
Biological Activity of Hydrophilic Extract of Chlorella vulgaris Grown on Post-Fermentation Leachate from a Biogas Plant Supplied with Stillage and Maize Silage


 ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
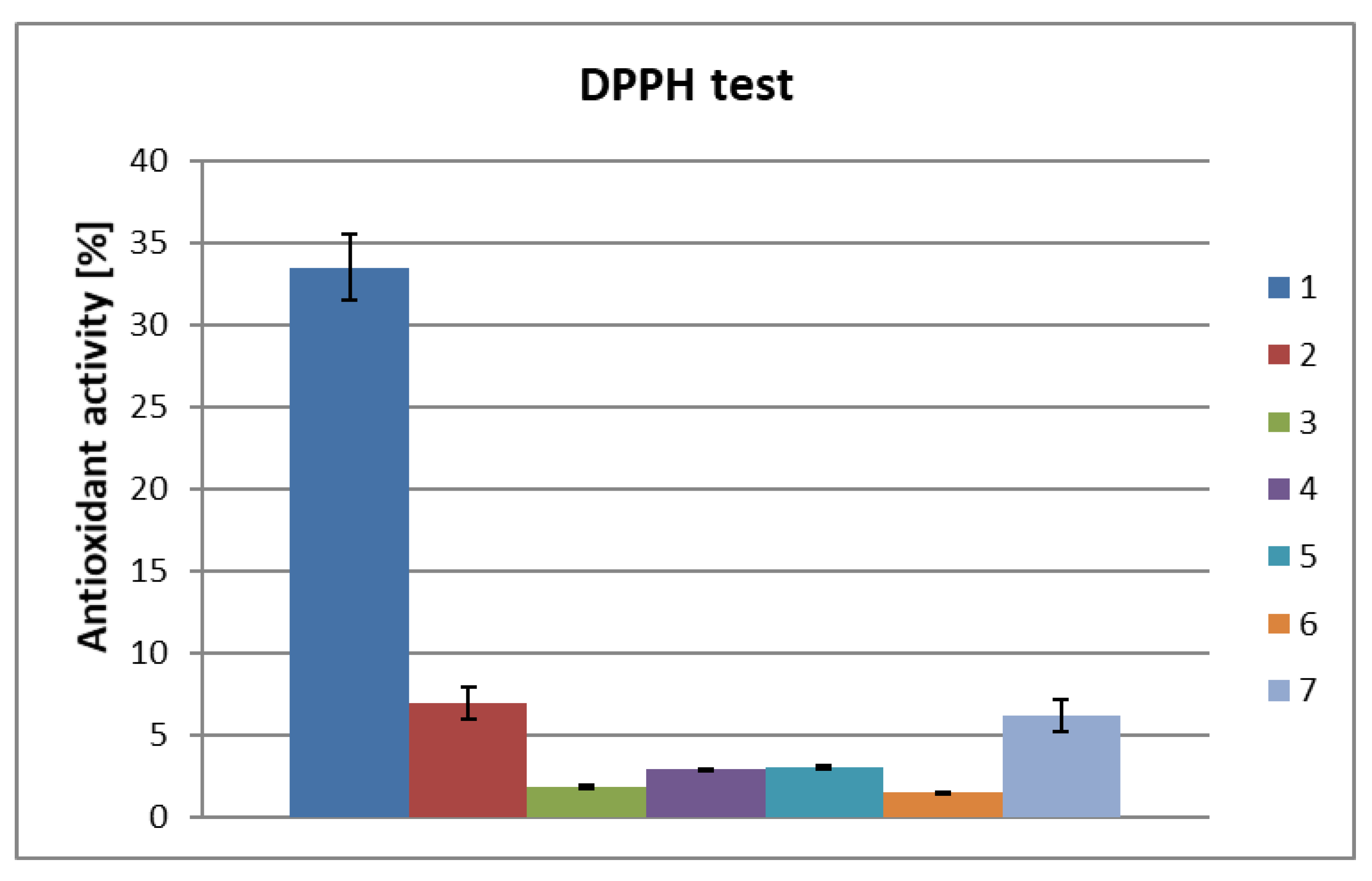
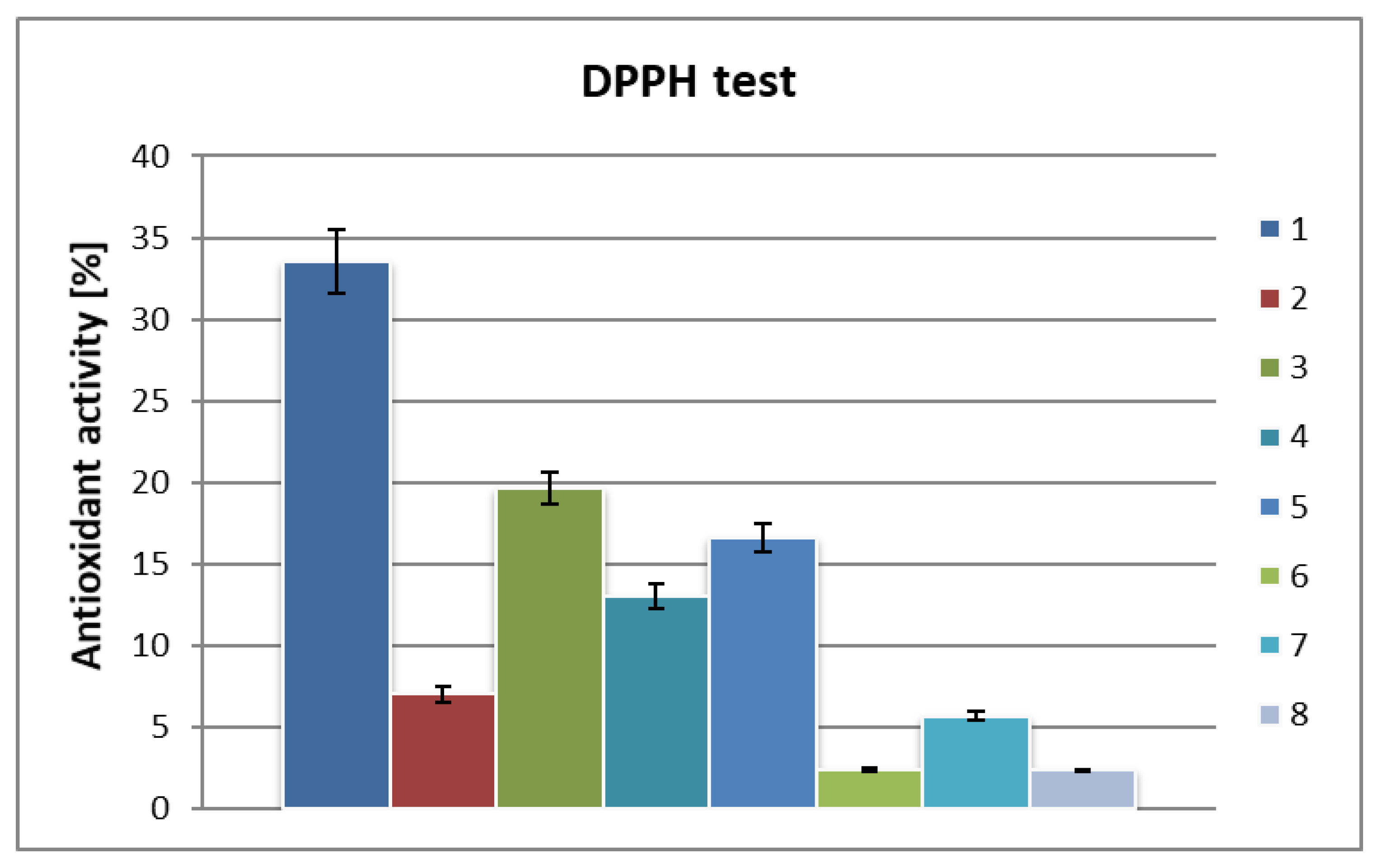
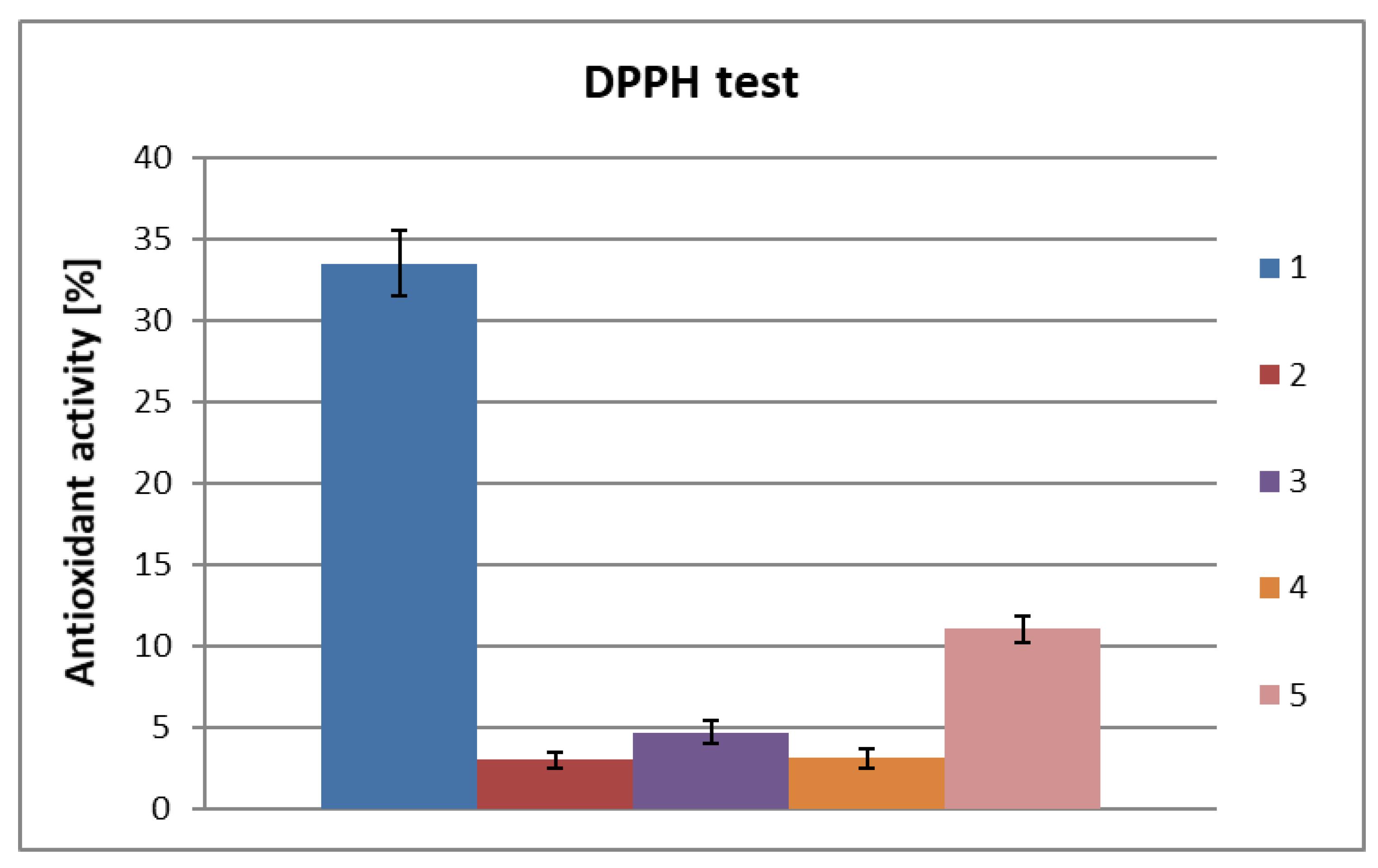
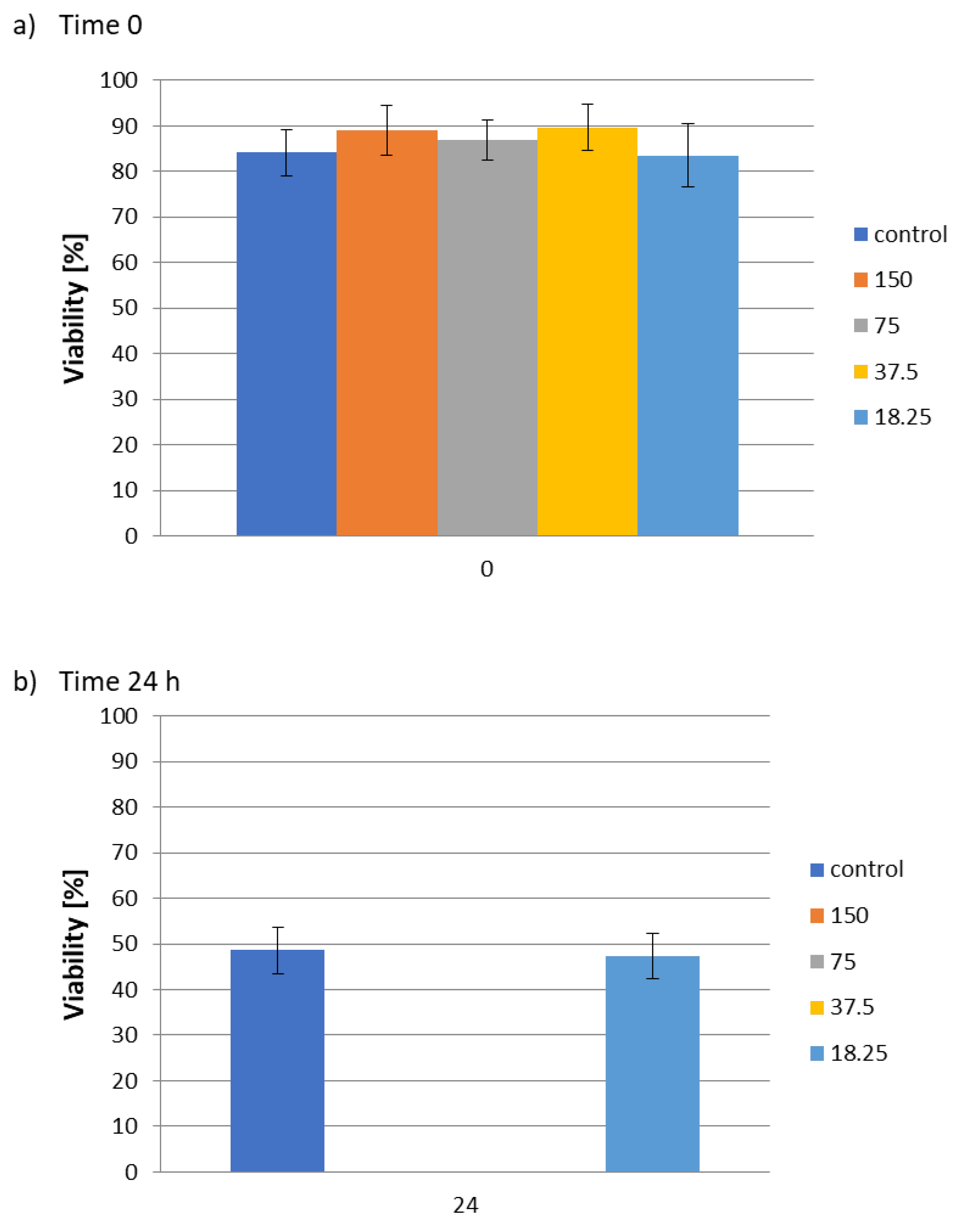
2. Results and Discussion
3. Materials and Methods
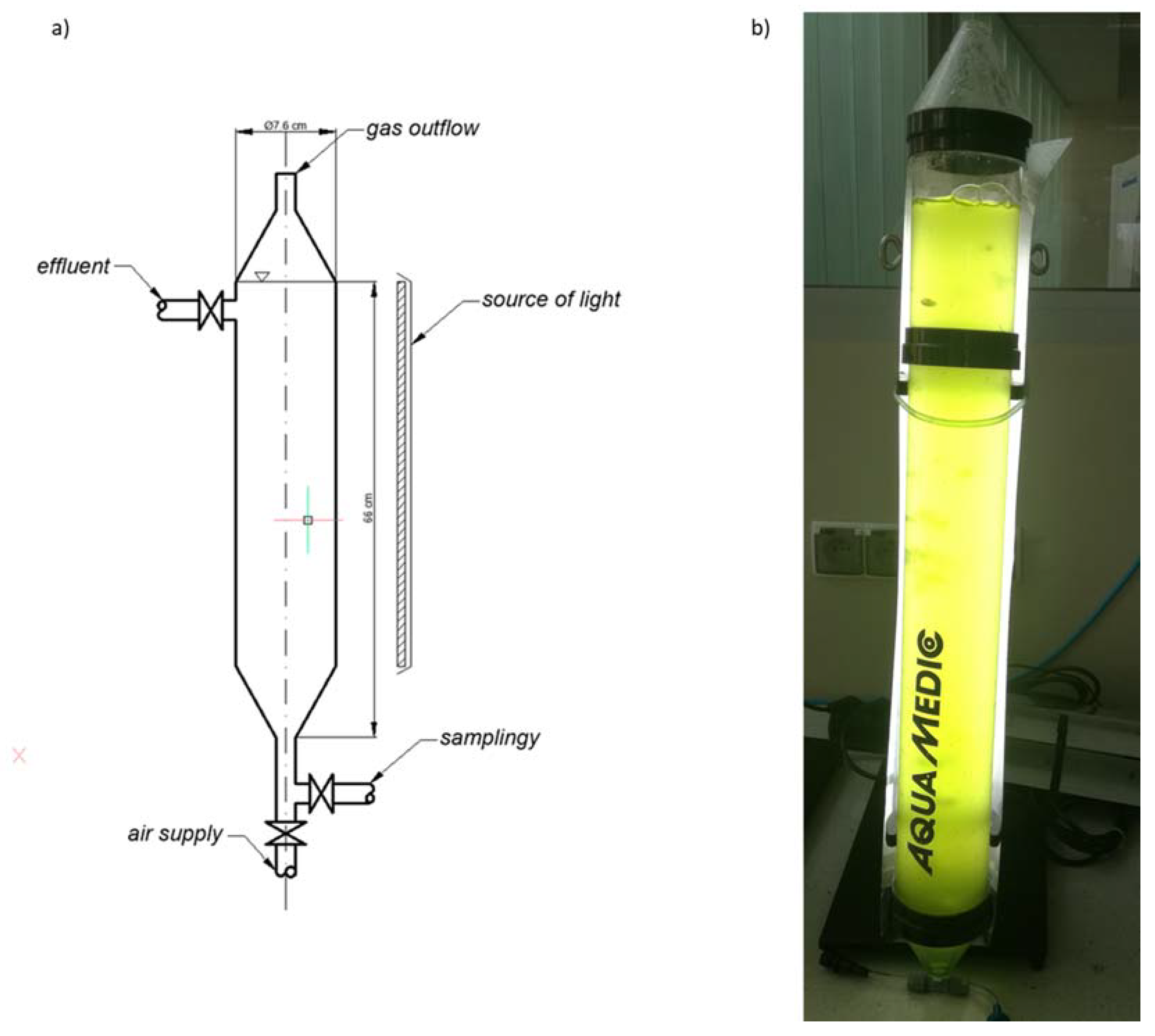
3.1. Research Stand
3.2. Analytical Methods for Assessing the Microalgae Culture
3.3. Disintegration of Cell Walls
3.4. Isolation of the Hydrophilic Fraction
3.5. Isolation of Protein/Polypeptide Fraction
3.6. Ultrafiltration
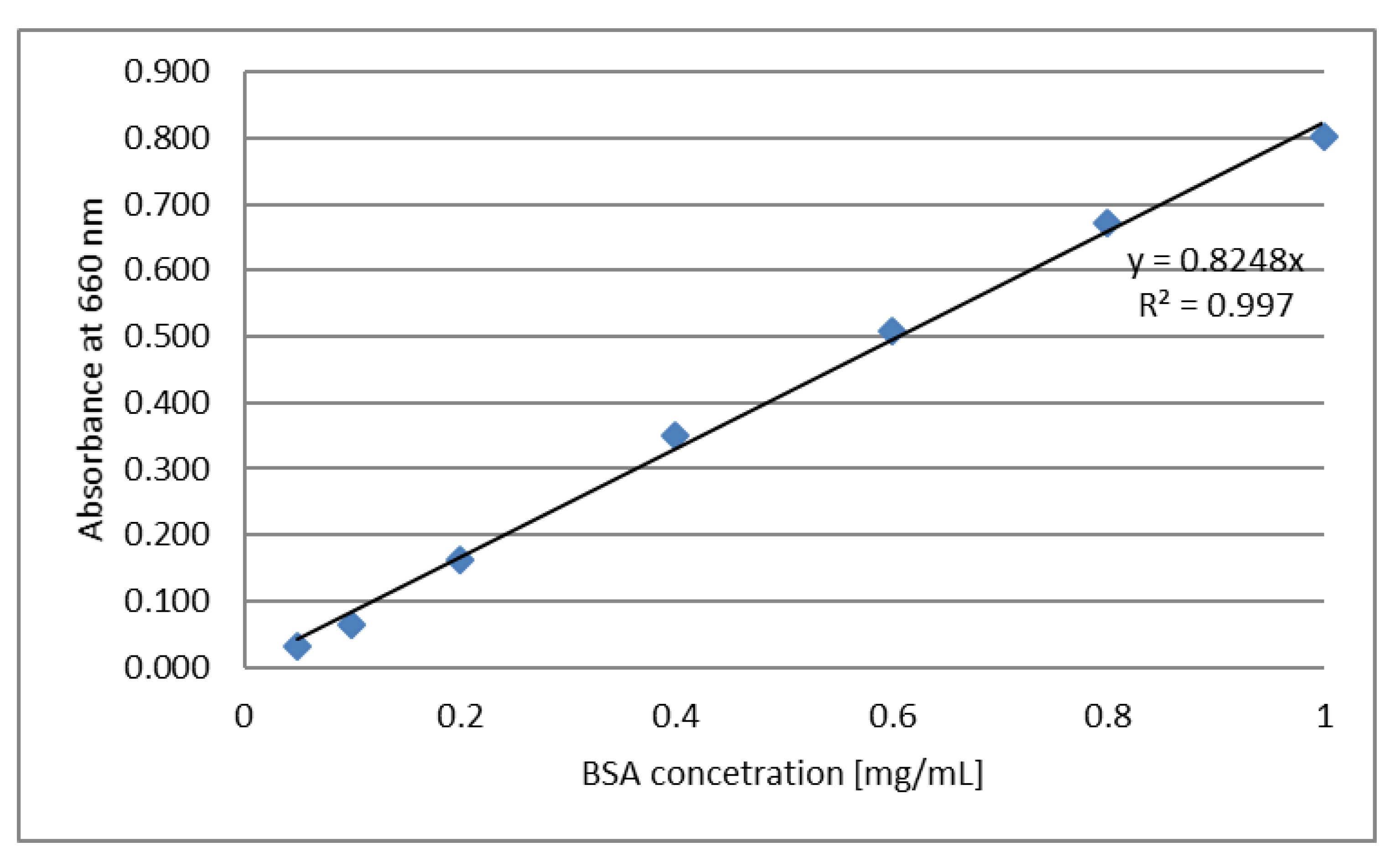
3.7. Determination of Protein Content by Lowry’s Method
3.8. Determination of Polyphenol Content
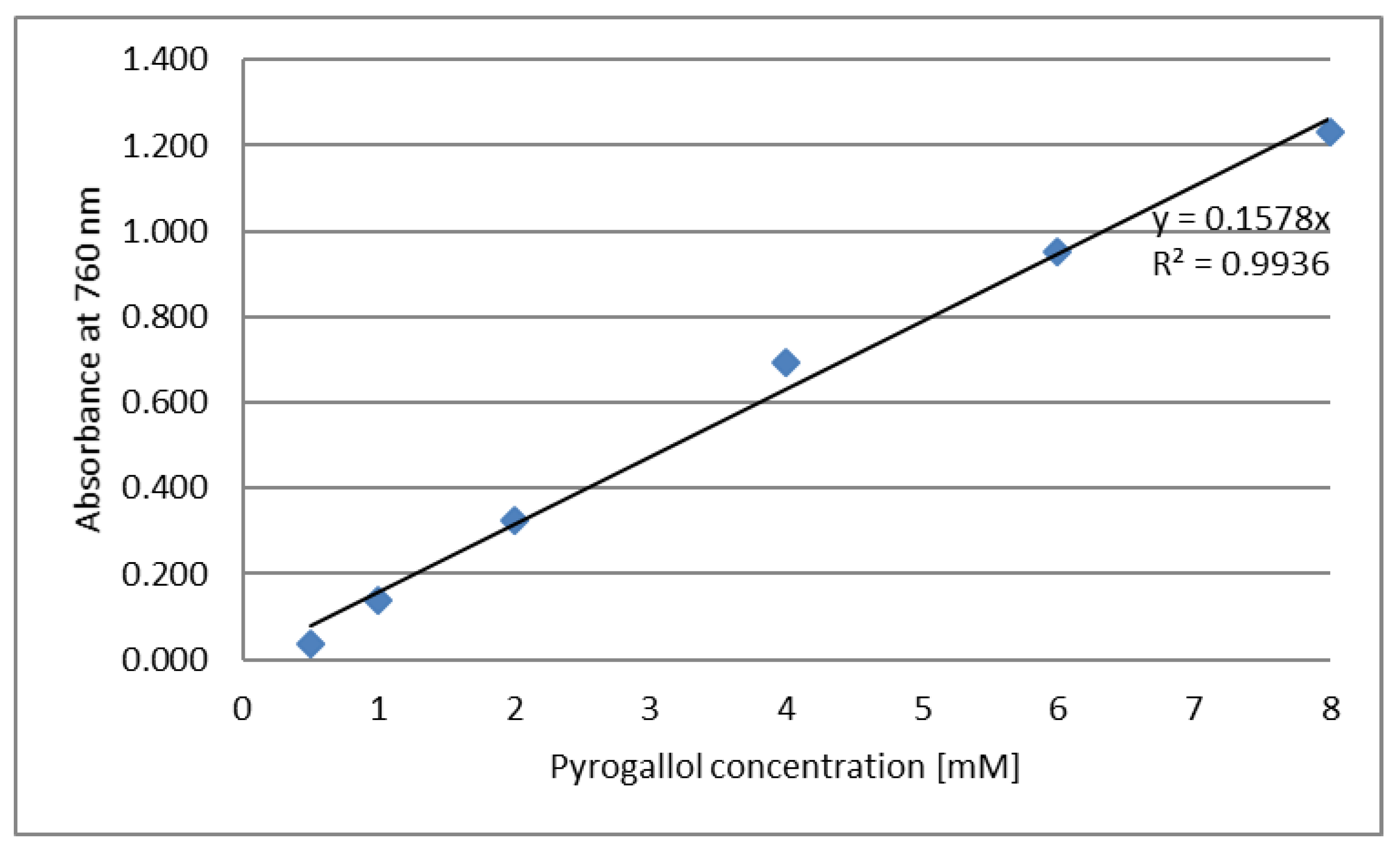
3.9. Antioxidant Activity
- A0–absorbance of DPPH solution
- At–absorbance of the tested sample after 30 min incubation with DPPH solution
- As–absorbance of the tested sample without DPPH solution.
3.10. Microbiological Tests
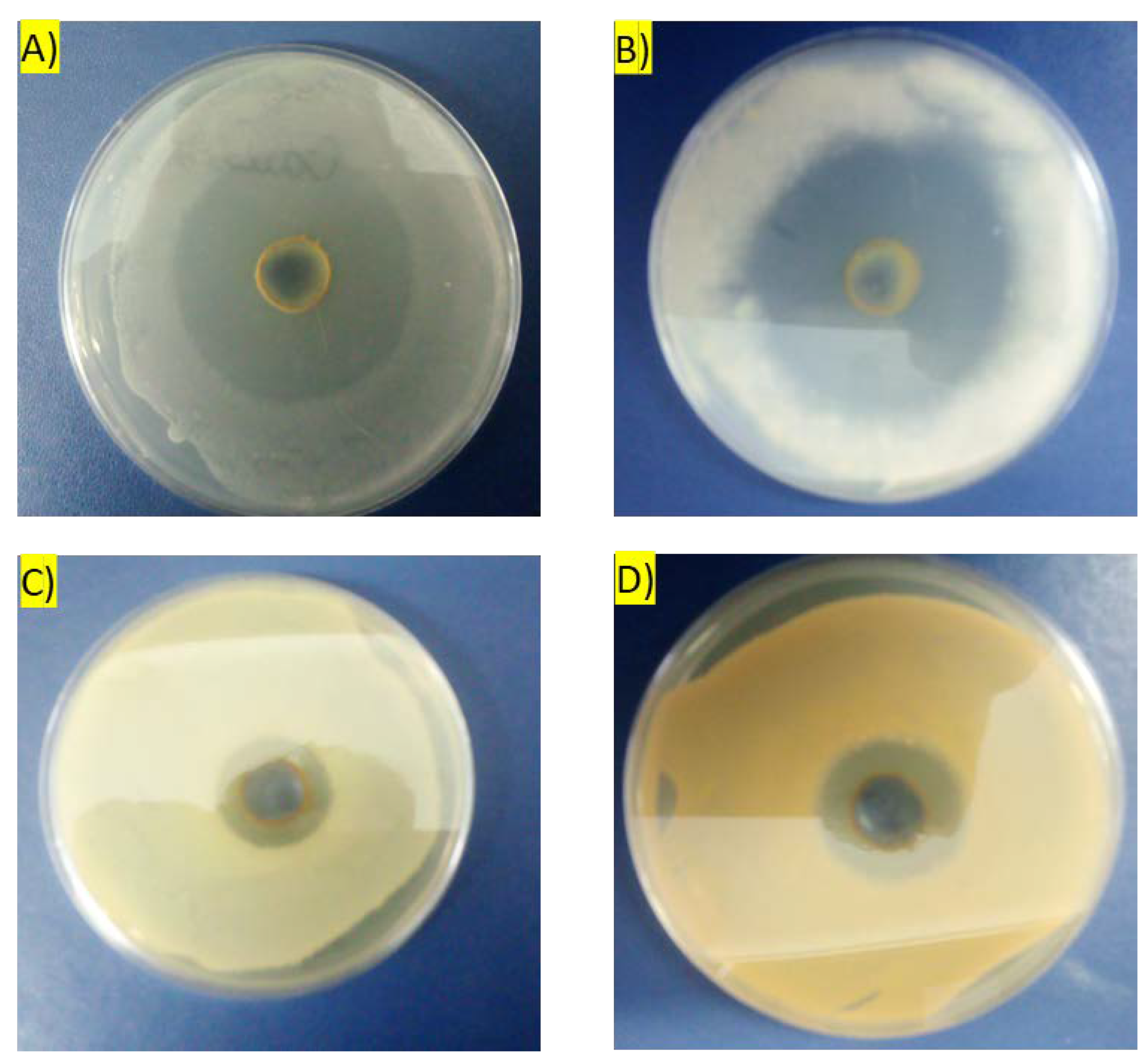
3.10.1. Inhibition Zone Plate Assay
3.10.2. Minimum Inhibitory Concentration Assay
3.11. Nematode Activity Assay
- V–viability [%]
- Vl–number of living nematodes
- Vn–number of all nematodes
3.12. Statistical Methods
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Priyadarshani, I.; Rath, B. Commercial and industrial applications of micro algae —A review. J. Algal Biomass Utln. 2012, 3, 89–100. [Google Scholar]
- Wajahatullah, K.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Prasanth, R.; Mark, H.D.; Alan, C.T.; James, C.S.; Jeff, N.; Balakrishan, P. Seaweed Extracts as Biostimulants of Plant Growth and Development. J. Plant. Growth. Regul. 2009, 28, 386–399. [Google Scholar]
- Garson, J. Marine Natural Products. Nat. Prod. Rep. 1989, 6, 143–170. [Google Scholar] [CrossRef]
- Saleh, B.; Al-Mariri, A. Antimicrobial Activity of the Marine Algal Extracts against Selected Pathogens. J. Agr. Sci. Tech. 2017, 19, 1067–1077. [Google Scholar]
- Sode, S.; Bruhn, A.; Balsby, T.J.S.; Larsen, M.M.; Gotfredsen, A.; Rasmussen, M.B. Bioremediation of Reject Water from Anaerobically Digested Waste Water Sludge with Macroalgae (Ulva lactuca, Chlorophyta). Bioresour. Technol. 2013, 146, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Oumaskour, K.; Boujaber, N.; Etahiri, S.; Assobhel, O. Anti-Inflammatory and Antimicrobial Activities of Twenty-Three Marine Algae from the Coast of SidiBouzid (El Jadida-Morocco). Int. J. Pharm. Pharm. Sci. 2013, 5, 145–149. [Google Scholar]
- Kausalya, M.; Rao, G.M.N. Antimicrobial Activity of Marine Algae. J. Algal Biomass Utln. 2015, 6, 78–87. [Google Scholar]
- Zbakh, H.; Chiheb, H.; Bouziane, H.; Sánchez, V.M.; Riadi, H. Antibacterial Activity of Benthic Marine Algae Extracts from the Mediterranean Coast of Morocco. J. Microb. Biotech. Food Sci. 2012, 2, 219–228. [Google Scholar]
- Malingin, D.L.; Hongayo, M.C.; Larino, R.C. Antibacterial and Antioxidant Effects of Brown Alga Padina australis Hauck Crude Extract. IAMURE Int. J. Sci. Clin. Lab. 2012, 2, 35–70. [Google Scholar]
- Jeyaseelan, E.C.; Kothai, S.; Kavitha, R.; Tharmila, S.; Thavaranjit, A.C. Antibacterial Activity of Some Selected Algae Present in the Costal Lines of Jaffna Peninsula. Int. J. Pharm. Biol. Arch. 2012, 3, 352–356. [Google Scholar]
- Alghazeer, R.; Whida, F.; Abduelrhman, E.; Gammoudi, F.; Azwai, S. Screening of Antibacterial Activity in Marine Green, Red and Brown Macroalgae from the Western Coast of Libya. Nat. Sci. 2013, 5, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Abo-State, M.A.M.; Shanab, S.M.M.; Ali, H.E.A.; Abdullah, M.A. Screening of Antimicrobial Activity of Selected Egyptian Cyanobacterial Species. J. Ecol. Health Environ. 2015, 3, 7–13. [Google Scholar]
- Karabay-Yavasoglu, N.U.; Sukatar, A.; Ozdemir, G.; Horzum, Z. Antimicrobial Activity of Volatile Components and Various Extracts of the Red Alga Janiarubens. Phytother. Res. 2007, 21, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; Abdelnourm, S.; Alagawany, M.; Abdo, M.; Sakr, M.A.; Khafaga, A.F.; Mahgoub, S.A.; Elnesr, S.S.; Gebriel, M.G. Microalgae in modern cancer therapy: Current knowledge. Biomed. Pharmacother. 2019, 111, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Martínez Andrade, K.A.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [Green Version]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2018, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.H.; Kim, S.K. Beneficial effect of peptides from microalgae on anticancer. Curr. Protein Pept. Sci. 2013, 14, 212–217. [Google Scholar] [CrossRef]
- Villarruel-López, A.; Ascencio, F.; Nuño, K. Microalgae, a Potential Natural Functional Food Source —A Review. Pol. J. Food Nutr. Sci. 2017, 67, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Chu, W.L. Biotechnological applications of microalgae. IeJSME 2012, 6, 24–37. [Google Scholar]
- Parmar, A.; Singh, N.K.; Pandey, A.; Gnansounou, E.; Madamwar, D. Cyanobacteria and microalgae: A positive prospect for biofuels. Bioresour. Technol. 2011, 102, 10163–10172. [Google Scholar] [CrossRef]
- Mostafa, S.S.M. Microalgal Biotechnology: Prospects and Applications. Plant. Sci. 2012, 12, 275–314. [Google Scholar]
- Ibañez, E.; Cifuentes, A. Benefits of using algae as natural sources of functional ingredients. J. Sci. Food Agric. 2013, 93, 703–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, A.P.; Gouveia, L.; Bandarra, N.M.; Franco, J.M.; Raymundo, A. Comparison of microalgal biomass profiles as novel functional ingredient for food products. Algal Res. 2013, 2, 164–173. [Google Scholar] [CrossRef] [Green Version]
- Safi, C.; Zebib, B.; Merah, O.; Pontalier, P.Y.; Vaca-Garcia, C. Morphology, composition, production, processing and applications of Chlorella vulgaris: A review. Renew. Sust. Energ. Rev. 2014, 35, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Fradique, M.; Batista, A.P.; Nunes, M.C.; Gouveia, L.; Bandarra, N.M.; Raymundo, A. Incorporation of Chlorella vulgaris and Spirulina maxima biomass in pasta products. Part 1: Preparation and evaluation. J. Sci. Food Agric. 2010, 90, 1656–1664. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-B.; Jiang, Y.; Chen, F. Isolation and purification of lutein from the microalga Chlorella vulgaris by extraction after saponification. J. Agric. Food Chem. 2002, 50, 1070–1072. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, B.; Dragone, G.; Abreu, A.; Geada, P.; Teixeira, J.; Vicente, A. Starch determination in Chlorella vulgaris – a comparison between acid and enzymatic methods. J. Appl. Phycol. 2012, 24, 1203–1208. [Google Scholar] [CrossRef] [Green Version]
- Grobbelaar, J.U. Quality Control and Assurance: Crucial for the sustainability of the applied phycology industry. J. Appl. Phycol. 2003, 15, 209–215. [Google Scholar] [CrossRef]
- Gulati, O.P.; Berry Ottaway, P. Legislation relating to nutraceuticals in the European Union with a particular focus on botanical-sourced products. Toxicology 2006, 221, 75–87. [Google Scholar] [CrossRef]
- Rodriguez-Garcia, I.; Guil-Guerrero, J.L. Evaluation of the antioxidant activity of three microalgal species for use as dietary supplements and in the preservation of foods. Food Chem. 2008, 108, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, L.; Oliveira, C. Microalgae as a raw material for biofuels production. J. Ind. Microbiol. Biotechnol. 2009, 36, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.R.; Passarinho, P.C.; Gouveia, L. Bioethanol production from Scenedesmus obliquus sugars: The influence of photobioreactors and culture conditions on biomass production. Appl. Microbiol. Biotechnol. 2012, 96, 555–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, A.E.; Barbosa, T.A.; Jotta, J.; Tamagnini, P.; Gouveia, L. Biohydrogen production by Anabaena sp. PCC 7120 wild-type and mutants under different conditions: Light, Nickel and CO2. Biomass Bioenerg. 2011, 35, 4426–4434. [Google Scholar] [CrossRef]
- Ferreira, A.F.; Ribeiro, L.; Batista, A.P.; Marques, P.A.S.S.; Nobre, B.P.; Silva, P.P.; Gouveia, L.; Silva, C. A Biorefinery from Nannochloropsis sp. microalga —Energy and CO2 emission and economic analyses. Bioresour. Technol. 2013, 138, 235–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, A.P.; Moura, P.; Marques, P.A.S.S.; Ortigueira, J.; Alves, L.M.; Gouveia, L. Scenedesmus obliquus as a feedstock for bio-hydrogen production by Enterobacter aerogenes and Clostridium butyricum by dark fermentation. Fuel 2014, 117, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, L. From Tiny Microalgae to Huge Biorefineries. Oceanography 2014, 2, 120. [Google Scholar]
- Allen, J.; Unlu, S.; Demirel, Y.; Black, P.; Riekhof, W. Integration of biology, ecology and engineering for sustainable algal-based biofuel and bioproduct biorefinery. Bioresour. Bioprocess. 2018, 5, 47. [Google Scholar] [CrossRef] [Green Version]
- Schiano di Visconte, G.; Spicer, A.; Chuck, C.J.; Allen, M.J. The Microalgae Biorefinery: A Perspective on the Current Status and Future Opportunities Using Genetic Modification. Appl. Sci. 2019, 9, 4793. [Google Scholar] [CrossRef] [Green Version]
- Koyande, A.K.; Show, P.-L.; Guo, R.; Tang, B.; Ogino, C.; Chang, J.-S. Bio-processing of algal bio-refinery: A review on current advances and future perspectives. Bioengineered. 2019, 10, 574–592. [Google Scholar] [CrossRef] [Green Version]
- Laurens, L.M.L.; Markham, J.; Templeton, D.W.; Christensen, E.D.; Van Wychen, S.; Vadelius, E.W.; Chen-Glasser, M.; Dong, T.; Davis, R.; Pienkos, P.T. Development of algae biorefinery concepts for biofuels and bioproducts; a perspective on process-compatible products and their impact on cost-reduction. Energy Environ. Sci. 2017, 10, 1716–1738. [Google Scholar] [CrossRef] [Green Version]
- Kolesinska, B.; Fraczyk, J.; Binczarski, M.; Modelska, M.; Berlowska, J.; Dziugan, P.; Antolak, H.; Kaminski, Z.J.; Witonska, I.A.; Kregiel, D. Butanol Synthesis Routes for Biofuel Production: Trends and Perspectives. Materials 2019, 12, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collet, P.; Hélias, A.; Lardon, L.; Ras, M.; Goy, R.A.; Steyer, J.P. Lifecycle assessment of microalgae culture coupled to biogas production. Bioresour. Technol. 2011, 102, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Ehimen, E.A.; Sun, Z.F.; Carrington, C.G.; Birch, E.J.; Eaton-Rye, J.J. Anaerobic digestion of microalgae residues resulting from the biodiesel production process. Appl. Energ. 2011, 88, 3454–3463. [Google Scholar] [CrossRef]
- Gouveia, L.; Neves, C.; Sebastião, D.; Nobre, B.P.; Matos, C.T. Effect of light on the production of bioelectricity and pigments by a Photosynthetic Alga Microbial Fuel Cell. Bioresour. Technol. 2014, 154, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Powel, E.E.; Hill, G.A. Economic assessment of an integrated bioethanol-biodiesel-microbial fuel cell facility utilizing yeast and photosynthetic algae. Chem. Eng. Res. Design 2009, 87, 1340–1348. [Google Scholar] [CrossRef]
- Campenni’, L.; Nobre, B.P.; Santos, C.A.; Oliveira, A.C.; Aires-Barros, M.R.; Palavra, A.M.; Gouveia, L. Carotenoids and lipids production of autotrophic microalga Chlorella protothecoides under nutritional, salinity and luminosity stress conditions. Appl. Microbiol. Biotechnol. 2013, 97, 1383–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mussgnug, J.H.; Klassen, V.; Schlüter, A.; Kruse, O. Microalgae as substrates for fermentative biogas production in a combined biorefinery concept. J. Biotechnol. 2010, 150, 51–56. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Acharya, M.M.; Katyare, S.S. An Improved Micromethod for Tyrosine Estimation. Z. Naturforsch. 2004, 59, 897–900. [Google Scholar] [CrossRef] [Green Version]
- Barbarino, E.; Louren, S.O. An evaluation of methods for extraction and quantification of protein from marine macro- and microalgae. J. Appl. Phycol. 2005, 17, 447–460. [Google Scholar] [CrossRef]
- Becker, E.W. Micro-algae as a source of protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef]
- Safi, C.; Charton, M.; Pignolet, O.; Silvestre, F.; Vaca-Garcia, C.; Pontalier, P.-Y. Influence of microalgae cell wall characteristics on protein extractability and determination of nitrogen-to-protein conversion factors. J. Appl. Phycol. 2013, 25, 523–529. [Google Scholar] [CrossRef] [Green Version]
- Berliner, M.D. Proteins in Chlorella vulgaris. Microbios 1986, 46, 199–203. [Google Scholar]
- Vehapi, M.; Koçer, A.T.; Yılmaz, A.; Özçimen, D. Investigation of the antifungal effects of algal extracts on apple-infecting fungi. Arch. Microbiol. 2019. [Google Scholar] [CrossRef]
- Selivanova, E.A.; Ignatenko, M.E.; Nemtseva, N.V. Antagonistic activity of novel green microalgae strain. Zh. Mikrobiol. Epidemiol. Immunobiol. 2014, 4, 72–76. [Google Scholar]
- Pina-Pérez, M.C.; Rivas, A.; Martínez, A.; Rodrigo, D. Antimicrobial potential of macro and microalgae against pathogenic and spoilage microorganisms in food. Food Chem. 2017, 235, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Mudimu, O.; Rybalka, N.; Bauersachs, T.; Born, J.; Friedl, T.; Schulz, R. Biotechnological Screening of Microalgal and Cyanobacterial Strains for Biogas Production and Antibacterial and Antifungal Effects. Metabolites 2014, 4, 373–393. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, Y.; Moradian, A.; Mohagheghzadeh, A.; Shokravi, S.; Morowvat, M.H. Antifungal and antibacterial activity of the microalgae collected from paddy fields of Iran: Characterization of Antimicrobial Activity of Chroococcus dipersus. J. Biol. Sci. 2007, 7, 904–910. [Google Scholar]
- Gaugler, R.; Lewis, E.; Stuart, R.J. Ecology in the service of biological control: The case of entomopathogenic nematodes. Oecologia 1997, 109, 483–489. [Google Scholar] [CrossRef]
- Choleva, B.; Bileva, T.; Tzvetkov, Y.; Barakov, P. Preliminary study of the green algae chlorella (Chlorella vulgaris) for control on the root-knot nematode (Meloidogyne arenaria) in tomato plants and ectoparasite Xiphinema indexin grape seedlings. Commun. Agric. Appl. Biol. Sci. 2005, 70, 915–926. [Google Scholar]
- Debowski, M.; Szwaja, S.; Zielinski, M.; Kisielewska, M.; Stanczyk-Mazanek, E. The Influence of Anaerobic Digestion Effluents (ADEs) Used as the Nutrient Sources for Chlorella sp. Cultivation on Fermentative Biogas Production. Waste Biomass Valori. 2017, 8, 1153–1161. [Google Scholar] [CrossRef] [Green Version]
- Debowski, M.; Rusanowska, P.; Zielinski, M.; Dudek, M.; Romanowska-Duda, Z. Biomass production and nutrient removal by Chlorella vulgaris from anaerobic digestion effluents. Energies 2018, 11, 1654. [Google Scholar] [CrossRef] [Green Version]
- Zielinski, M.; Debowski, M.; Szwaja, S.; Kisielewska, M. Anaerobic Digestion Effluents (ADEs) Treatment Coupling with Chlorella sp. Microalgae Production. Water Environ. Res. 2018, 90, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin-phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Agbor, G.A.; Vinson, J.A.; Donnelly, P.E. Folin-Ciocalteau Reagent for Polyphenolic Assay. Int. J. Food Sci. Nutr. Diet. 2014, 3, 147–156. [Google Scholar] [CrossRef]
- Zych, I.; Krzepiłko, A. Measurement of total antioxidant capacity of selected antioxidants and infusions using DPPH radical reduction. Chem.-Didact.-Ecol.-Metrol. 2010, 1, 51–54. [Google Scholar]
Sample Availability: Samples of the extracts are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Concentration of Tested Samples [mg/mL H2O] | Polyphenol Content (Folin–Ciocalteu Test) [mM] * |
|---|---|---|
| S1—water extraction, first repetition | 27.0 | 0.77 |
| S2—water extraction, second repetition | 21.5 | 0.84 |
| S3—water extraction, third repetition | 20.6 | 0.71 |
| S4—protein fraction obtained from S3 treated with TCA and then centrifuged once | 18.8 | 0.33 |
| S5—protein fraction obtained from S3 treated with TCA and then after isolation of S4. The supernatant was subjected to five centrifugations | 19.6 | 0.16 |
| MIC [mg/mL] | L. plantarum | S. epidermidis | E. coli | C. albicans | S. cerevisiae |
|---|---|---|---|---|---|
| water extract | 37.5–75 | 150 | 150 | 37.5–75 | lack of activity |
| protein fraction precipitated by TCA, pellet obtained by a single centrifugation | 16.25 | 16.25 | 32.5–65 | 32.5 | 32.50 |
| Parameters | Unit | Effluent | |
|---|---|---|---|
| Crude | After Distillation | ||
| Dry Mass (DM) | mg/L | 12740 ± 260 | 21 ± 4 |
| Chemical Oxygen Demand | mg O2/L | 8190 ± 170 | 1370 ± 20 |
| Biochemical Oxygen Demand | mg O2/L | 3960 ± 90 | 460 ± 30 |
| Ntotal | mg N/L | 3710 ± 100 | 2150 ± 110 |
| N-NH4 | mg N-NH4/L | 2540 ± 70 | 1450 ± 40 |
| Ptotal | mg P/L | 350 ± 20 | 6.0 ± 2.0 |
| P-PO4 | mg P-PO4/L | 240 ± 10 | 4.0 ± 1.0 |
| pH | - | 6.9 ± 0.4 | 7.1 ± 0.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zielinski, D.; Fraczyk, J.; Debowski, M.; Zielinski, M.; Kaminski, Z.J.; Kregiel, D.; Jacob, C.; Kolesinska, B. Biological Activity of Hydrophilic Extract of Chlorella vulgaris Grown on Post-Fermentation Leachate from a Biogas Plant Supplied with Stillage and Maize Silage. Molecules 2020, 25, 1790. https://doi.org/10.3390/molecules25081790
Zielinski D, Fraczyk J, Debowski M, Zielinski M, Kaminski ZJ, Kregiel D, Jacob C, Kolesinska B. Biological Activity of Hydrophilic Extract of Chlorella vulgaris Grown on Post-Fermentation Leachate from a Biogas Plant Supplied with Stillage and Maize Silage. Molecules. 2020; 25(8):1790. https://doi.org/10.3390/molecules25081790
Chicago/Turabian StyleZielinski, Dariusz, Justyna Fraczyk, Marcin Debowski, Marcin Zielinski, Zbigniew J. Kaminski, Dorota Kregiel, Claus Jacob, and Beata Kolesinska. 2020. "Biological Activity of Hydrophilic Extract of Chlorella vulgaris Grown on Post-Fermentation Leachate from a Biogas Plant Supplied with Stillage and Maize Silage" Molecules 25, no. 8: 1790. https://doi.org/10.3390/molecules25081790
APA StyleZielinski, D., Fraczyk, J., Debowski, M., Zielinski, M., Kaminski, Z. J., Kregiel, D., Jacob, C., & Kolesinska, B. (2020). Biological Activity of Hydrophilic Extract of Chlorella vulgaris Grown on Post-Fermentation Leachate from a Biogas Plant Supplied with Stillage and Maize Silage. Molecules, 25(8), 1790. https://doi.org/10.3390/molecules25081790