
In Vitro and In Vivo Effects of Flavonoids on Peripheral Neuropathic Pain



Abstract
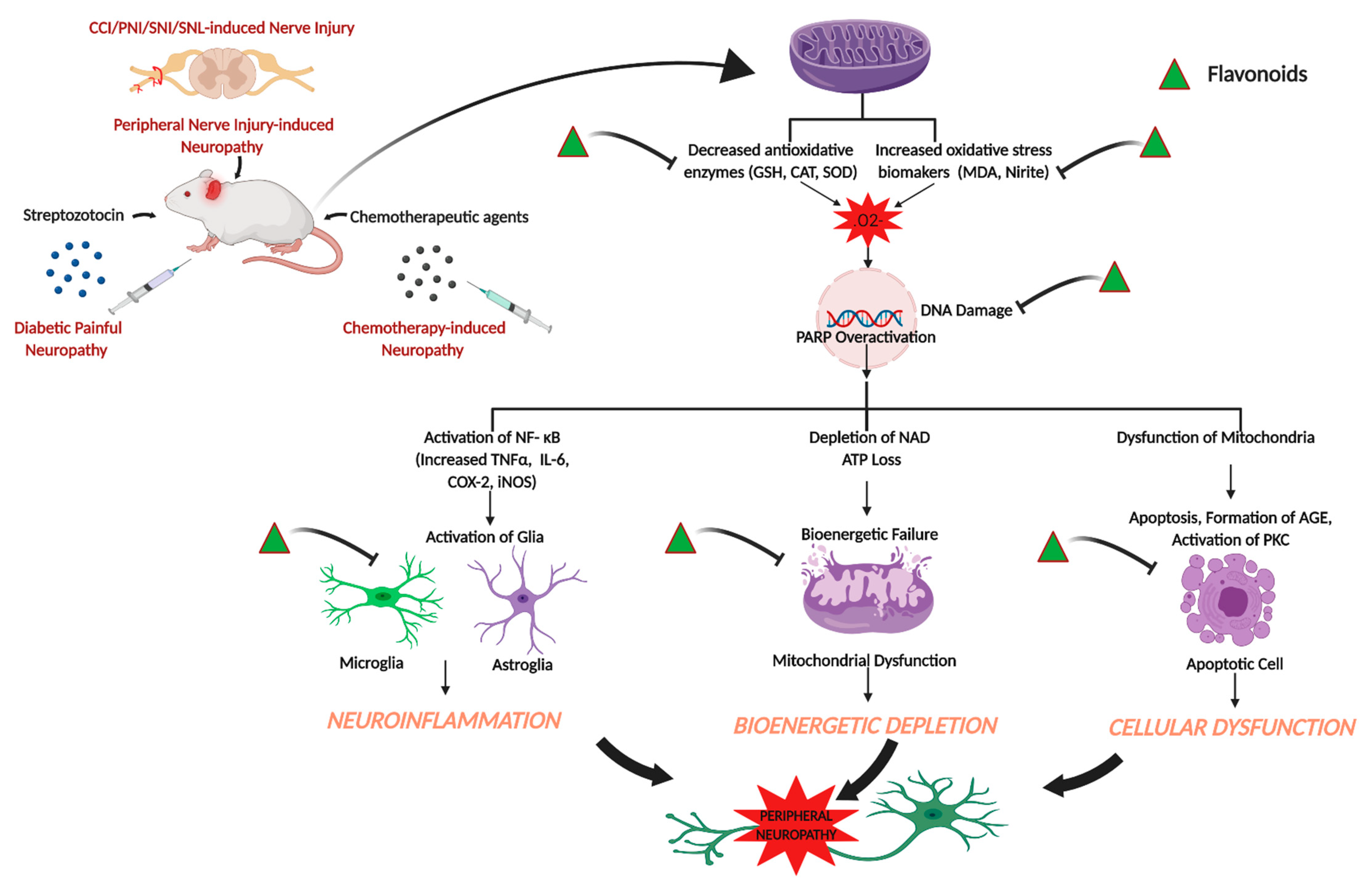
:1. Introduction
2. Flavonoids Classification, Structure, and Dietary Sources
3. Biosynthesis of Flavonoids
4. Neuropathic Pain
5. Chemotherapy-Induced Peripheral Neuropathy (CIPN)
Effects of Flavonoids on CIPN
6. Diabetic Painful Neuropathy
Effects of Flavonoids on Diabetic Painful Neuropathy
7. Other Peripheral Nerve Injury Models
7.1. Sciatic Nerve Chronic Constriction Injury (CCI)
7.2. Effects of Flavonoids on Sciatic Nerve CCI Model
7.3. Partial Sciatic Nerve Injury (PNI)
7.4. Spared Nerve Injury (SNI)
7.5. Spinal Nerve Ligation (SNL)
7.6. Effects of Flavonoids on Other Peripheral Neuropathic Pain Models
8. Future Directions
9. Overall Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- International Association for the Study of Pain Neuropathic Pain. Available online: https://www.iasp-pain.org/GlobalYear/NeuropathicPain (accessed on 5 March 2020).
- Gold, M.; Gebhart, G.; Fishman, S. Bonica’s Management of Pain; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2010; pp. 24–34. [Google Scholar]
- Levine, J.D.; Fields, H.L.; Basbaum, A.I. Peptides and the primary afferent nociceptor. J. Neurosci. 1993, 13, 2273–2286. [Google Scholar] [CrossRef] [PubMed]
- Pitcher, M.H.; Von Korff, M.; Bushnell, M.C.; Porter, L. Prevalence and Profile of High-Impact Chronic Pain in the United States. J. Pain 2019, 20, 146–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B.; et al. Neuropathic pain. Nat. Rev. Dis Primers 2017, 3, 17002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Hecke, O.; Austin, S.K.; Khan, R.A.; Smith, B.H.; Torrance, N. Neuropathic pain in the general population: A systematic review of epidemiological studies. Pain 2014, 155, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Abbott, C.A.; Malik, R.A.; van Ross, E.R.; Kulkarni, J.; Boulton, A.J. Prevalence and characteristics of painful diabetic neuropathy in a large community-based diabetic population in the U.K. Diabetes Care 2011, 34, 2220–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvez, R.; Rejas, J.; Perez, M.; Gomez, M. [Prevalence of neuropathic pain in Spain: Clinical, working and health care implications]. Med. Clin. (Barc) 2005, 125, 221–229. [Google Scholar]
- Freynhagen, R.; Baron, R.; Tolle, T.; Stemmler, E.; Gockel, U.; Stevens, M.; Maier, C. Screening of neuropathic pain components in patients with chronic back pain associated with nerve root compression: A prospective observational pilot study (MIPORT). Curr. Med. Res. Opin. 2006, 22, 529–537. [Google Scholar] [CrossRef]
- Kaki, A.M.; El-Yaski, A.Z.; Youseif, E. Identifying Neuropathic Pain among Patients With Chronic Low-Back Pain: Use of the Leeds Assessment of Neuropathic Symptoms and Signs Pain Scale. Reg. Anesth. Amp. Pain Med. 2005, 30, 1–422. [Google Scholar] [CrossRef]
- Argoff, C.E.; Cole, B.E.; Fishbain, D.A.; Irving, G.A. Diabetic peripheral neuropathic pain: Clinical and quality-of-life issues. Mayo Clin. Proc. 2006, 81, S3–S11. [Google Scholar] [CrossRef]
- Montero, J.H.; Gutierrez-Rivas, E.; Pardo, J.F.; Navarro, C.D. Epidemiological study of prevalence, incidence and neuropathic pain characterization in neurology units. Prevadol study. Neurol. (Barc. Spain) 2005, 20, 385–389. [Google Scholar]
- Torrance, N.; Smith, B.H.; Bennett, M.I.; Lee, A.J. The epidemiology of chronic pain of predominantly neuropathic origin. Results from a general population survey. J. Pain 2006, 7, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.H.; Torrance, N. Epidemiology of neuropathic pain and its impact on quality of life. Curr. Pain Headache Rep. 2012, 16, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Meacham, K.; Shepherd, A.; Mohapatra, D.P.; Haroutounian, S. Neuropathic Pain: Central vs. Peripheral Mechanisms. Curr. Pain Headache Rep. 2017, 21, 28. [Google Scholar] [CrossRef]
- Harden, R.N. Gabapentin: A new tool in the treatment of neuropathic pain. Acta Neurol. Scand Suppl. 1999, 173, 43–47. [Google Scholar] [CrossRef]
- Attal, N. Pharmacologic treatment of neuropathic pain. Acta Neurol. Belg. 2001, 101, 53–64. [Google Scholar]
- Finnerup, N.B.; Attal, N.; Haroutounian, S.; McNicol, E.; Baron, R.; Dworkin, R.H.; Gilron, I.; Haanpaa, M.; Hansson, P.; Jensen, T.S.; et al. Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet Neurol 2015, 14, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Bril, V.; England, J.; Franklin, G.M.; Backonja, M.; Cohen, J.; Del Toro, D.; Feldman, E.; Iverson, D.J.; Perkins, B.; Russell, J.W.; et al. Rehabilitation, Evidence-based guideline: Treatment of painful diabetic neuropathy: Report of the American Academy of Neurology, the American Association of Neuromuscular and Electrodiagnostic Medicine, and the American Academy of Physical Medicine and Rehabilitation. Neurology 2011, 76, 1758–1765. [Google Scholar]
- Ngo, L.T.; Okogun, J.I.; Folk, W.R. 21st century natural product research and drug development and traditional medicines. Nat. Prod. Rep. 2013, 30, 584–592. [Google Scholar] [CrossRef]
- da Silva, K.A.B.S.; Manjavachi, M.N.; Paszcuk, A.F.; Pivatto, M.; Viegas, C.; Bolzani, V.S.; Calixto, J.B. Plant derived alkaloid (−)-cassine induces anti-inflammatory and anti-hyperalgesics effects in both acute and chronic inflammatory and neuropathic pain models. Neuropharmacology 2012, 62, 967–977. [Google Scholar] [CrossRef]
- Garg, G.; Adams, J.D. Treatment of neuropathic pain with plant medicines. Chin. J. Integr Med. 2012, 18, 565–570. [Google Scholar] [CrossRef]
- Liu, M.; Zhou, L.; Chen, Z.; Hu, C. Analgesic effect of iridoid glycosides from Paederia scandens (LOUR.) MERRILL (Rubiaceae) on spared nerve injury rat model of neuropathic pain. Pharm. Biochem. Behav. 2012, 102, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Yimam, M.; Brownell, L.; Hodges, M.; Jia, Q. Analgesic effects of a standardized bioflavonoid composition from Scutellaria baicalensis and Acacia catechu. J. Diet. Suppl. 2012, 9, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCallum, J.L.; Yang, R.; Young, J.C.; Strommer, J.N.; Tsao, R. Improved high performance liquid chromatographic separation of anthocyanin compounds from grapes using a novel mixed-mode ion-exchange reversed-phase column. J. Chromatogr. A 2007, 1148, 38–45. [Google Scholar] [CrossRef]
- Tsao, R.; McCallum, J. Chemistry of Flavonoids. U: Fruit and Vegetable Phytochemicals: Chemistry, Nutritional Value and Stability; Blackwell Publishing: Hoboken, NJ, USA, 2009. [Google Scholar]
- Tsao, R.; Yang, R.; Young, J.C.; Zhu, H. Polyphenolic Profiles in Eight Apple Cultivars Using High-Performance Liquid Chromatography (HPLC). J. Agric. Food Chem. 2003, 51, 6347–6353. [Google Scholar] [CrossRef]
- Zhao, F.; Watanabe, Y.; Nozawa, H.; Daikonnya, A.; Kondo, K.; Kitanaka, S. Prenylflavonoids and Phloroglucinol Derivatives from Hops (Humulus lupulus). J. Nat. Prod. 2005, 68, 43–49. [Google Scholar] [CrossRef]
- Kawaii, S.; Tomono, Y.; Katase, E.; Ogawa, K.; Yano, M. Quantitation of Flavonoid Constituents in Citrus Fruits. J. Agric. Food Chem. 1999, 47, 3565–3571. [Google Scholar] [CrossRef]
- Grayer, R.J.; Veitch, N.C. Flavanones and dihydroflavonols. In Flavonoids; CRC Press: Boca Raton, FL, USA, 2005; pp. 924–1009. [Google Scholar]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, D.; Yang, D.; Zhen, W.; Zhang, J.; Peng, S. The effect of icariin on bone metabolism and its potential clinical application. Osteoporos. Int. 2018, 29, 535–544. [Google Scholar] [CrossRef]
- Prior, R.L.; Lazarus, S.A.; Cao, G.; Muccitelli, H.; Hammerstone, J.F. Identification of procyanidins and anthocyanins in blueberries and cranberries (Vaccinium spp.) using high-performance liquid chromatography/mass spectrometry. J. Agric. Food Chem. 2001, 49, 1270–1276. [Google Scholar] [CrossRef]
- Si, W.; Gong, J.; Tsao, R.; Kalab, M.; Yang, R.; Yin, Y. Bioassay-guided purification and identification of antimicrobial components in Chinese green tea extract. J. Chromatogr. A 2006, 1125, 204–210. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Pan, Q.-H.; Shi, Y.; Duan, C.-Q. Chemical synthesis of proanthocyanidins in vitro and their reactions in aging wines. Molecules (Basel Switzerland) 2008, 13, 3007–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.F.; Taylor, A.W.; Deinzer, M.L. Quantitative analysis of xanthohumol and related prenylflavonoids in hops and beer by liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 1999, 832, 97–107. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Clifford, M.N. Flavanones, chalcones and dihydrochalcones – nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1073–1080. [Google Scholar] [CrossRef]
- Engelhardt, U.H.; Finger, A.; Kuhr, S. Determination of flavone C-glycosides in tea. Z Lebensm Unters 1993, 197, 239–244. [Google Scholar] [CrossRef]
- Carnat, A.; Carnat, A.P.; Fraisse, D.; Ricoux, L.; Lamaison, J.L. The aromatic and polyphenolic composition of Roman camomile tea. Fitoterapia 2004, 75, 32–38. [Google Scholar] [CrossRef]
- Caristi, C.; Bellocco, E.; Gargiulli, C.; Toscano, G.; Leuzzi, U. Flavone-di-C-glycosides in citrus juices from Southern Italy. Food Chem. 2006, 95, 431–437. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Czemerys, R. Antioxidant activity and phenolic compounds in 32 selected herbs. Food Chem. 2007, 105, 940–949. [Google Scholar] [CrossRef]
- Wijaya, G.Y.; Mares, D.J. Apigenin di-C-glycosides (ACG) content and composition in grains of bread wheat (Triticum aestivum) and related species. J. Cereal Sci. 2012, 56, 260–267. [Google Scholar] [CrossRef]
- Pereira-Caro, G.; Cros, G.; Yokota, T.; Crozier, A. Phytochemical profiles of black, red, brown, and white rice from the Camargue region of France. J. Agric. Food Chem. 2013, 61, 7976–7986. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, S.C.Q.; Taveira, M.; Cabrita, A.R.J.; Fonseca, A.J.M.; Valentão, P.; Andrade, P.B. European marketable grain legume seeds: Further insight into phenolic compounds profiles. Food Chem. 2017, 215, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Rimbach, G. Which sources of flavonoids: Complex diets or dietary supplements? Adv. Nutr. 2011, 2, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.M.; Chen, R.Y.; Leung, L.K.; Chan, F.L.; Huang, Y.; Chen, Z.-Y. Difference in flavonoid and isoflavone profile between soybean and soy leaf. Biomed. Pharmacother. 2002, 56, 289–295. [Google Scholar] [CrossRef]
- Zaheer, K.; Humayoun Akhtar, M. An updated review of dietary isoflavones: Nutrition, processing, bioavailability and impacts on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1280–1293. [Google Scholar] [CrossRef]
- Bowles, D.; Isayenkova, J.; Lim, E.K.; Poppenberger, B. Glycosyltransferases: Managers of small molecules. Curr. Opin. Plant. Biol. 2005, 8, 254–263. [Google Scholar] [CrossRef]
- Ferrer, J.L.; Austin, M.B.; Stewart, C. Jr.; Noel, J.P. Structure and function of enzymes involved in the biosynthesis of phenylpropanoids. Plant. Physiol. Biochem. 2008, 46, 356–370. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.S.; Baron, R.; Haanpaa, M.; Kalso, E.; Loeser, J.D.; Rice, A.S.; Treede, R.D. A new definition of neuropathic pain. Pain 2011, 152, 2204–2205. [Google Scholar] [CrossRef]
- O’Connor, A.B. Neuropathic pain: Quality-of-life impact, costs and cost effectiveness of therapy. Pharmacoeconomics 2009, 27, 95–112. [Google Scholar] [CrossRef]
- Langley, P.C.; Van Litsenburg, C.; Cappelleri, J.C.; Carroll, D. The burden associated with neuropathic pain in Western Europe. J. Med. Econ. 2013, 16, 85–95. [Google Scholar] [CrossRef]
- Yawn, B.P.; Wollan, P.C.; Weingarten, T.N.; Watson, J.C.; Hooten, W.M.; Melton, L.J. 3rd, The prevalence of neuropathic pain: Clinical evaluation compared with screening tools in a community population. Pain Med. 2009, 10, 586–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borsook, D. Neurological diseases and pain. Brain 2012, 135, 320–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, J.C.; Sandroni, P. Central Neuropathic Pain Syndromes. Mayo Clin. Proc. 2016, 91, 372–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnerup, N.B.; Haroutounian, S.; Kamerman, P.; Baron, R.; Bennett, D.L.; Bouhassira, D.; Cruccu, G.; Freeman, R.; Hansson, P.; Nurmikko, T.; et al. Neuropathic pain: An updated grading system for research and clinical practice. Pain 2016, 157, 1599–1606. [Google Scholar] [CrossRef] [Green Version]
- Hershman, D.L.; Lacchetti, C.; Dworkin, R.H.; Lavoie Smith, E.M.; Bleeker, J.; Cavaletti, G.; Chauhan, C.; Gavin, P.; Lavino, A.; Lustberg, M.B.; et al. Prevention and management of chemotherapy-induced peripheral neuropathy in survivors of adult cancers: American Society of Clinical Oncology clinical practice guideline. J. Clin. Oncol. 2014, 32, 1941–1967. [Google Scholar] [CrossRef] [Green Version]
- Fallon, M.T. Neuropathic pain in cancer. Br. J. Anaesth 2013, 111, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Banach, M.; Juranek, J.K.; Zygulska, A.L. Chemotherapy-induced neuropathies-a growing problem for patients and health care providers. Brain Behav. 2017, 7, e00558. [Google Scholar] [CrossRef]
- Brown, T.J.; Sedhom, R.; Gupta, A. Chemotherapy-Induced Peripheral Neuropathy. Jama Oncol. 2019, 5, 750. [Google Scholar] [CrossRef] [Green Version]
- Maestri, A.; De Pasquale Ceratti, A.; Cundari, S.; Zanna, C.; Cortesi, E.; Crino, L. A pilot study on the effect of acetyl-L-carnitine in paclitaxel- and cisplatin-induced peripheral neuropathy. Tumori 2005, 91, 135–138. [Google Scholar] [CrossRef]
- Argyriou, A.A.; Cavaletti, G.; Briani, C.; Velasco, R.; Bruna, J.; Campagnolo, M.; Alberti, P.; Bergamo, F.; Cortinovis, D.; Cazzaniga, M.; et al. Clinical pattern and associations of oxaliplatin acute neurotoxicity: A prospective study in 170 patients with colorectal cancer. Cancer 2013, 119, 438–444. [Google Scholar] [CrossRef] [Green Version]
- Argyriou, A.A.; Polychronopoulos, P.; Iconomou, G.; Chroni, E.; Kalofonos, H.P. A review on oxaliplatin-induced peripheral nerve damage. Cancer Treat. Rev. 2008, 34, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.; Arnold, D. Oxaliplatin: A review of approved uses. Expert Opin. Pharmacother. 2012, 13, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, M.I.; Pereira, A.F.; Nogueira, R.B.; Rolim, F.E.; Brito, G.A.; Wong, D.V.; Lima-Junior, R.C.; de Albuquerque Ribeiro, R.; Vale, M.L. The antioxidant effects of the flavonoids rutin and quercetin inhibit oxaliplatin-induced chronic painful peripheral neuropathy. Mol. Pain 2013, 9, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwingel, T.E.; Klein, C.P.; Nicoletti, N.F.; Dora, C.L.; Hadrich, G.; Bica, C.G.; Lopes, T.G.; da Silva, V.D.; Morrone, F.B. Effects of the compounds resveratrol, rutin, quercetin, and quercetin nanoemulsion on oxaliplatin-induced hepatotoxicity and neurotoxicity in mice. Naunyn Schmiedebergs Arch. Pharm. 2014, 387, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Subhan, F.; Ahmad, N.; Sewell, R.D.E. The flavonoid 6-methoxyflavone allays cisplatin-induced neuropathic allodynia and hypoalgesia. Biomed. Pharm. 2017, 95, 1725–1733. [Google Scholar] [CrossRef] [PubMed]
- Chtourou, Y.; Gargouri, B.; Kebieche, M.; Fetoui, H. Naringin Abrogates Cisplatin-Induced Cognitive Deficits and Cholinergic Dysfunction Through the Down-Regulation of AChE Expression and iNOS Signaling Pathways in Hippocampus of Aged Rats. J. Mol. Neurosci. 2015, 56, 349–362. [Google Scholar] [CrossRef]
- Gao, W.; Zan, Y.; Wang, Z.J.; Hu, X.Y.; Huang, F. Quercetin ameliorates paclitaxel-induced neuropathic pain by stabilizing mast cells, and subsequently blocking PKCepsilon-dependent activation of TRPV1. Acta Pharm. Sin. 2016, 37, 1166–1177. [Google Scholar] [CrossRef] [Green Version]
- Nadipelly, J.; Sayeli, V.; Kadhirvelu, P.; Shanmugasundaram, J.; Cheriyan, B.V.; Subramanian, V. Effect of certain trimethoxy flavones on paclitaxel - induced peripheral neuropathy in mice. Integr. Med. Res. 2018, 7, 159–167. [Google Scholar] [CrossRef]
- Sayeli, V.; Nadipelly, J.; Kadhirvelu, P.; Cheriyan, B.V.; Shanmugasundaram, J.; Subramanian, V. Effect of flavonol and its dimethoxy derivatives on paclitaxel-induced peripheral neuropathy in mice. J. Basic Clin. Physiol. Pharmacol. 2018, 29, 525–535. [Google Scholar] [CrossRef]
- Gui, Y.; Zhang, J.; Chen, L.; Duan, S.; Tang, J.; Xu, W.; Li, A. Icariin, a flavonoid with anti-cancer effects, alleviated paclitaxel-induced neuropathic pain in a SIRT1-dependent manner. Mol. Pain 2018, 14, 1744806918768970. [Google Scholar] [CrossRef]
- Shao, H.; Xue, Q.; Zhang, F.; Luo, Y.; Zhu, H.; Zhang, X.; Zhang, H.; Ding, W.; Yu, B. Spinal SIRT1 activation attenuates neuropathic pain in mice. PLoS ONE 2014, 9, e100938. [Google Scholar] [CrossRef]
- Li, S.; Pan, M.-H.; Lo, C.-Y.; Tan, D.; Wang, Y.; Shahidi, F.; Ho, C.-T. Chemistry and health effects of polymethoxyflavones and hydroxylated polymethoxyflavones. J. Funct. Foods 2009, 1, 2–12. [Google Scholar] [CrossRef]
- Boulton, A.J.; Gries, F.A.; Jervell, J.A. Guidelines for the diagnosis and outpatient management of diabetic peripheral neuropathy. Diabet Med. 1998, 15, 508–514. [Google Scholar] [CrossRef]
- Kaur, S.; Pandhi, P.; Dutta, P. Painful diabetic neuropathy: An update. Ann. Neurosci. 2011, 18, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, S.; Boulton, A.J.; Dickenson, A.H. Mechanisms and management of diabetic painful distal symmetrical polyneuropathy. Diabetes Care 2013, 36, 2456–2465. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N. Advanced glycation endproducts--role in pathology of diabetic complications. Diabetes Res. Clin. Pr. 2005, 67, 3–21. [Google Scholar] [CrossRef]
- Toth, C.; Rong, L.L.; Yang, C.; Martinez, J.; Song, F.; Ramji, N.; Brussee, V.; Liu, W.; Durand, J.; Nguyen, M.D.; et al. Receptor for advanced glycation end products (RAGEs) and experimental diabetic neuropathy. Diabetes 2007, 4, 1002–1017. [Google Scholar] [CrossRef] [Green Version]
- Du, X.L.; Edelstein, D.; Rossetti, L.; Fantus, I.G.; Goldberg, H.; Ziyadeh, F.; Wu, J.; Brownlee, M. Hyperglycemia-induced mitochondrial superoxide overproduction activates the hexosamine pathway and induces plasminogen activator inhibitor-1 expression by increasing Sp1 glycosylation. Proc. Natl. Acad. Sci. USA 2000, 97, 12222–12226. [Google Scholar] [CrossRef] [Green Version]
- Yamagishi, S.-I.; Uehara, K.; Otsuki, S.; Yagihashi, S. Differential influence of increased polyol pathway on protein kinase C expressions between endoneurial and epineurial tissues in diabetic mice. J. Neurochem. 2003, 87, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Uehara, K.; Yamagishi, S.-I.; Otsuki, S.; Chin, S.; Yagihashi, S. Effects of Polyol Pathway Hyperactivity on Protein Kinase C Activity, Nociceptive Peptide Expression, and Neuronal Structure in Dorsal Root Ganglia in Diabetic Mice. Diabetes 2004, 53, 3239–3247. [Google Scholar] [CrossRef] [Green Version]
- Cameron, N.E.; Cotter, M.A. Effects of protein kinase Cbeta inhibition on neurovascular dysfunction in diabetic rats: Interaction with oxidative stress and essential fatty acid dysmetabolism. Diabetes Metab. Res. Rev. 2002, 18, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Das Evcimen, N.; King, G.L. The role of protein kinase C activation and the vascular complications of diabetes. Pharm. Res. 2007, 55, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Matsumura, T.; Edelstein, D.; Rossetti, L.; Zsengellér, Z.; Szabó, C.; Brownlee, M. Inhibition of GAPDH activity by poly(ADP-ribose) polymerase activates three major pathways of hyperglycemic damage in endothelial cells. J. Clin. Investig. 2003, 112, 1049–1057. [Google Scholar] [CrossRef] [Green Version]
- Obrosova, I.G.; Julius, U.A. Role for poly(ADP-ribose) polymerase activation in diabetic nephropathy, neuropathy and retinopathy. Curr. Vasc. Pharm. 2005, 3, 267–283. [Google Scholar] [CrossRef] [PubMed]
- Addepalli, V.; Suryavanshi, S.V. Catechin attenuates diabetic autonomic neuropathy in streptozotocin induced diabetic rats. Biomed. Pharm. 2018, 108, 1517–1523. [Google Scholar] [CrossRef]
- Li, M.; Li, Q.; Zhao, Q.; Zhang, J.; Lin, J. Luteolin improves the impaired nerve functions in diabetic neuropathy: Behavioral and biochemical evidences. Int. J. Clin. Exp. Pathol. 2015, 8, 10112–10120. [Google Scholar] [PubMed]
- Mirshekar, M.; Roghani, M.; Khalili, M.; Baluchnejadmojarad, T.; Arab Moazzen, S. Chronic oral pelargonidin alleviates streptozotocin-induced diabetic neuropathic hyperalgesia in rat: Involvement of oxidative stress. Iran. Biomed. J. 2010, 14, 33–39. [Google Scholar]
- Tian, R.; Yang, W.; Xue, Q.; Gao, L.; Huo, J.; Ren, D.; Chen, X. Rutin ameliorates diabetic neuropathy by lowering plasma glucose and decreasing oxidative stress via Nrf2 signaling pathway in rats. Eur. J. Pharm. 2016, 771, 84–92. [Google Scholar] [CrossRef]
- Valsecchi, A.E.; Franchi, S.; Panerai, A.E.; Rossi, A.; Sacerdote, P.; Colleoni, M. The soy isoflavone genistein reverses oxidative and inflammatory state, neuropathic pain, neurotrophic and vasculature deficits in diabetes mouse model. Eur. J. Pharm. 2011, 650, 694–702. [Google Scholar] [CrossRef]
- Feldman, E.L.; Stevens, M.J.; Thomas, P.K.; Brown, M.B.; Canal, N.; Greene, D.A. A practical two-step quantitative clinical and electrophysiological assessment for the diagnosis and staging of diabetic neuropathy. Diabetes Care 1994, 17, 1281–1289. [Google Scholar] [CrossRef]
- Perkins, B.A.; Olaleye, D.; Zinman, B.; Bril, V. Simple screening tests for peripheral neuropathy in the diabetes clinic. Diabetes Care 2001, 24, 250–2506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Rejaie, S.S.; Aleisa, A.M.; Abuohashish, H.M.; Parmar, M.Y.; Ola, M.S.; Al-Hosaini, A.A.; Ahmed, M.M. Naringenin neutralises oxidative stress and nerve growth factor discrepancy in experimental diabetic neuropathy. Neurol. Res. 2015, 37, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Bachewal, P.; Gundu, C.; Yerra, V.G.; Kalvala, A.K.; Areti, A.; Kumar, A. Morin exerts neuroprotection via attenuation of ROS induced oxidative damage and neuroinflammation in experimental diabetic neuropathy. Biofactors 2018, 44, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Kishore, L.; Kaur, N.; Singh, R. Effect of Kaempferol isolated from seeds of Eruca sativa on changes of pain sensitivity in Streptozotocin-induced diabetic neuropathy. Inflammopharmacology 2018, 26, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, C.; Cui, W.G.; Ma, Q.; Zhou, W.H. Fisetin exerts antihyperalgesic effect in a mouse model of neuropathic pain: Engagement of spinal serotonergic system. Sci. Rep. 2015, 5, 9043. [Google Scholar] [CrossRef]
- Cardozo, L.F.; Pedruzzi, L.M.; Stenvinkel, P.; Stockler-Pinto, M.B.; Daleprane, J.B.; Leite, M., Jr.; Mafra, D. Nutritional strategies to modulate inflammation and oxidative stress pathways via activation of the master antioxidant switch Nrf2. Biochimie 2013, 95, 1525–1533. [Google Scholar] [CrossRef]
- Agca, C.A.; Tuzcu, M.; Hayirli, A.; Sahin, K. Taurine ameliorates neuropathy via regulating NF-kappaB and Nrf2/HO-1 signaling cascades in diabetic rats. Food Chem. Toxicol. 2014, 71, 116–121. [Google Scholar] [CrossRef]
- Kumar, A.; Mittal, R. Nrf2: A potential therapeutic target for diabetic neuropathy. Inflammopharmacology 2017, 25, 393–402. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-hydroxy-2’-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, G.; Vinik, A. Nerve growth factor and diabetic neuropathy. Exp. Diabesity Res. 2003, 4, 271–285. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, Y.; Xu, Z.Z.; Wang, X.; Park, J.Y.; Zhuang, Z.Y.; Tan, P.H.; Gao, Y.J.; Roy, K.; Corfas, G.; Lo, E.H.; et al. Distinct roles of matrix metalloproteases in the early- and late-phase development of neuropathic pain. Nat. Med. 2008, 14, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Kuhad, A.; Singh, P.; Chopra, K. Matrix metalloproteinases: Potential therapeutic target for diabetic neuropathic pain. Expert Opin. Targets 2015, 19, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Yong, V.W. Metalloproteinases: Mediators of pathology and regeneration in the CNS. Nat. Rev. Neurosci. 2005, 6, 931–944. [Google Scholar] [CrossRef]
- Naik, A.K.; Tandan, S.K.; Dudhgaonkar, S.P.; Jadhav, S.H.; Kataria, M.; Prakash, V.R.; Kumar, D. Role of oxidative stress in pathophysiology of peripheral neuropathy and modulation by N-acetyl-L-cysteine in rats. Eur. J. Pain 2006, 10, 573–579. [Google Scholar] [CrossRef]
- Liu, W.-T.; Han, Y.; Liu, Y.-P.; Song, A.A.; Barnes, B.; Song, X.-J. Spinal matrix metalloproteinase-9 contributes to physical dependence on morphine in mice. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 7613–7623. [Google Scholar] [CrossRef]
- Pittenger, G.L.; Mehrabyan, A.; Simmons, K.; Dublin, C.; Barlow, P.; Vinik, A.I. Small fiber neuropathy is associated with the metabolic syndrome. Metab. Syndr. Relat. Disord. 2005, 3, 113–121. [Google Scholar] [CrossRef]
- Kandhare, A.D.; Raygude, K.S.; Ghosh, P.; Ghule, A.E.; Bodhankar, S.L. Neuroprotective effect of naringin by modulation of endogenous biomarkers in streptozotocin induced painful diabetic neuropathy. Fitoterapia 2012, 83, 650–659. [Google Scholar] [CrossRef]
- Li, P.; Xiong, D.L.; Sun, W.P.; Xu, S.Y. Effects of baicalin on diabetic neuropathic pain involving transient receptor potential vanilloid 1 in the dorsal root ganglia of rats. Neuroreport 2018, 29, 1492–1498. [Google Scholar] [CrossRef]
- Visnagri, A.; Kandhare, A.D.; Chakravarty, S.; Ghosh, P.; Bodhankar, S.L. Hesperidin, a flavanoglycone attenuates experimental diabetic neuropathy via modulation of cellular and biochemical marker to improve nerve functions. Pharm. Biol. 2014, 52, 814–828. [Google Scholar] [CrossRef]
- Raposo, D.; Morgado, C.; Pereira-Terra, P.; Tavares, I. Nociceptive spinal cord neurons of laminae I–III exhibit oxidative stress damage during diabetic neuropathy which is prevented by early antioxidant treatment with epigallocatechin-gallate (EGCG). Brain Res. Bull. 2015, 110, 68–75. [Google Scholar] [CrossRef]
- Liu, M.; Liao, K.; Yu, C.; Li, X.; Liu, S.; Yang, S. Puerarin alleviates neuropathic pain by inhibiting neuroinflammation in spinal cord. Mediat. Inflamm. 2014, 2014, 485927. [Google Scholar] [CrossRef] [PubMed]
- Dyck, P.J.; Larson, T.; O’Brien, P.; Velosa, J. Patterns of quantitative sensation testing of hypoesthesia and hyperalgesia are predictive of diabetic polyneuropathy: A study of three cohorts. Nerve growth factor study group. Diabetes Care 2000, 23, 510–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freshwater, J.D.; Calcutt, N.A. Low doses of formalin reveal allodynia in diabetic rats. J. Neuropathic Pain Symptom Palliation 2005, 1, 39–46. [Google Scholar] [CrossRef]
- Calcutt, N.; Freshwater, J.; Mizisin, A. Prevention of sensory disorders in diabetic Sprague-Dawley rats by aldose reductase inhibition or treatment with ciliary neurotrophic factor. Diabetologia 2004, 47, 718–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stavniichuk, R.; Drel, V.R.; Shevalye, H.; Maksimchyk, Y.; Kuchmerovska, T.M.; Nadler, J.L.; Obrosova, I.G. Baicalein alleviates diabetic peripheral neuropathy through inhibition of oxidative-nitrosative stress and p38 MAPK activation. Exp. Neurol. 2011, 230, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, A.; Mitsuma, A.; Maeda, O.; Kajiyama, H.; Kiyoi, H.; Kodera, Y.; Nagino, M.; Goto, H.; Ando, Y. Quantitative assessment of chemotherapy-induced peripheral neurotoxicity using a point-of-care nerve conduction device. Cancer Sci. 2016, 107, 1453–1457. [Google Scholar] [CrossRef] [Green Version]
- Cameron, N.E.; Cotter, M.A.; Jack, A.M.; Basso, M.D.; Hohman, T.C. Protein kinase C effects on nerve function, perfusion, Na(+), K(+)-ATPase activity and glutathione content in diabetic rats. Diabetologia 1999, 42, 1120–1130. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Dai, X.; Zhang, Z.; Jiang, Y.; Ma, X.; Cai, X.; Li, Y. Proanthocyanidins protect against early diabetic peripheral neuropathy by modulating endoplasmic reticulum stress. J. Nutr. Biochem. 2014, 25, 765–772. [Google Scholar] [CrossRef]
- Akbar, S.; Subhan, F.; Karim, N.; Shahid, M.; Ahmad, N.; Ali, G.; Mahmood, W.; Fawad, K. 6-Methoxyflavanone attenuates mechanical allodynia and vulvodynia in the streptozotocin-induced diabetic neuropathic pain. Biomed. Pharmacother. 2016, 84, 962–971. [Google Scholar] [CrossRef]
- Colburn, R.W.; Rickman, A.J.; DeLeo, J.A. The effect of site and type of nerve injury on spinal glial activation and neuropathic pain behavior. Exp. Neurol. 1999, 157, 289–304. [Google Scholar] [CrossRef]
- Bennett, G.J.; Xie, Y.-K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. PAIN 1988, 33, 87–107. [Google Scholar] [CrossRef]
- Colleoni, M.; Sacerdote, P. Murine models of human neuropathic pain. Biochim. Biophys. Acta 2010, 1802, 924–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, P.J.; Wu, A.; Moalem-Taylor, G. Chronic constriction of the sciatic nerve and pain hypersensitivity testing in rats. J. Vis. Exp. Jove 2012, 3393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertozzi, M.M.; Rossaneis, A.C.; Fattori, V.; Longhi-Balbinot, D.T.; Freitas, A.; Cunha, F.Q.; Alves-Filho, J.C.; Cunha, T.M.; Casagrande, R.; Verri, W.A., Jr. Diosmin reduces chronic constriction injury-induced neuropathic pain in mice. Chem. Biol. Interact. 2017, 273, 180–189. [Google Scholar] [CrossRef]
- Carballo-Villalobos, A.I.; González-Trujano, M.-E.; Pellicer, F.; López-Muñoz, F.J. Antihyperalgesic Effect of Hesperidin Improves with Diosmin in Experimental Neuropathic Pain. Biomed. Res. Int. 2016, 2016, 8263463. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Bedi, O.; Sharma, N.; Singh, S.; Deshmukh, R.; Kumar, P. Anti-hyperalgesic and anti-nociceptive potentials of standardized grape seed proanthocyanidin extract against CCI-induced neuropathic pain in rats. J. Basic Clin. Physiol. Pharm. 2016, 27, 9–17. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, N.; Zhu, C.; Ma, L.; Yang, J.; Du, J.; Zhang, W.; Sun, T.; Niu, J.; Yu, J. Antinociceptive effect of isoorientin against neuropathic pain induced by the chronic constriction injury of the sciatic nerve in mice. Int. Immunopharmacol. 2019, 75, 105753. [Google Scholar] [CrossRef]
- Komirishetty, P.; Areti, A.; Sistla, R.; Kumar, A. Morin Mitigates Chronic Constriction Injury (CCI)-Induced Peripheral Neuropathy by Inhibiting Oxidative Stress Induced PARP Over-Activation and Neuroinflammation. Neurochem. Res. 2016, 41, 2029–2042. [Google Scholar] [CrossRef]
- Kuang, X.; Huang, Y.; Gu, H.F.; Zu, X.Y.; Zou, W.Y.; Song, Z.B.; Guo, Q.L. Effects of intrathecal epigallocatechin gallate, an inhibitor of Toll-like receptor 4, on chronic neuropathic pain in rats. Eur. J. Pharm. 2012, 676, 51–56. [Google Scholar] [CrossRef]
- Xifró, X.; Vidal-Sancho, L.; Boadas-Vaello, P.; Turrado, C.; Alberch, J.; Puig, T.; Verdú, E. Novel epigallocatechin-3-gallate (EGCG) derivative as a new therapeutic strategy for reducing neuropathic pain after chronic constriction nerve injury in mice. PLoS ONE 2015, 10, e0123122. [Google Scholar] [CrossRef] [Green Version]
- Valsecchi, A.E.; Franchi, S.; Panerai, A.E.; Sacerdote, P.; Trovato, A.E.; Colleoni, M. Genistein, a natural phytoestrogen from soy, relieves neuropathic pain following chronic constriction sciatic nerve injury in mice: Anti-inflammatory and antioxidant activity. J. Neurochem. 2008, 107, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Civi, S.; Emmez, G.; Dere, U.A.; Borcek, A.O.; Emmez, H. Effects of quercetin on chronic constriction nerve injury in an experimental rat model. Acta Neurochir. (Wien.) 2016, 158, 959–965; discussion 965. [Google Scholar]
- Hara, K.; Haranishi, Y.; Terada, T.; Takahashi, Y.; Nakamura, M.; Sata, T. Effects of intrathecal and intracerebroventricular administration of luteolin in a rat neuropathic pain model. Pharm. Biochem. Behav. 2014, 125, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Goecks, C.S.; Horst, A.; Moraes, M.S.; Scheid, T.; Kolberg, C.; Bello-Klein, A.; Partata, W.A. Assessment of oxidative parameters in rat spinal cord after chronic constriction of the sciatic nerve. Neurochem. Res. 2012, 37, 1952–1958. [Google Scholar] [CrossRef]
- Choi, D.C.; Lee, J.Y.; Lim, E.J.; Baik, H.H.; Oh, T.H.; Yune, T.Y. Inhibition of ROS-induced p38MAPK and ERK activation in microglia by acupuncture relieves neuropathic pain after spinal cord injury in rats. Exp. Neurol. 2012, 236, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Tal, M. A novel antioxidant alleviates heat hyperalgesia in rats with an experimental painful peripheral neuropathy. Neuroreport 1996, 7, 1382–1384. [Google Scholar] [CrossRef]
- Khalil, Z.; Liu, T.; Helme, R.D. Free radicals contribute to the reduction in peripheral vascular responses and the maintenance of thermal hyperalgesia in rats with chronic constriction injury. Pain 1999, 79, 31–37. [Google Scholar] [CrossRef]
- Obrosova, I.G.; Li, F.; Abatan, O.I.; Forsell, M.A.; Komjati, K.; Pacher, P.; Szabo, C.; Stevens, M.J. Role of poly(ADP-Ribose) polymerase activation in diabetic neuropathy. Diabetes 2004, 53, 711–720. [Google Scholar] [CrossRef] [Green Version]
- Sommer, C.; Kress, M. Recent findings on how proinflammatory cytokines cause pain: Peripheral mechanisms in inflammatory and neuropathic hyperalgesia. Neurosci. Lett. 2004, 361, 184–187. [Google Scholar] [CrossRef]
- Jagtap, P.; Szabo, C. Poly(ADP-ribose) polymerase and the therapeutic effects of its inhibitors. Nat. Rev. Drug Discov. 2005, 4, 421–440. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.C.; Snyder, S.H. Poly(ADP-ribose) polymerase is a mediator of necrotic cell death by ATP depletion. Proc. Natl. Acad. Sci. USA 1999, 96, 13978–13982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Morioka, N.; Kodama, K.; Tomori, M.; Yoshikawa, K.; Saeki, M.; Nakamura, Y.; Zhang, F.F.; Hisaoka-Nakashima, K.; Nakata, Y. Stimulation of nuclear receptor REV-ERBs suppresses production of pronociceptive molecules in cultured spinal astrocytes and ameliorates mechanical hypersensitivity of inflammatory and neuropathic pain of mice. Brain Behav. Immun. 2019, 78, 116–130. [Google Scholar] [CrossRef]
- Kuiper, G.G.J.M.; Carlsson, B.; Grandien, K.; Enmark, E.; Häggblad, J.; Nilsson, S.; Gustafsson, J.-A.K. Comparison of the Ligand Binding Specificity and Transcript Tissue Distribution of Estrogen Receptors α and β. Endocrinology 1997, 138, 863–870. [Google Scholar] [CrossRef]
- Wu, L.-J.; Zhuo, M. Targeting the NMDA receptor subunit NR2B for the treatment of neuropathic pain. Neurotherapeutics 2009, 6, 693–702. [Google Scholar] [CrossRef] [Green Version]
- Pérez, J.; Ware, M.A.; Chevalier, S.; Gougeon, R.; Shir, Y. Dietary Omega-3 Fatty Acids May Be Associated with Increased Neuropathic Pain in Nerve-Injured Rats. Anesth. Analg. 2005, 101, 444–448. [Google Scholar] [CrossRef]
- Martin, Y.B.; Avendaño, C. Effects of removal of dietary polyunsaturated fatty acids on plasma extravasation and mechanical allodynia in a trigeminal neuropathic pain model. Mol. Pain 2009, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Ko, G.D.; Nowacki, N.B.; Arseneau, L.; Eitel, M.; Hum, A. Omega-3 Fatty Acids for Neuropathic Pain: Case Series. Clin. J. Pain 2010, 26, 168–172. [Google Scholar] [CrossRef]
- Seltzer, Z.E.; Dubner, R.; Shir, Y. A novel behavioral model of neuropathic pain disorders produced in rats by partial sciatic nerve injury. Pain 1990, 43, 205–218. [Google Scholar] [CrossRef]
- Malmberg, A.B.; Basbaum, A.I. Partial sciatic nerve injury in the mouse as a model of neuropathic pain: Behavioral and neuroanatomical correlates. PAIN 1998, 76, 215–222. [Google Scholar] [CrossRef]
- Bura, S.A.; Nadal, X.; Ledent, C.; Maldonado, R.; Valverde, O. A 2A adenosine receptor regulates glia proliferation and pain after peripheral nerve injury. Pain 2008, 140, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Kiguchi, N.; Kobayashi, Y.; Ozaki, M.; Kishioka, S. Pioglitazone attenuates tactile allodynia and thermal hyperalgesia in mice subjected to peripheral nerve injury. J. Pharm. Sci. 2008, 108, 341–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, K.; Motoyama, N.; Kitayama, T.; Morioka, N.; Kifune, K.; Dohi, T. Spinal antiallodynia action of glycine transporter inhibitors in neuropathic pain models in mice. J. Pharm. Exp. 2008, 326, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, M.; Nagatani, Y.; Saitoh, K.; Takasu, K.; Ono, H. Pharmacological assessments of nitric oxide synthase isoforms and downstream diversity of NO signaling in the maintenance of thermal and mechanical hypersensitivity after peripheral nerve injury in mice. Neuropharmacology 2009, 56, 702–708. [Google Scholar] [CrossRef]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef]
- Shields, S.D.; Eckert, W.A., 3rd; Basbaum, A.I. Spared nerve injury model of neuropathic pain in the mouse: A behavioral and anatomic analysis. J. Pain 2003, 4, 465–470. [Google Scholar] [CrossRef]
- Kim, S.H.; Chung, J.M. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain 1992, 50, 355–363. [Google Scholar] [CrossRef]
- Mogil, J.S.; Wilson, S.G.; Bon, K.; Lee, S.E.; Chung, K.; Raber, P.; Pieper, J.O.; Hain, H.S.; Belknap, J.K.; Hubert, L.; et al. Heritability of nociception I: Responses of 11 inbred mouse strains on 12 measures of nociception. Pain 1999, 80, 67–82. [Google Scholar] [CrossRef]
- Kiso, T.; Watabiki, T.; Tsukamoto, M.; Okabe, M.; Kagami, M.; Nishimura, K.; Aoki, T.; Matsuoka, N. Pharmacological characterization and gene expression profiling of an L5/L6 spinal nerve ligation model for neuropathic pain in mice. Neuroscience 2008, 153, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Cherng, C.-H.; Lee, K.-C.; Chien, C.-C.; Chou, K.-Y.; Cheng, Y.-C.; Hsin, S.-T.; Lee, S.-O.; Shen, C.-H.; Tsai, R.-Y.; Wong, C.-S. Baicalin ameliorates neuropathic pain by suppressing HDAC1 expression in the spinal cord of spinal nerve ligation rats. J. Formos. Med. Assoc. 2014, 113, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.I.; Kim, W.M.; Lee, H.G.; Kim, Y.O.; Yoon, M.H. Role of neuronal nitric oxide synthase in the antiallodynic effects of intrathecal EGCG in a neuropathic pain rat model. Neurosci. Lett. 2012, 510, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Hagenacker, T.; Hillebrand, I.; Wissmann, A.; Büsselberg, D.; Schäfers, M. Anti-allodynic effect of the flavonoid myricetin in a rat model of neuropathic pain: Involvement of p38 and protein kinase C mediated modulation of Ca2+ channels. Eur. J. Pain 2010, 14, 992–998. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Xu, Y.; Han, F.; Sun, D.; Zhang, H.; Li, X.; Yao, X.; Wang, H. Quercetin alleviates thermal and cold hyperalgesia in a rat neuropathic pain model by inhibiting Toll-like receptor signaling. Biomed. Pharm. 2017, 94, 652–658. [Google Scholar] [CrossRef]
- Lu, Y.; Jiang, B.C.; Cao, D.L.; Zhang, Z.J.; Zhang, X.; Ji, R.R.; Gao, Y.J. TRAF6 upregulation in spinal astrocytes maintains neuropathic pain by integrating TNF-alpha and IL-1beta signaling. Pain 2014, 155, 2618–2629. [Google Scholar] [CrossRef] [Green Version]
- Haberland, M.; Montgomery, R.L.; Olson, E.N. The many roles of histone deacetylases in development and physiology: Implications for disease and therapy. Nat. Rev. Genet. 2009, 10, 32–42. [Google Scholar] [CrossRef]
- Bai, G.; Wei, D.; Zou, S.; Ren, K.; Dubner, R. Inhibition of class II histone deacetylases in the spinal cord attenuates inflammatory hyperalgesia. Mol. Pain 2010, 6, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiechio, S.; Copani, A.; Zammataro, M.; Battaglia, G.; Gereau, R.W.T.; Nicoletti, F. Transcriptional regulation of type-2 metabotropic glutamate receptors: An epigenetic path to novel treatments for chronic pain. Trends Pharm. Sci. 2010, 31, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Yaster, M.; Raja, S.N.; Tao, Y.X. Genetic knockout and pharmacologic inhibition of neuronal nitric oxide synthase attenuate nerve injury-induced mechanical hypersensitivity in mice. Mol. Pain 2007, 3, 29. [Google Scholar] [CrossRef] [Green Version]
- Knowles, R.G.; Moncada, S. Nitric oxide synthases in mammals. Biochem. J. 1994, 298, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Sung, C.S.; Wen, Z.H.; Chang, W.K.; Ho, S.T.; Tsai, S.K.; Chang, Y.C.; Wong, C.S. Intrathecal interleukin-1beta administration induces thermal hyperalgesia by activating inducible nitric oxide synthase expression in the rat spinal cord. Brain Res. 2004, 1015, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Bryan, N.S.; Bian, K.; Murad, F. Discovery of the nitric oxide signaling pathway and targets for drug development. Front. Biosci. (Landmark Ed.) 2009, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Aswar, M.; Kute, P.; Mahajan, S.; Mahajan, U.; Nerurkar, G.; Aswar, U. Protective effect of hesperetin in rat model of partial sciatic nerve ligation induced painful neuropathic pain: An evidence of anti-inflammatory and anti-oxidative activity. Pharmacol. Biochem. Behav. 2014, 124, 101–107. [Google Scholar] [CrossRef]
- Muto, N.; Matsuoka, Y.; Arakawa, K.; Kurita, M.; Omiya, H.; Taniguchi, A.; Kaku, R.; Morimatsu, H. Quercetin Attenuates Neuropathic Pain in Rats with Spared Nerve Injury. Acta Med. Okayama 2018, 72, 457–465. [Google Scholar]
- Vaya, J.; Tavori, H.; Khatib, S. Structure-Activity Relationships of Flavonoids. Curr. Org. Chem. 2011, 15, 2641–2657. [Google Scholar]
- Modak, B.; Contreras, M.L.; Gonzalez-Nilo, F.; Torres, R. Structure-antioxidant activity relationships of flavonoids isolated from the resinous exudate of Heliotropium sinuatum. Bioorg. Med. Chem. Lett. 2005, 15, 309–312. [Google Scholar] [CrossRef]
- Lin, C.-Z.; Zhu, C.-C.; Hu, M.; Wu, A.-Z.; Bairu, Z.-D.; Kangsa, S.-Q. Structure-activity relationships of antioxidant activity in vitro about flavonoids isolated from Pyrethrum tatsienense. J. Intercult Ethnopharmacol. 2014, 3, 123–127. [Google Scholar] [CrossRef]
- Lee, S.K.; Nam, K.A.; Hoe, Y.H.; Min, H.Y.; Kim, E.Y.; Ko, H.; Song, S.; Lee, T.; Kim, S. Synthesis and evaluation of cytotoxicity of stilbene analogues. Arch. Pharm. Res. 2003, 26, 253–257. [Google Scholar] [CrossRef]
- Basu, P.; Maier, C. Phytoestrogens and breast cancer: In vitro anticancer activities of isoflavones, lignans, coumestans, stilbenes and their analogs and derivatives. Biomed. Pharmacother. 2018, 107, 1648–1666. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |


{kind=link}
| Anthocyanins | ||
 |  |  |
| Cyanidin | Delphinidin | Pelargonidin |
 |  |  |
| Peonidin | Petunidin | Malvidin |
| Chalcones | ||
 |  |  |
| Arbutin | Chalconaringenin | Phloretin |
 |  | |
| Phloridzin | Xanthohumol | |
| Flavanones | ||
 |  |  |
| Eriodictyol | Hesperetin | Hesperidin |
 |  |  |
| Naringenin | Naringin | 6-methoxyflavanone |
| Flavones | ||
 |  |  |
| Apigenin | Baicalein | Baicalin |
 |  |  |
| Diosmin | Isoorientin | Luteolin |
 |  |  |
| Nobiletin | Tangeretin | 7,2′,3′-trimethoxy flavone |
 |  |  |
| 7,2′,4′-trimethoxy flavone | 7,3′,4′-trimethoxy flavone | 7,5,4′-trimethoxy flavone |
 | ||
| 6-methoxyflavone | ||
| Flavonols | ||
 |  |  |
| Fisetin | Icariin | Isorhamnetin |
 |  |  |
| Kaempferol | Morin | Myricetin |
 |  |  |
| Quercetin | Rutin | 3′,4′-dimethoxy flavonol |
 |  |  |
| 6,3′-dimethoxy flavonol | 7,2′-dimethoxy flavonol | 7,3′-dimethoxy flavonol |
 | ||
| Flavonol | ||
| Flavan-3-ol | ||
 |  |  |
| Catechin | (−)-epicatechin | Epigallocatechin gallate |
 | ||
| Proanthocyanidins | ||
| Isoflavones | ||
 |  |  |
| Biochanin A | Daidzein | Formononetin |
 |  |  |
| Genistein | Glycitein | Puerarin |
| Flavonoid Subgroup | Example | Dietary Sources | References |
|---|---|---|---|
| Anthocyanins | Cyanidin | Edible red (red clover, red hibiscus, red pineapple sage, pink blossom); blue (blue chicory, blue rosemary, cornflower); purple (purple mint, purple passionflower, purple sage); berries, blackcurrants, black carrot, purple potato, red cabbage | Khoo et al. [25] |
| Delphinidin | |||
| Pelargonidin | |||
| Peonidin | |||
| Petunidin | |||
| Malvidin | |||
| Chalcones | Arbutin | Hops, hop containing beers and herbal teas | Stevens et al. [38]; Zhao et al. [29] |
| Chalconaringenin | |||
| Phloretin | |||
| Phloridzin | |||
| Xanthohumol | |||
| Flavanones | Eriodictyol | Citrus fruits like lemon, lime mandarin, orange; grapefruit, herbal tea (Honeybush tea), potato | Tomás-Barberán & Clifford [39] |
| Hesperetin | |||
| Hesperidin | |||
| Naringenin | |||
| Flavones | Apigenin | Dry herbs and teas (Roman Chamomile flowers, tansy leaf, fenugreek seed, rosemary, sage, black tea, green tea, oolong tea); juices and wines (bergamot juice, mandarin orange juice, citron juice, orange juice); fruits, vegetables, olive oil and honey (kiwi, spinach, parsley, celery, lettuce, artichoke, broccoli, watermelon, pumpkin, peas); cereals and legumes (chickpea, fava pea, field pea, wheat grain, black, brown, red and white rice) | Engelhardt et al., 1993 [40]; Carnat et al., 2004 [41]; Caristi et al., 2006 [42]; Wojdylo et al., 2007 [43]; Wijaya & Mares, 2012 [44]; Pereira-Caro et al., 2013 [45]; Magalhães et al. [46] |
| Baicalein | |||
| Diosmin | |||
| Isoorientin | |||
| Luteolin | |||
| Nobiletin | |||
| Tangeretin | |||
| Flavonols | Fisetin | Fruits (apples, berries, grapes), vegetables (curly kale, leek, lettuce, onions, tomatoes), tea, red wine | Egert & Rimbach [47] |
| Isorhamnetin | |||
| Kaempferol | |||
| Morin | |||
| Myricetin | |||
| Quercetin | |||
| Flavan-3-ol | Catechin | Dietary supplements, beverages, whole and processed foods | Prior et al. [34]; Si et al. [35] |
| (−)-Epicatechin | |||
| Epigallocatechin gallate | |||
| Isoflavones | Biochanin A | Kidney beans, lentils, mung bean sprouts, mung beans, red clover, soy products, soybeans, soy products | Ho et al. [48]; Zaheer and Akhtar [49] |
| Daidzein | |||
| Formononetin | |||
| Genistein | |||
| Glycitein | |||
| Puerarin |
| Flavonoids | Animals | Dose mg/kg (Route of Administration) | Effects/Mechanisms of Action | Reference | |
|---|---|---|---|---|---|
| Behavioral Evaluation | Biochemical/Molecular Parameters | ||||
| Icariin | Male Sprague Dawley rats | 25, 50, and 100 mg/kg | ↓ Paclitaxel-induced mechanical allodynia in long term | ↓ Paclitaxel-induced increase of TNF-α, IL-1β, and IL-6, astrocytes, NF-κB (p65) phosphorylation in spinal cord | Gui et al. [74] |
| Intrathecal | Reversed paclitaxel-induced downregulation of SIRT1 expression and H4 acetylation | ||||
| 7,2′,3′-trimethoxy flavone, 7,2′,4′-trimethoxy flavone, 7,3′,4′-trimethoxy flavone and 7,5,4′-trimethoxy flavone | Swiss albino mice Either sex | 25, 50, 100 and 200 mg/kg | ↓ Paclitaxel-induced tactile allodynia, cold allodynia and thermal hyperalgesia | X Proinflammatory cytokines (TNFα, IL-1β) and free radicals (DPPH, NO) | Nadipelly et al. [72] |
| Subcutaneous | |||||
| 20, 30, 60, 120, 240 µM—in vitro | |||||
| Flavonol, 3′,4′-dimethoxy flavonol, 6,3′-dimethoxy flavonol, 7,2′-dimethoxy flavonol and 7,3′-dimethoxy flavonol | Male Swiss albino mice | 25,50, 100, and 200 mg/kg | ↓ Tactile allodynia, cold allodynia and thermal hyperalgesia | X TNFα, IL-1β, DPPH, NO | Sayeli et al. [73] |
| Subcutaneous | |||||
| 20, 30, 60, 120, 240 µM—in vitro | |||||
| 6-methoxyflavone | Male Sprague-Dawley rats | 25, 50 and 75 mg/kg | ↓ Cisplatin-induced mechanical allodynia and heat hypoalgesia | - | Shahid et al. [69] |
| Intraperitoneal | Elicited no detectable deficit in motor control | ||||
| Quercetin | Male Sprague-Dawley rats and mice | 20 and 60 mg/kg | ↑ Heat hyperalgesia and mechanical allodynia in paclitaxel-treated rats and mice | ↓ Expressions of PKCε and TRPV1 in spinal cords and DRGs of paclitaxel-treated rats and mice | Gao et al. [71] |
| Intraperitoneal | X Translocation of PKCε from cytoplasm to membrane in spinal cord and DRG in paclitaxel-treated rats and mice | ||||
| 3, 10, 30 μM/L and 20 and 60 μM/L—in vitro | ↓ Histamine release in RBL-2H3 cells in vitro as well as in plasma of quercetin-treated rats | ||||
| Naringin | Wistar rats. Sex not specified | 25, 50, and 100 mg/kg | Cisplatin with naringin prevented behavioral impairment observed in only cisplatin treated group | X Cisplatin-induced increase in acetylcholinesterase | Chtourou et al. [70] |
| ↓ Na+, K+-ATPase, Ca2+-ATPase, and Mg2+-ATPase activities | |||||
| Oral gavage | X Cisplatin-induced anxiogenic effect in elevated T-maze test | Altered oxidative biomarkers, antioxidant enzymes, nonenzymatic antioxidant, increase in ROS, iNOS mRNA expression, and NO levels in hippocampus | |||
| Quercetin, quercetin nanoemulsion, and rutin | Male BALB/c mice | Quercetin, quercetin nanoemulsion, and rutin (20 mg/kg) | ↓ Oxaliplatin-induced mechanical allodynia | ↓ Nociceptive biomarker c-Fos in dorsal horn of spinal cord | Schwingel et al. [68] |
| Oral gavage | |||||
| Quercetin and rutin | Male Swiss mice | Rutin and quercetin (25, 50, and 100 mg/kg) | X Oxaliplatin-induced peripheral neuropathy | X Lipid peroxidation, tyrosine nitrosylation, and peroxynitrite-associated neuronal damage | Azevedo et al. [67] |
| Intraperitoneal | |||||
| Flavonoids | Animals | Flavonoids (Dose mg/kg and Route of Administration) | Effects/Mechanisms of Action | Reference | ||
|---|---|---|---|---|---|---|
| Behavioral Evaluation/Other Diabetic Parameters | Electrophysiological/Functional Evaluation | Histopathological/Biochemical/Molecular Parameters | ||||
| Catechin | Male Sprague Dawley rats | 25 mg/kg and 50 mg/kg | ↑ Body weight compared to diabetic animals | - | Improved hemodynamic parameters (heart rate, mean atrial pressure and left ventricular systolic pressure), oxidative stress parameters (MDA, GSH, CAT, SOD) | Addepalli et al. [89] |
| Intraperitoneal | ↓ Heart hypertrophy, plasma glucose levels | Reversed diabetes-induced neuronal damage and reduced circulatory MMP-9 | ||||
| Morin | Male Sprague-Dawley rats | 50 and 100 mg/kg | ↓ Mechanical hyperalgesia and mechanical allodynia | Improved measurement of MNCV, SNCV, and nerve blood flow (NBF) | ↑ Mitochondrial-specific superoxide dismutase 2 (SOD2) expression in high glucose-treated N2A cells | Bachewal et al. [97] |
| Oral gavage | ↓ Glucose-induced ROS generation by increasing expression of Nrf2 and its downstream effectors NQO1 and HO-1 in N2A cells | |||||
| 10 and 20 µM—In vitro | ↓ IKK (ser176/180) phosphorylation, levels of TNFα and IL-6 | |||||
| X Translocation and expression of NF-κB in N2A cells | ||||||
| ↓ Levels of TNFα and IL-6 | ||||||
| Grape seed proanthocyanidins (GSPs) and its metabolites C (+)-catechin; EC, (−)-epicatechin | Male Sprague-Dawley rats | 250 mg/kg | GSPs - Improved diabetic parameters, especially low-density lipoprotein level | GSPs—↑ Nerve conduction velocity (NCV) in sciatic/tibial nerves | GSPs—↑ Normal mitochondria, endoplasmic reticulum in sciatic nerves and partially improved myelin sheath morphology | Ding et al. [122] |
| Oral | GSPs—↓ Free Ca2+ concentrations and ER stress markers (GRP78, CHOP, phospho-JNK, total JNK and cleaved caspase-12) | |||||
| (+)-catechin; EC, (−)-epicatechin | GSPs treated cells showed similar cell viability, LDH release extent, apoptosis/necrosis cell fractions to treatment with serum treated from healthy rats | |||||
| 2.5, 5, 10 µM | C and EC—Partially ameliorated cell injury in cells treated with serum from diabetic rats | |||||
| C and EC—X Cell injury, Ca2+ overload and ER stress | ||||||
| Kaempferol | Male Wistar rats | 5 and 10 mg/kg | ↓ Blood glucose level at the end of the study (90 days) | ↑ MNCV compared to diabetic control rats | ↑ Levels of GSH, SOD, and thiobarbituric acid reactive substances (TBARS) | Kishore et al. [98] |
| ↑ Diabetes-induced thermal and mechanical hyperalgesia | ↓ NO level, sciatic AGEs, TNF-α, TGF-β and IL-1β | |||||
| Baicalin | Male Sprague-Dawley rats | 10, 20, and 40 µg/kg | ↓ Diabetes-induced mechanical allodynia and thermal hyperalgesia | - | ↓ Both mRNA and protein expressions of TRPV1 in DRG of diabetic rats | Li et al. [112] |
| Intraperitoneal | ||||||
| 6-Methoxyflavanone | Female Sprague-Dawley rats and BALB/c mice | 10 and 30 mg/kg | No acute toxicity in animals ascertained by a lack of cyanosis, ataxia, convulsions, writhing or mortality | - | Thermal antinociception was antagonized by opioid receptor antagonist naloxone and GABA antagonist pentylenetetrazole | Akbar et al. [123] |
| Intraperitoneal | ↓ Thermal nociception in streptozotocin-induced diabetic neuropathy model at 30- and 60-min post-treatment | |||||
| Elicited anti-allodynic and anti-vulvodynic effects | ||||||
| Rutin | Male Sprague-Dawley rats | 5, 25, and 50 mg/kg | ↓ Plasma glucose level | ↑ MNCV and SNCV in diabetic rats | ↑ Na+, K+-ATPase activities in sciatic nerves | Tian et al. [92] |
| ↓ Caspase-3 expression in DRG neurons | ||||||
| ↓ MDA and ROS levels | ||||||
| Partially increased antioxidant enzymes SOD, GPx, glutathione-S-transferase (GST), and CAT in sciatic nerves | ||||||
| ↑ H2S, Nrf2 and HO-1 in DRG neurons | ||||||
| Intraperitoneal | ↓ Diabetes-induced mechanical hyperalgesia, thermal hyperalgesia, and cold allodynia | ↓ NF-κB, IкBα, p-IкBα, IL-6 and TNF-α in DRG neurons of diabetic rats | ||||
| Naringenin | Male Wister albino | 25 and 50 mg/kg/day | X Fasting blood glucose level and high dose of naringenin increased insulin level | - | ↓ TNFα, IL-1β and IL-6, NO level | Al-Rejaie et al. [96] |
| ↓ Elevated TBARS in sciatic nerves | ||||||
| ↑ GSH, SOD, CAT, GPx and GR levels in sciatic nerves | ||||||
| Improved decreased sciatic expressions of insulin growth factor and NGF levels in sciatic nerves | ||||||
| Intraperitoneal | Improved mechanical and thermal hyperalgesia by increasing tail and paw withdrawal latency time | In histological analyses, low dose—partial focal peripheral axonal loss and regenerating thin myelinated axons, indicative of mild degenerative and regenerative neuropathy high dose—minimal axonal degenerative changes without regenerative features, indicative of minor degenerative neuropathy | ||||
| Luteolin | Male Sprague-Dawley rats | 50 mg/kg, 100 mg/kg and 200 mg/kg | ↓ Plasma glucose levels | Improved nerve function by increasing nerve blood flow (NBF) and nerve conduction velocity (NCV) | ↓ ROS and MDA levels | Li et al. [90] |
| Intraperitoneal | ↓ Diabetes-induced cold allodynia and mechanical and thermal hyperalgesia | ↑ Antioxidant enzymes SOD, GST, GPx and CAT along with Nrf2 and HO-1 in nerve tissues in diabetic rats | ||||
| Epigallocatechin-gallate (EGCG) | Male Wistar rats | 2 g/L | Did not affect blood glucose concentration, body weight or liquid intake compared to diabetic animals | - | X Increase of (8-hydroxy-2-deoxyguanosine (8-OHdG) immunoreaction and Fos expression in spinal cord | Raposo et al. [114] |
| Oral gavage | Ameliorated diabetes-induced tactile allodynia and mechanical hyperalgesia | X Higher percentage of 8-OHdG-IR cells that co-localized with Fos | ||||
| Fisetin | Male C57BL/6J mice | 10 mg/kg | ↑ Body weight and slightly decreased food/water intake | - | ↓ Exacerbated oxidative stress by reducing lipid peroxide, ROS production | Zhao et al. [99] |
| ↑ Increased CAT activity in spinal cord, DRG, and sciatic nerve | ||||||
| Co-administration of ROS donor tert-butyl hydroperoxide(t-BOOH) abrogated antinociceptive activity | ||||||
| Co-administration of ROS scavenger phenyl-N-tert-butylnitrone potentiated antinociceptive activity | ||||||
| Oral gavage | Ameliorated diabetes-induced thermal hyperalgesia and mechanical allodynia | Intrathecal administration of GABAA receptor antagonist bicuculline attenuated antinociceptive activity although or GABAB receptor antagonist phaclofen did not alter antinociceptive activity | ||||
| Puerarin | Male Sprague-Dawley rats | 4, 20, and 100 nM | Did not affect mechanical withdrawal threshold | - | ↓ NF-κB, IL-6, IL-1β, and TNF-α in spinal cord | Liu et al. [115] |
| X Activation of microglia and astroglia in spinal cord | ||||||
| Intrathecal | ↓ Diabetes-induced mechanical allodynia | ↓ Diabetes-induced elevation of TNF-α, IL-1β, and IL-6 and NF-κB DNA binding activities | ||||
| X Overexpression of NF-κB p65 and p65 nucleus translocation | ||||||
| Hesperidin | Sprague Dawley rats | 25, 50 and 100 mg/kg | X Body weight loss, increased plasma glucose level, elevated intake of food and water and urine output | ↑ MNCV and SNCV compared to diabetic rats | ↓ Serum glucosuria, cholesterol, and triglyceride levels | Visnagri et al. [113] |
| Sex not specified | Oral gavage | ↓ Elevated glycated hemoglobin and aldose reductase levels, hemodynamic parameters (SBP, DBP, and MABP, neural lipid peroxidase, NO, and total calcium levels) | ||||
| ↑ Serum insulin, neural SOD, glutathione, and Na+K+ATPase levels | ||||||
| ↑ Plasma glucose level compared to diabetic rats | ↓ mRNA expressions of TNF-α and IL-1β | |||||
| ↑ Diabetes-induced mechano-tactile allodynia and thermal hyperalgesia | Restored diabetes-induced histological aberrations by reducing infiltration of neutrophil and macrophages | |||||
| Naringin | Male Wistar rats | 40 and 80 mg/kg | Ameliorated decreased body weights and increased plasma glucose level | ↑ MNCV | ↑ SOD level | Kandhare et al. [111] |
| ↓ Diabetes-induced increase in food intake, water intake, and urine output | ||||||
| Intraperitoneal | ↓ Decrease in diabetes-induced mechano-tactile allodynia, mechanical hyperalgesia, and thermal hyperalgesia | ↓ TNFα, lipid peroxide, elevated neural nitrite, Na-K-ATPase levels along with percentage of apoptosis | ||||
| Baicalein | C57Bl6/J mice | 30 mg/kg | ↓ Weight gain | Alleviated MNCV and SNCV deficits in diabetic mice | ↓ Diabetes-associated nitrated protein accumulation in sciatic nerve and normalized this variable in spinal cord | Stavniichuk et al. [119] |
| Did not affect non-fasting glycemia | ↓ 12(S) hydroxyeicosatetraenoic acid concentrations but did not alter sciatic nerve and spinal cord LO overexpression | |||||
| Sex not specified | Intraperitoneal | Ameliorated thermal hypoalgesia and tactile allodynia in diabetic mice | Normalized sciatic nerve phosphorylated p38 MAPK expression without affecting total p38 MAPK expression | |||
| Genistein | Male C57BL/6J mice | 3 and 6 mg/kg | Did not alter blood glucose concentrations or body weight decrease or decrease hyperglycemia | - | ↓ Pro-inflammatory cytokines TNFα, IL-1β and IL-6, ROS levels in sciatic nerves; MDA and ROS levels in brain and liver; iNOS in thoracic aorta | Valsecchi et al. [93] |
| ↑ NGF, eNOS and SOD | ||||||
| Did not modify decreased cerebral activities of CAT and GPx | ||||||
| Restored hepatic GPx activity but it did not modify CAT activity decrease | ||||||
| Subcutaneous | ↑ Diabetes-induced mechanical allodynia | Restored the GSH content and the GSH and GSSG ratio in liver but did not modify total glutathione content | ||||
| Pelargonidin | Male Albino Wistar rats | 10 mg/kg | Administration for 8 weeks prevented weight loss and reduced serum glucose level | - | ↓ Increased MDA content and nitrite levels | Mirshekar et al. [91] |
| ↓ Serum glucose level | ↑ Increased SOD level | |||||
| Oral gavage | Ameliorated thermal hyperalgesia by increasing tail-flick response latency | |||||
| Flavonoids | Animals | Flavonoids (Dose mg/kg and Route of Administration) | Effects/Mechanisms of Action | Reference | ||
|---|---|---|---|---|---|---|
| Behavioral Evaluation/Other CCI-Induced Parameters | Electrophysiological/Functional Evaluation | Histopathological/Biochemical/Molecular Parameters | ||||
| Isoorientin | Male pathogen-free Institute of Cancer Research (ICR) mice | 7.5, 15, and 30 mg/kg | ↓ CCI-induced mechanical and cold allodynia and thermal hyperalgesia | Restored CCI-induced SNCV and SNAP | ↑ Levels of total antioxidant capacity (T-AOC), total superoxide dismutase (T-SOD), CAT | Zhang et al. [131] |
| Intragastrical | ↓ MDA concentrations | |||||
| X MMP-9, astrocyte, microglia overexpression in spinal cord | ||||||
| ↓ Protein expressions of TNF-α, IL-6, and IL-1β in spinal cord | ||||||
| Ameliorated CCI-induced histopathological changes, such as disordered myelinated nerve fibers, swollen axons, and neuron gaps, abnormal ultrastructure of sciatic nerve and reduced abnormal myelin sheath percentage | ||||||
| Diosmin | Male Swiss mice | 1, 10 mg/kg | X CCI-induced mechanical and thermal hyperalgesia by NO/cGMP/PKG/KATP channel signaling pathway | - | Single treatment—X mRNA expressions of spinal cord cytokine (IL-1β, IL-33, St2) | Bertozzi et al. [128] |
| Prolonged treatment—↓ TNFα mRNA expression in spinal cord | ||||||
| Intraperitoneal | Single treatment—X Glial cells activation microglia (Iba-1), oligodendrocytes (Olig2) | |||||
| Prolonged treatment—X (Gfap), Iba-1, and Olig2 mRNA expressions | ||||||
| Diosmin and Hesperidin | Male Wistar rats | Hesperidin (10, 100, 316.2, 562.3, 1000 mg/kg), Diosmin (10, 100 mg/kg) | Hesperidin—↓ Mechanical and thermal hyperalgesia | - | Combined antihyperalgesic activity mediated by D2, GABAA, and opioids receptors, but not 5-HT1A receptor | Carballo-Villalobos et al. [129] |
| Intraperitoneal | Hesperidin + Diosmin − Improved antihyperalgesic activity | |||||
| Quercetin | Male Wistar rats | 100 mg/kg | Alleviated mechanical and thermal hypersensitivity higher than morphine and gabapentin | - | - | Çivi et al. [136] |
| Pre-injury administration showed long-term effects on mechanical hypersensitivity | ||||||
| Grape seed Proanthocyanidins (GSPE) | Wistar rats | 100 and 200 mg/kg | ↓ Mechanical and thermal hyperalgesia | - | ↓ MDA and nitrate levels in sciatic nerves | Kaur et al. [130] |
| Co-treatment of GSPE and morphine attenuated morphine tolerance, enhanced antihyperalgesic activity | ||||||
| Either sex | Oral gavage | ↑ GSH level, SOD, and CAT compared to GSPE-alone- and morphine-alone treatments | ||||
| Morin | Male Sprague-Dawley rats | 15 and 30 mg/kg | Improved CCI-induced thermal hyperalgesia, mechanical and cold allodynia | Improved SFI but did not completely recover to normal SFI | Restored levels of GSH, ATP | Komirishett et al. [132] |
| ↓ Nitrite levels in spinal cord | ||||||
| Oral gavage | ↓ Spontaneous pain, corrected foot deformity | ↓ Inflammatory markers (PARP, iNOS, COX-2, NF-κB and phospho-NF-κB, TNF-α and IL-6) in spinal cord | ||||
| ↓ poly (ADP) ribose (PAR) and NF-κB levels | ||||||
| EGCG and its two synthetic derivative compounds 23 and 30 | Female Balb-c mice | 10, 30, 50 and 100 mg/kg | EGCG and compound 30 but not compound 23—↓ CCI-induced thermal hyperalgesia | - | EGCG and compound 30 but not compound 23—↓ FASN in dorsal horn of spinal cord | Xifró et al. [134] |
| Intraperitoneal | EGCG and compounds 23 and 30—No effects on FASN protein expression | |||||
| EGCG and compound 30 but not compound 23—↓ mRNA and protein expressions of TNF-α, IL-1β, IL-6 in dorsal horn of spinal cord | ||||||
| EGCG and compound 30 but not compound 23—↓ NF-κB protein expression in dorsal horn of spinal cord | ||||||
| Compound 30 but not EGCG and compound 23—↓ Phosphorylation and protein expression of NMDAR receptor subunit NMDAR2B in dorsal horn of spinal cord | ||||||
| Fisetin | Male C57BL/6J mice | 5, 15 and 45 mg/kg | ↓ CCI-induced thermal hyperalgesia but not nociceptive sensitivity to mechanical stimuli | - | ↓ Escalated MAO-A to level like non-injured animals but did not affect MAO-B in sham or CCI mice | Zhao et al. [99] |
| Oral gavage | ↓ CCI-induced co-morbid depressive and anxiety-like behaviors | Exhibited antinociceptive activity with involvement of serotonergic system (coupled with 5-HT7) | ||||
| Luteolin | Male Sprague-Dawley rats | 0.1–1.5 mg | Spinal administration reduced cold and mechanical, but not thermal hyperalgesia by activating GABAA receptors in a flumazenil-insensitive manner and µ-opioid receptor | - | - | Hara et al. [137] |
| Intrathecal or intracerebroventricular | Supraspinal administration showed no antihyperalgesic activity | |||||
| High concentration inhibited motor function | ||||||
| Puerarin | Male Sprague-Dawley rats | 4, 20, and 100 nM | ↓ CCI-induced mechanical allodynia | - | X Activation of microglia and astroglia in spinal dorsal horn | Liu et al. [115] |
| Intrathecal | ↓ TNF-α, IL-1β, IL-6, DNA binding activities, overexpression of NF-κB as well as nuclear translocation of p65 | |||||
| EGCG | Male Sprague-Dawley rats | 1 mg/kg | Improved CCI-induced mechanical allodynia and thermal hyperalgesia | - | ↓ IL-1β and TNF-α | Kuang et al. [133] |
| ↑ Anti-inflammatory cytokine (IL-10) | ||||||
| Intrathecal | ↓ mRNA and protein expressions of TLR4 and HMGB1 | |||||
| ↓ NF-κB expression in lumbar spinal dorsal horn | ||||||
| Genistein | Male C57BL/6J mice | 1, 3, 7.5, 15, and 30 mg/kg | Reversed CCI-induced thermal hyperalgesia and mechanical allodynia | - | ↓ mRNA expressions of both IL-1β and IL-6 in sciatic nerve and protein expression of IL-1β in DRG and spinal cord | Valsecchi et al. [135] |
| Subcutaneous | ↓ ROS and MDA levels | |||||
| ↑ GPX and CAT activities in CCI operated animals | ||||||
| X NF-κB activation but did not modify NF transcription in spinal cord | ||||||
| Normalized nerved injury-induced increase of iNOS and nNOS | ||||||
| Flavonoids | Animals | Flavonoids (Dose mg/kg and Route of Administration) | Effects/Mechanisms of Action | Reference | ||
|---|---|---|---|---|---|---|
| Behavioral Evaluation | Electrophysiological/Functional Evaluation | Biochemical/Molecular Parameters | ||||
| SNI | ||||||
| Quercetin | Male Sprague-Dawley rats | 0.1, 1% | Pre-surgery administration—↓ Mechanical allodynia | - | X GFAP in satellite glial cells of ipsilateral L5 DRG | Muto et al. [179] |
| Oral gavage | Post-surgery administration—Did not affect SNI-induced pain | |||||
| SNL | ||||||
| Quercetin | Sprague Dawley rats | 10–100 mg/kg | Single or continuous administration—↓ SNL-induced thermal and cold hyperalgesia | - | ↓ Phosphorylation of TAK1, IKK and JNK2 in cultured astrocytes | Ji et al. [169] |
| Sex not specified | Oral gavage | Pre-surgery administration—↓ Neuropathic pain symptoms when administered | X NF- activity via TAK1 in HEK293 cells | |||
| ↓ Protein expressions of TNF-α and IL-1β; mRNA expressions of MMP- 9, MMP-2 and CCL2 | ||||||
| Baicalin | Male Wistar rats | 10 µg | ↓ SNL-induced mechanical allodynia and thermal hyperalgesia | - | Reversed histone-H3 acetylation and HDAC1 expression of SNL-induced spinal cord dorsal horn | Cherng et al. [166] |
| Intrathecal | ↑Antinociceptive activity of morphine | |||||
| Epigallocatechin-3-gallate | Male Sprague Dawley rats | 10–50 mg/kg | ↓ SNL-induced mechanical allodynia | - | X nNOS expression in spinal cord of SNL rats | Choi et al. [167] |
| Intrathecal | ||||||
| Myricetin | Male Wistar rats | 0.1–10 mg/kg | ↓ SNL-induced mechanical allodynia and thermal hyperalgesia | Low concentrations—↓ Voltage activated calcium channel currents (ICa(V)) in vitro mediated by PKC but not p38 | - | Hagenacker et al. [168] |
| Intraperitoneal | ||||||
| 0.1–5 µM (low) 10–100 µM (high)—In vitro | High concentrations—↑ Voltage activated calcium channel currents (ICa(V)) in vitro mediated by p38 but not PKC | |||||
| Partial sciatic nerve ligation | ||||||
| Hesperetin | Wistar rats | 20, 50 mg/kg | ↓ Partial sciatic nerve ligation-induced mechanical and thermal hyperalgesia and mechanical allodynia | ↑ Motor nerve conduction velocity | ↓ TNF-α mRNA expression in sciatic nerve | Aswar et al. 2014 [178] |
| Either sex | Oral gavage | ↓ Different tissue biomarkers, such as total protein, NO, lipid peroxidase, IL-1β and IL-6 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basu, P.; Basu, A. In Vitro and In Vivo Effects of Flavonoids on Peripheral Neuropathic Pain. Molecules 2020, 25, 1171. https://doi.org/10.3390/molecules25051171
Basu P, Basu A. In Vitro and In Vivo Effects of Flavonoids on Peripheral Neuropathic Pain. Molecules. 2020; 25(5):1171. https://doi.org/10.3390/molecules25051171
Chicago/Turabian StyleBasu, Paramita, and Arpita Basu. 2020. "In Vitro and In Vivo Effects of Flavonoids on Peripheral Neuropathic Pain" Molecules 25, no. 5: 1171. https://doi.org/10.3390/molecules25051171
APA StyleBasu, P., & Basu, A. (2020). In Vitro and In Vivo Effects of Flavonoids on Peripheral Neuropathic Pain. Molecules, 25(5), 1171. https://doi.org/10.3390/molecules25051171