 43–51. [Google Scholar] [CrossRef]
- Mayer, H.K.; Fiechter, G. UHPLC analysis of biogenic amines in different cheese varieties. Food Control. 2018, 93, 9–16. [Google Scholar] [CrossRef]
- Buñková, L.; Adamcová, G.; Hudcová, K.; Velichová, H.; Pachlová, V.; Lorencová, E.; Buñka, F. Monitoring of biogenic amines in cheeses manufactured at small-scale farms and in fermented dairy products in the Czech Republic. Food Chem. 2013, 141, 548–551. [Google Scholar] [CrossRef]
- Piras, C.; Marincola, F.C.; Savorani, F.; Engelsen, S.B.; Cosentino, S.; Viale, S.; Pisano, M.B. A NMR metabolomics study of the ripening process of the Fiore Sardo cheese produced with autochthonous adjunct cultures. Food Chem. 2013, 141, 2137–2147. [Google Scholar] [CrossRef]
- Renes, E.; Diezhandino, I.; Fernández, D.; Ferrazza, R.E.; Tornadijo, M.E.; Fresno, J.M. Effect of autochthonous starter cultures on the biogenic amine content of ewe's milk cheese throughout ripening. Food Microbiol. 2014, 44, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Linares, D.M.; Cruz Martin, M.; Ladero, V.; Alvarez, M.A.; Fernandez, M. Biogenic amines in dairy products. Crit. Rev. Food Sci. Nutr. 2011, 51, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Burdychova, R.; Komprda, T. Biogenic amine-forming microbial communities in cheese. FEMS Microbiol. Lett. 2007, 276, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Foulquié Moreno, M.R.; Sarantinopoulos, P.; Tsakalidou, E.; De Vuyst, L. The role and application of enterococci in food and health. Int. J. Food Microbiol. 2006, 106, 1–24. [Google Scholar] [CrossRef]
- McCabe-Sellers, B.; Staggs, C.G.; Bogle, M.L. Tyramine in foods and monoamine oxidase inhibitor drugs: A crossroad where medicine, nutrition, pharmacy, and food industry converge. J. Food Compos. Anal. 2006, 19, S58–S65. [Google Scholar] [CrossRef]
- Marcobal, A.; de Las Rivas, B.; Landete, J.M.; Tabera, L.; Muñoz, R. Tyramine and phenylethylamine biosynthesis by food bacteria. Crit. Rev. Food Sci. Nutr. 2012, 52, 448–467. [Google Scholar] [CrossRef] [Green Version]
- Linares, D.M.; del Rio, B.; Redruello, B.; Ladero, V.; Martin, M.C.; Fernandez, M.; Ruas-Madiedo, P.; Alvarez, M.A. Comparative analysis of the in vitro cytotoxicity of the dietary biogenic amines tyramine and histamine. Food Chem. 2016, 197 Pt A, 658–663. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, G.I.; Bashammakh, A.S.; Alsibaai, A.A.; Alwael, H.; El-Shahawi, M.S. A critical overview on the chemistry, clean-up and recent advances in analysis of biogenic amines in foodstuffs. TRAC-Trends Anal. Chem. 2016, 78, 84–94. [Google Scholar] [CrossRef]
- Mayer, H.K.; Fiechter, G.; Fischer, E. A new ultra-pressure liquid chromatography method for the determination of biogenic amines in cheese. J. Chromatogr. A 2010, 1217, 3251–3257. [Google Scholar] [CrossRef]
- Herrero-Fresno, A.; Martínez, N.; Sánchez-Llana, E.; Díaz, M.; Fernández, M.; Martin, M.C.; Ladero, V.; Alvarez, M.A. Lactobacillus casei strains isolated from cheese reduce biogenic amine accumulation in an experimental model. Int. J. Food Microbiol. 2012, 157, 297–304. [Google Scholar] [CrossRef]
- Michaelidou, A.; Katsiari, M.C.; Kondyli, E.; Voutsinas, L.P.; Alichanidis, E. Effect of a commercial adjunct culture on proteolysis in low-fat Feta-type cheese. Int. Dairy J. 2003, 13, 179–189. [Google Scholar] [CrossRef]
- Madrau, M.A.; Mangia, N.P.; Murgia, M.A.; Sanna, M.G.; Garau, G.; Leccis, L.; Caredda, M.; Deiana, P. Employment of autochthonous microflora in Pecorino Sardo cheese manufacturing and evolution of physicochemical parameters during ripening. Int. Dairy J. 2006, 16, 876–885. [Google Scholar] [CrossRef]
- Kurata, K.; Nagasawa, M.; Tomonaga, S.; Aoki, M.; Morishita, K.; Denbow, D.M.; Furuse, M. Orally administered L-ornithine elevates brain L-ornithine levels and has an anxiolytic-like effect in mice. Nutr. Neurosci. 2011, 14, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Tillakaratne, N.J.; Medina-Kauwe, L.; Gibson, K.M. gamma-Aminobutyric acid (GABA) metabolism in 276 mammalian neural and nonneural tissues. Comp. Biochem. Physiol. A Physiol. 1995, 112, 247–263. [Google Scholar] [CrossRef]
- Cui, R.; Li, B.; Suemaru, K.; Araki, H. Psychological stress-induced changes in sleep patterns and their generation mechanism. Yakugaku Zasshi 2008, 128, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Miao, K.; Niyaphorn, S.; Qu, X. Production of Gamma-Aminobutyric Acid from Lactic Acid Bacteria: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.J.; Steele, J.L. Fermented Dairy Products. In Food Microbiology; Doyle, M.P., Buchanan, R.L., Eds.; ASM Press: Washington, DC, USA, 2013; pp. 825–839. [Google Scholar]
- Pachlová, V.; Buňková, L.; Flasarová, R.; Salek, R.N.; Dlabajová, A.; Butor, I.; Buňkaa, F. Biogenic amine production by nonstarter strains of Lactobacillus curvatus and Lactobacillus paracasei in the model system of Dutch-type cheese. LWT—Food Sci. Technol. 2018, 97, 730–735. [Google Scholar] [CrossRef]
- Romano, A.; Ladero, V.; Alvarez, M.A.; Lucas, P.M. Putrescine production via the ornithine decarboxylation pathway improves the acid stress survival of Lactobacillus brevis and is part of a horizontally transferred acid resistance locus. Int. J. Food Microbiol. 2014, 175, 14–19. [Google Scholar] [CrossRef]
- Diaz, M.; del Rio, B.; Ladero, V.; Redruello, B.; Fernández, M.; Martin, M.C.; Alvarez, M.A. Isolation and typification of histamine-producing Lactobacillus vaginalis strains from cheese. Int. J. Food Microbiol. 2015, 215, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Grandvalet, C.; Coton, E.; Coton, M.; Barnavon, L.; Bach, B.; et al. Biogenic amines in fermented foods. Eur. J. Clin. Nutr. 2010, 64, 64–951. [Google Scholar] [CrossRef] [Green Version]
- Ladero, V.; Linares, D.M.; Fernández, M.; Alvarez, M.A. Real time quantitative PCR detection of histamine-producing lactic acid bacteria in cheese: Relation with histamine content. Food Res. Int. 2008, 41, 1015–1019. [Google Scholar] [CrossRef]
- Linares, D.M.; del Rio, B.; Redruello, B.; Fernández, M.; Martin, M.C.; Ladero, V.; Alvarez, M.A. The use of qPCR-based methods to identify and quantify food spoilage microorganisms. In Novel Food Preservation and Microbial Assessment Techniques; Boziaris, I.S., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 313–334. [Google Scholar]
- Yılmaz, C.; Gökmen, V. Formation of tyramine in yoghurt during fermentation—Interaction between yoghurt starter bacteria and Lactobacillus Plant. Food Res. Int. 2017, 97, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.A.; Moreno-Arribas, M.V. The problem of biogenic amines in fermented foods and the use of potential biogenic amine-degrading microorganisms as a solution. Trends Food Sci. Technol. 2014, 39, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Ladero, V.; Herrero-Fresno, A.; Martinez, N.; del Río, B.; Linares, D.M.; Fernández, M.; Martin, M.C.; Alvarez, M.A. Genome sequence analysis of the biogenic amine-degrading strain Lactobacillus casei 5b. Genome Announc. 2014, 2, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Redruello, B.; Ladero, V.; Cuesta, I.; Álvarez-Buylla, J.R.; Cruz Martín, M.; Fernández, M.; Alvarez, M.A. A fast, reliable, ultra high performance liquid chromatography method for the simultaneous determination of amino acids, biogenic amines and ammonium ions in cheese, using diethyl ethoxymethylenemalonate as a derivatising agent. Food Chem. 2013, 139, 1029–1035. [Google Scholar] [CrossRef]
- Comas-Basté, O.; Sánchez-Pérez, S.; Veciana-Nogués, M.T.; Latorre-Moratalla, M.; Vidal-Carou, M.C. Histamine Intolerance: The Current State of the Art. Biomolecules 2020, 10, 1181. [Google Scholar] [CrossRef]
- Moret, S.; Bortolomeazzi, R.; Feruglio, M.; Lercker, G. Determination of biogenic amines in Italian cheeses. Sci. Tec. Latt. Casearia 1992, 43, 187–198. [Google Scholar]
- Novella-Rodríguez, S.; Veciana-Nogués, M.T.; Roig-Sagués, A.X.; Trujillo-Mesa, A.J.; Vidal-Carou, M.C. Evaluation of biogenic amines and microbial counts throughout the ripening of goat cheeses from pasteurized and raw milk. J. Dairy Res. 2004, 71, 245–252. [Google Scholar] [CrossRef]
- Fernández, M.; Lìnares, D.M.; del Río, B.; Ladero, V.; Álvarez, M.A. HPLC quantification of biogenic amines in cheeses: Correlation with PCR-detection of tyramine-producing microorganisms. J. Dairy Res. 2007, 74, 276–282. [Google Scholar] [CrossRef]
- Garbowska, M.; Pluta, A.; Berthold-Pluta, A. Antihypertensive Peptide Activity in Dutch-Type Cheese Models Prepared with Different Additional Strains of Lactobacillus Genus Bacteria. Appl. Sci. Basel 2019, 9, 1679. [Google Scholar] [CrossRef] [Green Version]
- Garbowska, M.; Pluta, A.; Berthold-Pluta, A. Proteolytic and ACE-inhibitory activities of Dutch-type cheese models prepared with different strains of Lactococcus lactis. Food Biosci. 2020, 35, 1–7. [Google Scholar] [CrossRef]
Sample Availability: Examined samples are available from the authors. |


{kind=link}
| Added | Control | Lb. rhamnosus 489 | Lb. delbrueckii 490 | Lb. acidophilus 2499 | Lb. casei 2639 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Amino Acids | Time of Ripening (Week) | 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 |
| Alanine | 4.58 b ± 0.35 | 12.48 B ± 3.60 | 11.43 A ± 3.40 | 6.23 b ± 0.23 | 9.50 B ± 0.93 | 19.91 B ± 1.22 | ND a | 0.68 A ± 0.25 | 12.71 A ± 1.61 | 11.80 c ± 2.12 | 36.28 C ± 4.40 | 52.35 C ± 2.55 | 1.16 a ± 0.08 | 10.93 B ± 1.84 | 17.13AB ± 1.44 | |
| Arginine | 18.31 cd ± 2.40 | 17.68 A ± 3.28 | 16.59 A ± 3.34 | ND a | 16.51 A ± 0.56 | 16.81 A ± 1.24 | 20.86 d ± 1.28 | 18.33 A ± 1.12 | 16.28 A ± 0.38 | 15.56 bc ± 0.69 | 16.99 A ± 0.57 | 18.07 A ± 1.38 | 14.59 b ± 1.07 | 16.27 A ± 0.41 | 18.11 A ± 0.55 | |
| Asparagine | 73.11 a ± 5.57 | 77.93 A ± 3.39 | 88.81 A ± 5.50 | 91.29 b ± 1.87 | 132.66 D ± 2.20 | 129.17 C ± 1.81 | 83.14 ab ± 7.38 | 95.16 B ± 3.90 | 108.74 B ± 10.12 | 75.53 ab ± 5.03 | 110.65 C ± 4.01 | 127.54 C ± 5.55 | 77.59 ab ± 9.69 | 79.68 A ± 4.85 | 84.16 A ± 5.21 | |
| Asparagine acid | 16.56 a ± 0.39 | 31.45 B ± 3.58 | 54.50 C ± 5.38 | 72.03 d ± 1.96 | 113.21 D ± 4.00 | 119.89 E ±1.68 | 62.50 c ± 2.51 | 11.89 A ± 3.01 | 1.75 A ± 0.64 | 120.68 e ± 5.55 | 76.76 C ± 4.51 | 70.18 D ± 4.32 | 32.91 b ± 2.33 | 36.40 B ± 0.83 | 42.40 B ± 2.76 | |
| Glutamine | ND a | ND A | ND A | 20.22 b ± 3.59 | 41.21 D ± 1.59 | 38.71 C ± 2.25 | ND a | 13.34 B ± 0.34 | 12.74 B ± 0.42 | ND a | 24.88 C ± 3.54 | 34.47 C ± 4.85 | ND a | ND A | ND A | |
| Glutamic acid | 10.94 a ± 0.63 | 11.64 A ± 3.01 | 30.76 A ± 5.36 | 18.25 a ± 1.06 | 355.78 D ± 13.56 | 443.94 C ± 6.32 | 172.47 c ± 5.88 | 183.27 B ± 6.50 | 287.40 B ± 7.85 | 56.16 b ± 5.36 | 235.09 C ± 12.61 | 571.11 D ± 21.53 | 8.68 a ± 0.33 | 13.98 A ± 1.66 | 36.51 A ± 3.38 | |
| Glycine | 0.30 a ± 0.09 | 0.32 A ± 0.18 | 0.49 A ± 0.19 | 5.04 b ± 0.12 | 14.06 C ± 1.61 | 14.26 C ± 2.46 | 0.36 a ± 0.05 | 3.85 B ± 0.77 | 1.97 A ± 0.59 | 0.30 a ± 0.27 | 3.37 B ± 0.50 | 6.00 B ± 0.22 | 0.14 a ± 0.01 | 1.05 A ± 0.06 | 1.30 A ± 0.01 | |
| Histidine | 0.13 a ± 0.03 | 28.31 B ± 4.70 | 123.91 C ± 5.55 | ND a | ND A | 45.82 B ± 3.15 | ND a | 69.27 D ± 3.16 | 144.86 D ± 4.88 | 125.24 b ± 5.11 | 52.18 C ± 3.38 | 43.02 B ± 4.54 | ND a | ND A | ND A | |
| Leucine | 88.12 c ± 6.31 | 97.22 D ± 2.82 | 105.94 C ± 4.13 | 35.71 a ± 6.03 | 66.32 B ± 1.85 | 88.01 B ± 4.09 | 61.53 b ± 0.83 | 64.30 B ± 2.84 | 64.33 A ± 4.52 | 44.05 a ± 4.37 | 52.37 A ± 3.09 | 58.49 A ± 3.40 | 72.06 b ± 2.54 | 83.82 C ± 2.44 | 86.85 B ± 2.04 | |
| Isoleucine | 84.40 d ± 5.58 | 77.09 C ± 6.79 | 70.88 B ± 4.08 | 55.42 b ± 3.04 | 67.22 BC ± 3.84 | 77.01 B ± 2.59 | ND a | 54.20 A ± 0.87 | 47.62 A ± 3.71 | 66.02 c ± 4.83 | 59.42 AB ± 4.35 | 52.80 A ± 2.71 | 66.61 c ± 1.51 | 75.81 C ± 5.71 | 79.42 B ± 3.02 | |
| Lysine | 15.91 b ± 5.54 | 18.15 B ± 4.83 | 30.41 B ± 7.26 | ND a | ND A | 19.84 AB ± 2.05 | 24.63 c ± 0.16 | 29.38 C ± 6.13 | 129.15 C ± 5.12 | 29.85 c ± 2.27 | 40.54 D ± 3.36 | 137.26 C ± 5.19 | ND a | 1.70 A ± 0.10 | 9.16 A ± 0.40 | |
| Methionine | 10.16 a ± 2.67 | 12.77AB ± 5.42 | 15.06 A ± 1.97 | 11.94 a ± 1.68 | 20.41 B ± 1.37 | 27.94 B ± 7.65 | 7.15 a ± 0.27 | 18.49AB ± 1.57 | 13.25 A ± 0.75 | 40.61 b ± 4.55 | 18.62 AB ± 2.56 | 15.71 A ± 0.95 | 8.64 a ± 0.45 | 11.01 A ± 2.27 | 15.37 A ± 2.50 | |
| Phenylalanine | 3.35 a ± 3.21 | 26.32 A ± 3.55 | 41.65 B ± 4.03 | 33.27 b ± 1.99 | 47.16 B ± 3.54 | 67.74 D ± 1.36 | 42.66 c ± 3.49 | 44.83 B ± 1.24 | 34.28 AB ± 1.73 | 46.10 c ± 2.32 | 49.21 B ± 1.06 | 58.13 C ± 4.38 | ND a | 24.58 A ± 3.21 | 27.20 A ± 1.41 | |
| Proline | 24.40 a ± 5.54 | 37.74 B ± 4.50 | 38.01 A ± 3.00 | 66.40 c ± 5.45 | 69.31 C ± 2.71 | 96.57 C ± 3.87 | 48.73 b ± 5.14 | 68.80 C ± 0.44 | 79.51 B ± 0.72 | 80.00 d ± 0.96 | 75.19 C ± 4.10 | 72.92 B ± 2.86 | 22.39 a ± 1.74 | 23.88 A ± 3.25 | 31.45 A ± 1.09 | |
| Serine | ND a | ND A | ND A | 11.54 b ± 1.85 | 22.95 D ± 1.42 | 24.81 D ± 2.01 | ND a | 6.51 B ± 0.42 | 5.52 B ± 0.30 | ND a | 11.81 C ± 1.88 | 20.18 C ± 1.82 | ND a | ND A | ND A | |
| Threonine | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Tryptophan | ND a | ND A | 20.89 A ± 2.64 | ND a | 20.08 B ± 1.70 | 21.71 A ± 0.80 | 38.43 c ± 1.24 | 20.86 B ± 0.90 | 22.57 A ± 2.74 | 33.38 b ± 2.84 | 28.78 C ± 0.90 | 21.96 A ± 1.19 | ND a | 19.97 B ± 1.40 | 21.94 A ± 1.32 | |
| Tyrosine | ND a | ND A | ND A | 42.05 b ± 1.95 | 62.61 B ± 5.10 | 38.33 B ± 2.60 | ND a | ND A | ND A | 86.90 c ± 4.87 | 77.11 C ± 1.48 | 52.00 C ± 2.04 | 3.65 a ± 0.35 | ND A | ND A | |
| Valine | 30.04 a ± 4.34 | 36.32 AB ± 5.44 | 34.27 A ± 4.22 | 47.32 b ± 2.01 | 100.48 D ± 7.25 | 145.05 C ± 5.90 | 23.78 a ± 3.55 | 44.09 B ± 3.40 | 41.63 A ± 2.95 | 63.02 c ± 2.49 | 67.03 C ± 1.57 | 76.75 B ± 4.67 | 22.83 a ± 3.19 | 30.82 A ± 1.27 | 37.89 A ± 1.73 | |
| Cystine | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Taurine | 13.58 a ± 1.14 | 29.50 B ± 4.89 | 49.87 D ± 0.93 | 10.50 a ± 0.69 | 12.89 A ± 1.66 | 15.85 A ± 1.45 | 11.10 a ± 2.03 | 12.62 A ± 1.43 | 15.55 A ± 0.54 | 38.70 c ± 1.66 | 35.63 B ± 2.80 | 30.00 B ± 1.38 | 18.34 b ± 1.09 | 32.91 B ± 2.18 | 38.78 C ± 3.58 | |
| Citrulline | 21.28 a ± 2.22 | 25.93 A ± 5.96 | 29.73 B ± 3.91 | 22.76 a ± 2.26 | 28.26 A ± 2.89 | 26.86 AB ± 1.42 | 29.60 b ± 1.17 | 25.55 A ± 1.11 | 22.59 A ± 0.41 | 25.14 ab ± 1.66 | 23.40 A ± 0.87 | 21.91 A ± 1.55 | 22.80 a ± 1.07 | 23.33 A ± 0.78 | 27.07 AB ± 3.11 | |
| Sarcosine | 32.78 b ± 8.88 | ND A | ND A | ND a | ND A | ND A | 45.32 c ± 2.96 | 41.64 B ± 2.63 | 33.26 B ± 2.64 | 36.28 bc ± 2.83 | ND A | ND A | 37.52 bc ± 1.67 | ND A | ND A | |
| Ornithine | ND a | 33.36 A ± 3.71 | 37.72 A ± 3.39 | 46.03 b ± 1.13 | 54.96 B ± 3.98 | 77.60 D ± 3.38 | 103.43 d ± 3.13 | 54.50 B ± 3.64 | 46.44 B ± 3.33 | 88.20 c ± 3.21 | 57.67 B ± 4.18 | 55.78 C ± 2.42 | ND a | 34.39 A ± 0.72 | 36.28 A ± 2.74 | |
| Total free amino acids (TFAA) | 447.94 a ± 16.08 | 574.22 A ± 9.36 | 800.92 B ± 28.63 | 595.99 b ± 20.13 | 1255.57 D ± 30.56 | 1555.83 D ± 33.50 | 775.69 c ± 6.67 | 881.57 B ± 19.30 | 1142.14 C ± 15.02 | 1083.51d ± 25.40 | 1152.98C ± 38.18 | 1596.64 D ± 53.44 | 409.91 a ± 7.57 | 520.53 A ± 15.38 | 611.02 A ± 20.54 | |
| Added | Control | Lb. rhamnosus 489 | Lb. delbrueckii 490 | Lb. acidophilus 2499 | Lb. casei 2639 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Biogenic Amines | Time of Ripening (Week) | 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 |
| Agmatine | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Phenylethylamine | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Histamine | 17.69 a ± 0.72 | 22.33 A ± 2.25 | 40.98 A ± 4.93 | 36.21 c ± 1.61 | 58.46 D ± 3.05 | 71.64 B ± 3.31 | 57.07 d ± 1.66 | 51.83 C ± 2.14 | 41.05 A ± 2.26 | 53.25 d ± 1.68 | 51.50 C ± 1.73 | 46.61 A ± 3.26 | 27.03 b ± 1.58 | 32.16 B ± 2.12 | 45.51 A ± 2.16 | |
| Cadaverine | 0.64 a ± 0.09 | 0.78 AB ± 0.10 | 4.62 B ± 0.91 | 0.67 a ± 0.06 | 0.69 A ± 0.01 | 0.69 A ± 0.00 | 4.36 b ± 0.30 | 1.07 C ± 0.13 | 0.41 A ± 0.10 | 5.46 c ± 0.34 | 0.87 ABC ± 0.08 | 0.54 A ± 0.06 | 0.59 a ± 0.07 | 0.92 BC ± 0.04 | 0.89 A ± 0.02 | |
| Putrescine | 0.32 b ± 0.10 | 0.38 B ± 0.30 | 3.05 B ± 0.28 | 0.21 b ± 0.00 | ND A | ND A | ND a | ND A | 4.62 C ± 1.06 | 1.17 c ± 0.10 | 0.27 AB ± 0.08 | ND A | ND a | 0.64 B ± 0.04 | 0.46 A ± 0.02 | |
| Spermidine | 1.62 d ± 0.12 | 1.17 D ± 0.23 | 0.48 A ± 0.03 | 0.29 b ± 0.01 | 0.40 B ± 0.04 | 0.37 A ± 0.01 | ND a | ND A | 0.92 B ± 0.11 | ND a | 0.76 C ± 0.10 | 1.19 B ± 0.31 | 0.56 c ± 0.05 | 1.08 D ± 0.04 | 1.03 B ± 0.01 | |
| Spermine | 1.60 b ± 0.10 | 0.31 BC ± 0.10 | ND A | 0.29 a ± 0.10 | 0.23 B ± 0.00 | ND A | 3.26 c ± 0.26 | ND A | ND A | 1.30 b ± 0.04 | 0.37 C ± 0.04 | 0.34 B ± 0.05 | ND a | 0.21 B ± 0.01 | 0.56 C ± 0.06 | |
| Tryptamine | 2.49 b ± 0.36 | 3.88 C ± 1.08 | ND A | 6.95 c ± 0.45 | 4.85 C ± 0.29 | 4.70 C ± 0.16 | ND a | ND A | 1.72 B ± 0.35 | ND a | 1.88 B ± 0.07 | ND A | ND a | 5.03 C ± 0.06 | ND A | |
| Tyramine | 1.21 b ± 0.06 | 1.32 AB ± 0.37 | 4.29 B ± 0.31 | 0.43 a ± 0.04 | ND A | ND A | 2.34 c ± 0.11 | 6.77 C ± 1.26 | ND A | 4.80 d ± 0.25 | 1.87 B ± 0.08 | ND A | 0.42 a ± 0.03 | ND A | ND A | |
| Total BA | 25.56 a ± 1.13 | 30.17 A ± 3.19 | 53.41 A ± 4.78 | 45.05 b ± 2.21 | 64.64 C ± 3.18 | 77.41 B ± 3.41 | 67.03 c ± 1.22 | 59.67 C ± 2.70 | 48.72 A ± 3.16 | 65.98 c ± 1.12 | 57.52 C ± 1.73 | 48.68 A ± 3.09 | 28.61 a ± 1.63 | 40.04 B ± 2.13 | 48.45 A ± 2.10 | |
| Added | Control | Lb. rhamnosus 489 | Lb. delbrueckii 490 | Lb. acidophilus 2499 | Lb. casei 2639 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bioactive Peptides | Time of Ripening (Week) | 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 |
| Anserine | 66.33 b ± 3.79 | 85.17 A ± 2.98 | 79.29 A ± 2.62 | 71.29 b ± 0.96 | 114.97 B ± 5.44 | 99.20 B ± 2.54 | ND a | 101.76 B ± 7.54 | 92.96 AB ± 10.09 | 93.04 d ± 3.12 | 114.34 B ± 4.86 | 119.02 C ± 2.17 | 83.55 c ± 3.57 | 82.55 A ± 7.13 | 80.64 A ± 3.56 | |
| L-carnosine | 8.37 b ± 1.07 | ND A | ND A | 33.08 c ± 2.80 | 32.53 C ± 2.36 | 3.74 B ± 0.51 | ND a | ND A | ND A | ND a | 8.74 B ± 0.75 | 17.09 C ± 0.94 | ND a | ND A | ND A | |
| Total BP | 74.70 b ± 4.87 | 85.17 AB ± 2.98 | 79.29 A ± 2.62 | 104.37 e ± 2.28 | 147.50 D ± 7.78 | 102.94 B ± 3.00 | ND a | 101.76 B ± 7.54 | 92.96 AB ± 10.09 | 93.04 d ± 3.12 | 123.07 C ± 5.39 | 136.11 C ± 2.67 | 83.55 c ± 3.57 | 82.55 A ± 7.13 | 80.64 A ± 3.56 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garbowska, M.; Pluta, A.; Berthold-Pluta, A. Contents of Functionally Bioactive Peptides, Free Amino Acids, and Biogenic Amines in Dutch-Type Cheese Models Produced with Different Lactobacilli. Molecules 2020, 25, 5465. https://doi.org/10.3390/molecules25225465
Garbowska M, Pluta A, Berthold-Pluta A. Contents of Functionally Bioactive Peptides, Free Amino Acids, and Biogenic Amines in Dutch-Type Cheese Models Produced with Different Lactobacilli. Molecules. 2020; 25(22):5465. https://doi.org/10.3390/molecules25225465
Chicago/Turabian StyleGarbowska, Monika, Antoni Pluta, and Anna Berthold-Pluta. 2020. "Contents of Functionally Bioactive Peptides, Free Amino Acids, and Biogenic Amines in Dutch-Type Cheese Models Produced with Different Lactobacilli" Molecules 25, no. 22: 5465. https://doi.org/10.3390/molecules25225465
APA StyleGarbowska, M., Pluta, A., & Berthold-Pluta, A. (2020). Contents of Functionally Bioactive Peptides, Free Amino Acids, and Biogenic Amines in Dutch-Type Cheese Models Produced with Different Lactobacilli. Molecules, 25(22), 5465. https://doi.org/10.3390/molecules25225465