
Quercetin and Rutin as Modifiers of Aphid Probing Behavior


Abstract
:1. Introduction
2. Results
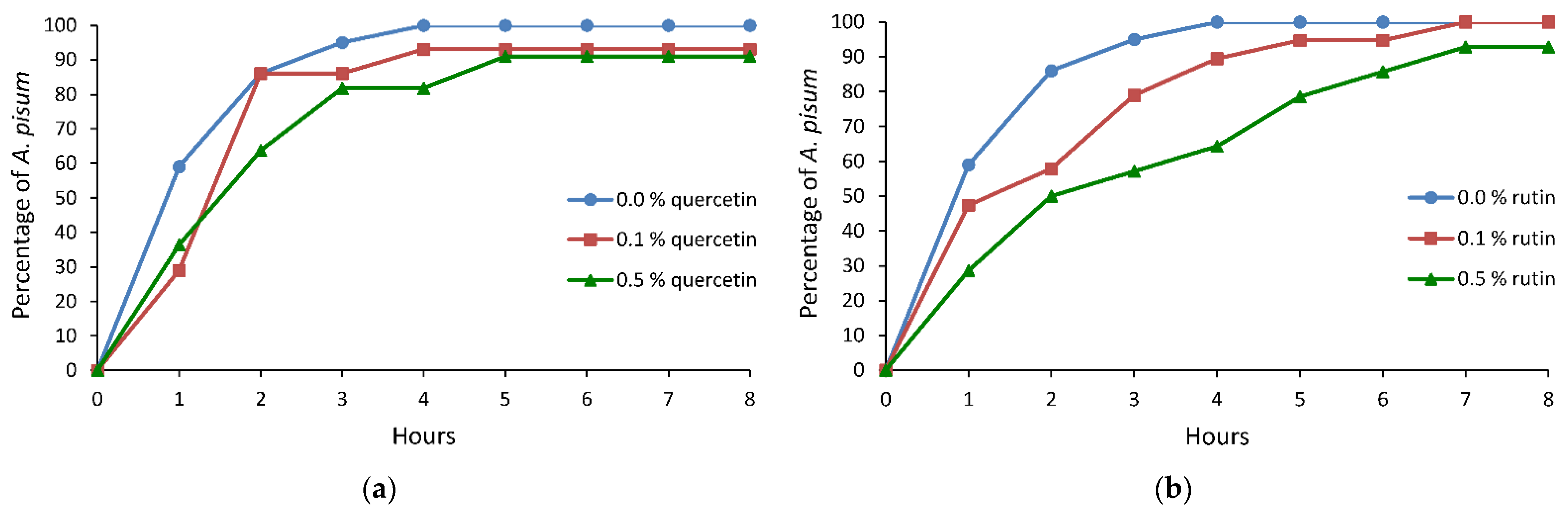
2.1. Probing Behavior of Acyrthosiphon pisum on Pisum sativum
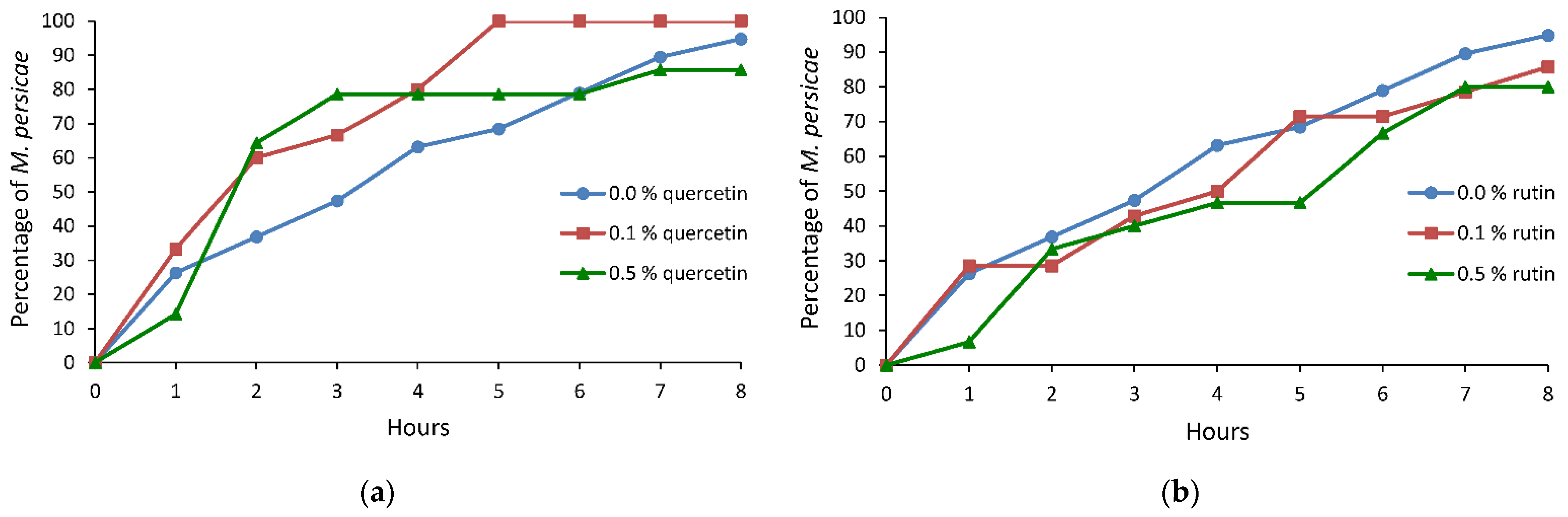
2.2. Probing and Settling Behavior of Myzus persicae on Brassica rapa subsp. pekinensis
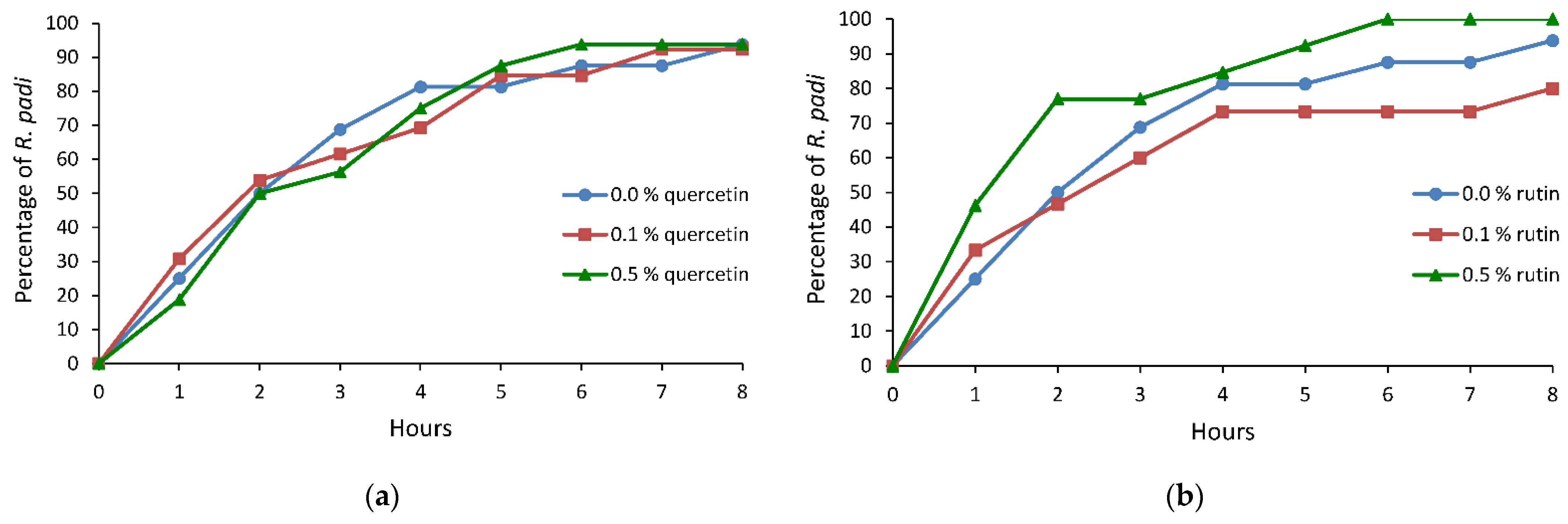
2.3. Probing and Settling Behavior of Rhopalosiphum padi on Avena sativa
3. Discussion
4. Materials and Methods
4.1. Cultures of Plants and Aphids
4.2. Application of Quercetin and Rutin
4.3. Behavioral Responses of Aphids
- Aphid Probing Behavior (No-Choice Test)
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pettersson, J.; Tjallingii, W.F.; Hardie, J. Host-plant selection and feeding. In Aphids as Crop Pests, 2nd ed.; van Emden, H.F., Harrington, R., Eds.; CABI: Wallingford, UK, 2017; pp. 173–195. [Google Scholar]
- Tjallingii, W.F.; Hogen Esch, T.H. Fine-structure of aphid stylet routes in plant-tissues in correlation with EPG signals. Physiol. Entomol. 1993, 18, 317–328. [Google Scholar] [CrossRef]
- Stevens, M.; Lacomme, C. Transmission of plant viruses. In Aphids as Crop Pests, 2nd ed.; van Emden, H.F., Harrington, R., Eds.; CABI: Wallingford, UK, 2017; pp. 323–361. [Google Scholar]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification Guide, 1st ed.; John Wiley and Sons: Chichester, UK; New York, NY, USA; Brisbane, Australia; Toronto, Japan; Singapore, 1985. [Google Scholar]
- Dedryver, C.-A.; Ralec, A.L.; Fabre, F. The conflicting relationships between aphids and men: A review of aphid damage and control strategies. Comptes Rendus Biol. 2010, 333, 539–553. [Google Scholar] [CrossRef]
- Köhler, H.R.; Triebskorn, R. Wildlife ecotoxicology of pesticides: Can we track effects to the population level and beyond? Science 2013, 341, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Casida, J.E.; Durkin, K.A. Neuroactive insecticides: Targets, selectivity, resistance, and secondary effects. Annu. Rev. Entomol. 2013, 58, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, J.R.; Dachbrodt-Saaydeh, S.; Kudsk, P.; Messean, A. Toward a reduced reliance in conventional pesticides in European agriculture. Plant Dis. 2016, 110, 10–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, S.P.; Devine, G.; Devonshire, A.L. Insecticide resistance. In Aphids as Crop Pests, 2nd ed.; van Emden, H.F., Harrington, R., Eds.; CABI: Wallingford, UK, 2017; pp. 426–447. [Google Scholar]
- Isman, M. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Zehnder, G.; Gurr, G.M.; Kuhne, S.; Wade, M.R.; Wratten, S.D.; Wyss, E. Arthropod pest management in organic crops. Annu. Rev. Entomol. 2007, 52, 57–80. [Google Scholar] [CrossRef] [Green Version]
- Pickett, J.A.; Khan, Z.R. Plant volatile-mediated signalling and its application in agriculture: Successes and challenges. New Phytol. 2016, 212, 856–870. [Google Scholar] [CrossRef] [Green Version]
- Rivera, M.J.; Martini, X.; Conover, D.; Mafra-Neto, A.; Carrillo, D.; Stelinski, L.L. Evaluation of semiochemical based push-pull strategy for population suppression of ambrosia beetle vectors of laurel wilt disease in avocado. Sci. Rep. 2020, 10, 2670. [Google Scholar] [CrossRef] [Green Version]
- Frazier, J.L.; Chyb, S. Use of feeding inhibitors in insect control. In Regulatory Mechanisms in Insect Feeding, 1st ed.; Chapman, R.F., de Boer, G., Eds.; Chapman & Hall: New York, NY, USA, 1995; pp. 364–381. [Google Scholar]
- Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Developing sustainable pest control from chemical ecology. Agric. Ecosyst. Environ. 1997, 64, 149–156. [Google Scholar] [CrossRef]
- Pickett, J.A.; Woodcock, C.M.; Midega, C.A.O.; Khan, Z.R. Push–pull farming systems. Curr. Opin. Biotech. 2014, 26, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [Green Version]
- Koul, O. Insect Antifeedants; CRC Press: London, UK; New York, NY, USA; Washington, WA, USA, 2004; pp. 25–42. [Google Scholar]
- Goławska, S.; Łukasik, I. Antifeedant activity of luteolin and genistein against the pea aphid, Acyrthosiphon pisum. J. Pest Sci. 2012, 85, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Kaur, I.; Kariyat, R. The multifunctional roles of polyphenols in plant-herbivore interactions. Int. J. Mol. Sci. 2021, 22, 1442. [Google Scholar] [CrossRef] [PubMed]
- Soriano, I.R.; Asenstorfer, R.E.; Schmidt, O.; Riley, I.T. Inducible flavone in oats (Avena sativa) is a novel defense against plant-parasitic nematodes. Phytopathology 2004, 94, 1207–1214. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Carrillo, J.; Siemann, E.; Ding, J. Herbivore-specific induction of indirect and direct defensive responses in leaves and roots. AoB Plants. 2019, 11, plz003. [Google Scholar] [CrossRef] [PubMed]
- Calatayud, P.A.; Rahbé, Y.; Delobel, B.; Khuong-Huu, F.; Tertuliano, M.; Le Rü, B. Influence of secondary compounds in the phloem sap of cassava on expression of antibiosis towards the mealybug Phenacoccus manihoti. Entomol. Exp. Appl. 1994, 72, 47–57. [Google Scholar] [CrossRef]
- Simmonds, M.S.J. Importance of flavonoids in insect-plant interactions: Feeding and oviposition. Phytochemistry 2001, 56, 245–252. [Google Scholar] [CrossRef]
- Simmonds, M.S.J. Flavonoid-insect interactions: Recent advances in our knowledge. Phytochemistry 2003, 64, 21–30. [Google Scholar] [CrossRef]
- Goławska, S.; Sprawka, I.; Łukasik, I.; Goławski, A. Are naringenin and quercetin useful chemicals in pest-management strategies? J. Pest. Sci. 2014, 87, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Kozak, A.; Chrzanowski, G.; Sempruch, C.; Klewek, A.; Chwedczuk, M. Effect of selected flavonoids on the behavior of the bird cherry-oat aphid (Rhopalosiphum padi L.) during the colonization of winter wheat. Prog. Plant Prot. 2015, 55, 202–206. [Google Scholar]
- Shi, G.; Kang, Z.; Ren, F.; Zhou, Y.; Guo, P. Effects of quercetin on the growth and expression of immune-pathway-related genes in silkworm (Lepidoptera: Bombycidae). J. Insect. Sci. 2020, 20, 23. [Google Scholar] [CrossRef] [PubMed]
- Bentivenha, J.P.F.; Canassa, V.F.; Baldin, E.L.; Borguini, M.G.; Lima, G.P.; Lourenção, A.L. Role of the rutin and genistein flavonoids in soybean resistance to Piezodorus guildinii (Hemiptera: Pentatomidae). Arthropod. Plant Interact. 2018, 12, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Gómez, J.D.; Vital, C.E.; Oliveira, M.G.A.; Ramos, H.J.O. Broad range flavonoid profiling by LC/MS of soybean genotypes contrasting for resistance to Anticarsia gemmatalis (Lepidoptera: Noctuidae). PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Zavala, J.A.; Scopel, A.L.; Ballaré, C.L. Effects of ambient UV-B radiation on soybean crops: Impact on leaf herbivory by Anticarsia gemmatalis. Plant Ecol. 2001, 156, 121–130. [Google Scholar] [CrossRef]
- Calatayud, P.A. Influence of linamarin and rutin on biological performances of Phenacoccus manihoti in artificial diets. Entomol Exp. Appl. 2000, 96, 81–86. [Google Scholar] [CrossRef]
- Tertuliano, M.; Calatayud, P.; Le Rü, B. Seasonal changes of secondary compounds in the phloem sap of cassava in relation to fertilisation and to infestation by the cassava mealybug. Insect Sci. Its Appl. 1999, 19, 91–98. [Google Scholar] [CrossRef]
- Atteyat, M.; Abu-Romman, S.; Abu-Darwish, M.; Ghabeish, I. Impact of flavonoids against woolly apple aphid, Eriosoma lanigerum (Hausmann) and its sole parasitoid, Aphelinus mali (Hald.). J. Agric. Sci. 2012, 4, 227–236. [Google Scholar] [CrossRef]
- Skrzecz, I.; Sowińska, A.; Janiszewski, W. Impact of botanical antifeedants on the development of Melolontha melolontha L. beetles. Sylwan 2014, 158, 779–786. [Google Scholar]
- Lattanzio, V.; Lattanzio, V.M.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. Phytochem. Advan. Res. 2006, 661, 23–67. [Google Scholar]
- van Helden, M.; Tjallingii, W.F. Tissue localisation of lettuce resistance to the aphid Nasonovia ribisnigri using electrical penetration graphs. Entomol. Exp. Appl. 1993, 68, 269–278. [Google Scholar] [CrossRef]
- Prado, E.; Tjallingii, W.F. Aphid activities during sieve element punctures. Entomol. Exp. Appl. 1994, 72, 157–165. [Google Scholar] [CrossRef]
- Kordan, B.; Dancewicz, K.; Wróblewska, A.; Gabryś, B. Intraspecific variation in alkaloid profile of four lupine species with implications for the pea aphid probing behaviour. Phytochem. Lett. 2012, 5, 71–77. [Google Scholar] [CrossRef]
- Paprocka, M.; Gliszczyńska, A.; Dancewicz, K.; Gabryś, B. Novel hydroxy- and epoxy-cis-jasmone and dihydrojasmone derivatives affect the foraging of the peach potato aphid Myzus persicae (Sulzer) (Homoptera: Aphididae). Molecules 2018, 23, 2362. [Google Scholar] [CrossRef]
- Mayoral, A.M.; Tjallingii, W.F.; Castañera, P. Probing behaviour of Diuraphis noxia on five cereal species with different hydroxamic acid levels. Entomol. Exp. Appl. 1996, 78, 341–348. [Google Scholar] [CrossRef]
- Gabryś, B.; Dancewicz, K.; Gliszczyńska, A.; Kordan, B.; Wawrzeńczyk, C. Systemic deterrence of aphid probing and feeding by novel β-damascone analogues. J. Pest. Sci. 2015, 88, 507–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fereres, A.; Moreno, A. Behavioural aspects influencing plant virus transmission by homopteran insects. Virus Res. 2009, 141, 158–168. [Google Scholar] [CrossRef]
- Wensler, R.J.D.; Filshie, B.K. Gustatory sense organs in the food canal of aphids. J. Morphol. 1969, 129, 473–492. [Google Scholar] [CrossRef]
- Ponsen, M.B. Alimentary tract. In Aphids, Their Biology, Natural Enemies and Control, 1st ed.; Minks, A.K., Harrewijn, P., Eds.; Elsevier: Amsterdam, The Netherland, 1987; Volume A, pp. 79–97. [Google Scholar]
- Martin, B.; Collar, J.L.; Tjallingii, W.F.; Fereres, A. Intracellular ingestion and salivation by aphids may cause the acquisition and inoculation of non-persistently transmitted plant viruses. J. Gen. Virol. 1997, 78, 2701–2705. [Google Scholar] [CrossRef]
- Garzo, E.; Moreno, A.; Plaza, M.; Fereres, A. Feeding behavior and virus-transmission ability of insect vectors exposed to systemic insecticides. Plants 2020, 9, 895. [Google Scholar] [CrossRef]
- Grudniewska, A.; Dancewicz, K.; Białońska, A.; Wawrzeńczyk, C.; Gabryś, B. Piperitone-derived saturated lactones: Synthesis and aphid behavior-modifying activity. J. Agric. Food Chem. 2013, 61, 3364–3372. [Google Scholar] [CrossRef] [PubMed]
- Dancewicz, K.; Sznajder, K.; Załuski, D.; Kordan, B.; Gabryś, B. Behavioral sensitivity of Myzus persicae to volatile isoprenoids in plant tissues. Entomol. Exp. Appl. 2016, 160, 229–240. [Google Scholar] [CrossRef]
- Stec, K.; Kozłowska, J.; Wróblewska-Kurdyk, A.; Kordan, B.; Anioł, M.; Gabryś, B. Effect of naringenin and its derivatives on the probing behavior of Myzus persicae (Sulz.). Molecules 2020, 25, 3185. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Brunetti, C.; Fini, A.; Sebastiani, F.; Gori, A.; Tattini, M. Modulation of phytohormone signaling: A primary function of flavonoids in plant–environment interactions. Front. Plant Sci. 2018, 9, 1042. [Google Scholar] [CrossRef] [Green Version]
- Scarano, A.; Chieppa, M.; Santino, A. Looking at flavonoid biodiversity in horticultural crops: A colored mine with nutritional benefits. Plants 2018, 7, 98. [Google Scholar] [CrossRef] [Green Version]
- Kianersi, F.; Abdollahi, M.R.; Mirzaie-asl, A.; Dastan, D.; Rasheed, F. Identification and tissue-specific expression of rutin biosynthetic pathway genes in Capparis spinosa elicited with salicylic acid and methyl jasmonate. Sci. Rep. 2020, 10, 8884. [Google Scholar] [CrossRef]
- Marsafari, M.; Samizadeh, M.; Rabiei, B.; Mehrabi, A.; Koffas, M.; Xu, P. Biotechnological production of flavonoids: An update on plant metabolic engineering, microbial host selection, and genetically encoded biosensors. Biotechnol. J. 2020, 15, 1900432. [Google Scholar] [CrossRef]
- Mitchell, C.; Brennan, R.M.; Graham, J.; Karley, A.J. Plant defense against herbivorous pests: Exploiting resistance and tolerance traits for sustainable crop protection. Front. Plant Sci. 2016, 7, 1132. [Google Scholar] [CrossRef] [Green Version]
- Riddick, E.W. Potential of quercetin to reduce herbivory without disrupting natural enemies and pollinators. Agriculture 2021, 11, 476. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Aphid-plant interactions: What goes on in the depth of the tissue? Proc. Sect. Exp. Appl. Entomol. Neth. Entomol. Soc. 1995, 6, 163–169. [Google Scholar]
- Hardie, J.; Holyoak, M.; Taylor, N.J.; Griffiths, D.C. The combination of electronic monitoring and video-assisted observations of plant penetration by aphids and behavioural effects of polygodial. Èntomol. Exp. Appl. 1992, 62, 233–239. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| EPG Variable 1 | Control | Quercetin | Rutin | ||
|---|---|---|---|---|---|
| 0.1% | 0.5% | 0.1% | 0.5% | ||
| General aspects | n = 22 | n = 14 | n = 11 | n = 19 | n = 14 |
| Total duration of no-probing (h) 2 | 0.4 ± 0.1 a,A | 0.4 ± 0.2 a | 0.3 ± 0.1 a | 0.4 ± 0.1 A | 0.6 ± 0.1 A |
| Total duration of pathway C + F + G (h) 2 | 2.9 ± 0.3 a,A | 3.2 ± 0.5 a | 3.2 ± 0.6 a | 4.6 ± 0.3 A | 4.7 ± 0.4 A |
| Total duration of phloem phase E1 + E2 (h) 2 | 4.7 ± 0.3 a,A | 4.4 ± 0.6 a | 4.5 ± 0.6 a | 2.9 ± 0.4 A | 2.7 ± 0.4 A |
| Phloem phase index (E1 + E2) / (C + E1 + E2 + G + F) 2 | 0.62 ± 0.04 a,A | 0.56 ± 0.08 a | 0.58 ± 0.08 a | 0.38 ± 0.05 A | 0.37 ± 0.05 A |
| Number of probes 2 | 15.5 ± 1.9 a,A | 12.7 ± 2.4 a,b | 8.1 ± 2.6 b | 14.6 ± 1.7 A | 15.9 ± 1.7 A |
| Mean duration of a probe (h) 2 | 0.8 ± 0.2 a,A | 1.6 ± 0.6 a,b | 1.7 ± 0.3 b | 0.8 ± 0.2 A | 0.6 ± 0.1 A |
| Probing in non-phloem tissues before first phloem phase | n = 22 | n = 13 | n = 10 | n = 19 | n = 13 |
| Number of probes before first phloem phase 3 | 5.0 ± 1.0 a,A | 6.0 ± 1.4 a | 4.0 ± 2.0 a | 5.4 ± 1.2 A | 6.2 ± 1.5 A |
| Duration of first probe (min) 3 | 31.9 ± 12.7 a,A | 45.9 ± 36.2 a | 37.6 ± 22.4 a | 26.4 ± 12.1 A | 23.2 ± 11.4 A |
| Time from first probe to first phloem phase (h) 3 | 1.1 ± 0.2 a,A | 1.4 ± 0.2 a | 1.4 ± 0.4 a | 1.9 ± 0.4 A | 2.3 ± 0.6 A |
| Time from first probe to first sustained sap ingestion phase E2 > 10 min (h) 4 | 1.1 ± 0.2 a,A | 1.4 ± 0.2 a | 1.8 ± 0.4 a | 2.0 ± 0.4 A,B | 2.7 ± 0.5 B |
| Probing in phloem tissues | n = 22 | n = 13 | n = 10 | n = 19 | n = 13 |
| Duration of first phloem phase E1 + E2 (h) 3 | 1.5 ± 0.4 a,A | 2.8 ± 0.2 a | 2.9 ± 0.8 a | 1.4 ± 0.3 A | 1.1 ± 0.2 A |
| Mean duration of phloem sap ingestion phase E2 (h) 3 | 1.4 ± 0.4 a,A | 1.9 ± 0.6 a | 2.0 ± 0.4 a | 0.8 ± 0.1 A | 1.0 ± 0.2 A |
| Phloem salivation index E1 / (E1 + E2) 3 | 0.04 ± 0.01 a,A | 0.02 ± 0.01 a | 0.02 ± 0.01 a | 0.07 ± 0.04 A | 0.04 ± 0.01 A |
| EPG Variable 1 | Control | Quercetin | Rutin | ||
|---|---|---|---|---|---|
| 0.1% | 0.5% | 0.1% | 0.5% | ||
| General aspects | n = 19 | n = 15 | n = 14 | n = 14 | n = 15 |
| Total duration of no-probing (h) 2 | 0.9 ± 0.2 a,A | 1.1 ± 0.3 a | 1.2 ± 0.4 a | 1.2 ± 0.2 A,B | 2.5 ± 0.4 B |
| Total duration of pathway C + F + G (h) 2 | 3.7 ± 0.4 a,A | 2.9 ± 0.3 a | 3.9 ± 0.5 a | 4.0 ± 0.5 A | 3.6 ± 0.4 A |
| Total duration of phloem phase E1 + E2 (h) 2 | 3.4 ± 0.5 a,A | 4.0 ± 0.5 a | 2.8 ± 0.7 a | 2.8 ± 0.6 a | 1.8 ± 0.6 A |
| Phloem phase index 2 (E1 + E2) / (C + E1 + E2 + G + F) | 0.46 ± 0.07 a,A | 0.56 ± 0.06 a | 0.37 ± 0.09 a | 0.39 ± 0.08 A | 0.28 ± 0.08 A |
| Number of probes 2 | 27.1 ± 4.6 a,A | 27.5 ± 4.0 a | 26.4 ± 3.5 a | 32.4 ± 5.3 A,B | 43.9 ± 5.3 B |
| Mean duration of a probe (h) 2 | 0.6 ± 0.2 a,A | 0.5 ± 0.1 a | 0.4 ± 0.1 a | 0.3 ± 0.1 A,B | 0.2 ± 0.1 B |
| Probing in non-phloem tissues before first phloem phase | |||||
| Number of probes before first phloem phase 3 | 13.2 ± 2.3 a,A n = 18 | 14.2 ± 3.1 a n = 15 | 10.7 ± 2.5 a n = 12 | 19.1 ± 4.1 A,B n = 12 | 31.3 ± 6.0 B n = 12 |
| Duration of first probe (min) 3 | 21.2 ± 14.8 a,A n = 18 | 1.0 ± 0.2 a n = 15 | 22.8 ± 19.4 a n = 12 | 1.5 ± 0.5 A,B n = 12 | 0.4 ± 0.1 B n = 12 |
| Time from first probe to first phloem phase (h) 3 | 3.0 ± 0.5 a,A n = 18 | 2.1 ± 0.4 a n = 15 | 1.9 ± 0.4 a n = 12 | 3.1 ± 0.7 A n = 12 | 3.3 ± 0.6 A n = 12 |
| Time from first probe to first sustained sap ingestion phase E2 > 10 min (h) 4 | 3.3 ± 0.6 a,A n = 17 | 2.3 ± 0.4 a n = 15 | 2.1 ± 0.5 a n = 10 | 3.9 ± 0.8 A n = 12 | 4.0 ± 0.7 A n = 10 |
| Total duration of no-probing before first phloem phase (h) 3 | 0.4 ± 0.1 a,A n = 18 | 0.5 ± 0.2 a n = 15 | 0.3 ± 0.1 a n = 12 | 0.7 ± 0.2 A,B n = 12 | 1.3 ± 0.2 B n = 12 |
| Probing in phloem tissues2 | n = 18 | n=15 | n=12 | n = 12 | n = 12 |
| Duration of first phloem phase E1 + E2 (h) 3 | 2.8 ± 0.6 a,A | 2.2 ± 0.6 a | 2.2 ± 0.7 a | 1.5 ± 0.5 A | 1.5 ± 0.7 A |
| Mean duration of phloem sap ingestion phase E2 (h) 3 | 3.1 ± 0.5 a,A | 2.2 ± 0.6 a | 2.2 ± 0.7 a | 2.0 ± 0.5 A | 1.7 ± 0.7 A |
| Phloem salivation index E1 / (E1 + E2) 3 | 0.08 ± 0.04 a,A | 0.03 ± 0.02 a | 0.04 ± 0.01 a | 0.02 ± 0.01 A | 0.07 ± 0.02 A |
| EPG Variable 1 | Control | Quercetin | Rutin | ||
|---|---|---|---|---|---|
| 0.1% | 0.5% | 0.1% | 0.5% | ||
| General aspects | n = 16 | n = 13 | n = 16 | n = 15 | n = 13 |
| Total duration of no-probing (h) 2 | 0.6 ± 0.1 a,A | 1.0 ± 0.2 a | 1.0 ± 0.2 a | 0.9 ± 0.3 A | 0.9 ± 0.3 A |
| Total duration of pathway C + F + G (h) 2 | 4.3 ± 0.5 a,A | 4.9 ± 0.4 a | 4.8 ± 0.4 a | 4.2 ± 0.5 A | 4.3 ± 0.6 A |
| Total duration of phloem phase E1 + E2 (h) 2 | 3.1 ± 0.6 a,A | 2.1 ± 0.4 a | 2.2 ± 0.5 a | 2.9 ± 0.5 A | 2.7 ± 0.7 A |
| Phloem phase index 2 (E1 + E2) / (C + E1 + E2 + G + F) | 0.49 ± 0.07 a,A | 0.30 ± 0.06 a | 0.30 ± 0.06 a | 0.39 ± 0.07 A | 0.36 ± 0.09 A |
| Number of probes 2 | 8.8 ± 1.6 a,A | 12.1 ± 1.7 a | 9.0 ± 1.4 a | 7.9 ± 1.3 A | 8.3 ± 1.5 A |
| Mean duration of a probe (h) 2 | 1.3 ± 0.2 a,A | 0.7 ± 0.1 a | 1.6 ± 0.5 a | 1.7 ± 0.5 A | 1.6 ± 0.6 A |
| Probing in non-phloem tissues before first phloem phase | |||||
| Number of probes before first phloem phase 3 | 2.4 ± 0.5 a,A n = 15 | 2.8 ± 0.8 a n = 12 | 2.5 ± 0.8 a n = 15 | 1.8 ± 0.4 A n = 12 | 2.5 ± 0.9 A n = 13 |
| Duration of first probe (m) 3 | 69.4 ± 28.7 a,A n = 15 | 40.9 ± 9.3 a n = 12 | 89.2 ± 35.8 a n = 15 | 96.6 ± 45.0 A n = 12 | 104.0 ± 37.3 A n = 13 |
| Time from first probe to first phloem phase (h) 3 | 1.6 ± 0.3 a,A n = 15 | 1.7 ± 0.4 a n = 12 | 2.3 ± 0.5 a n = 15 | 2.1 ± 0.6 A n = 12 | 1.6 ± 0.5 A n = 13 |
| Time from first probe to first sustained sap ingestion phase E2 > 10 min (h) 4 | 2.9 ± 0.5 a,A n = 12 | 2.8 ± 0.7 a n = 10 | 4.1 ± 0.5 a n = 13 | 2.2 ± 0.3 A n = 11 | 2.8 ± 0.5 A n = 11 |
| Total duration of no-probing before first phloem phase (h) 3 | 0.2 ± 0.04 a,A n = 15 | 0.2 ± 0.1 a n = 12 | 0.2 ± 0.1 a n = 15 | 0.3 ± 0.2 A n = 12 | 0.4 ± 0.2 A n = 13 |
| Probing in phloem tissues | n = 15 | n = 12 | n = 15 | n = 12 | n = 13 |
| Duration of first phloem phase E1 + E2 (h) 3 | 2.1 ± 0.7 a,A | 0.7 ± 0.3 a | 0.5 ± 0.2 a | 0.8 ± 0.4 A | 1.4 ± 0.8 A |
| Mean duration of phloem sap ingestion phase E2 (h) 3 | 1.7 ± 0.6 a,A | 0.9 ± 0.3 a | 0.7 ± 0.2 a | 1.5 ± 0.4 A | 1.7 ± 0.7 A |
| Phloem salivation index E1 / (E1 + E2) 3 | 0.17 ± 0.09 a,A | 0.07 ± 0.03 a | 0.12 ± 0.06 a | 0.09 ± 0.08 A | 0.08 ± 0.04 A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stec, K.; Kordan, B.; Gabryś, B. Quercetin and Rutin as Modifiers of Aphid Probing Behavior. Molecules 2021, 26, 3622. https://doi.org/10.3390/molecules26123622
Stec K, Kordan B, Gabryś B. Quercetin and Rutin as Modifiers of Aphid Probing Behavior. Molecules. 2021; 26(12):3622. https://doi.org/10.3390/molecules26123622
Chicago/Turabian StyleStec, Katarzyna, Bożena Kordan, and Beata Gabryś. 2021. "Quercetin and Rutin as Modifiers of Aphid Probing Behavior" Molecules 26, no. 12: 3622. https://doi.org/10.3390/molecules26123622
APA StyleStec, K., Kordan, B., & Gabryś, B. (2021). Quercetin and Rutin as Modifiers of Aphid Probing Behavior. Molecules, 26(12), 3622. https://doi.org/10.3390/molecules26123622