
Bioefficacy of Epaltes divaricata (L.) n-Hexane Extracts and Their Major Metabolites against the Lepidopteran Pests Spodoptera litura (fab.) and Dengue Mosquito Aedes aegypti (Linn.)



 , , , ,
, , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Methodology
2.1. Insect Culture
2.2. Plant Harvesting and Crude Extract Preparation
2.3. Characterization of Plant Volatile through GC–MS
2.4. Chemicals
2.5. Larvicidal Bioassay
2.5.1. A. aegypti Larvicidal Assay
2.5.2. S. litura Larvicidal Assay
2.6. Larval and Pupal Duration Assay
2.7. Enzyme Assays
2.7.1. α- and β-Carboxylesterase Activity
2.7.2. GST and CYP450 Activity
2.8. Gut-Histological Assay
2.9. Data Analysis
3. Results
3.1. Chemical Characterization of NH-EDx
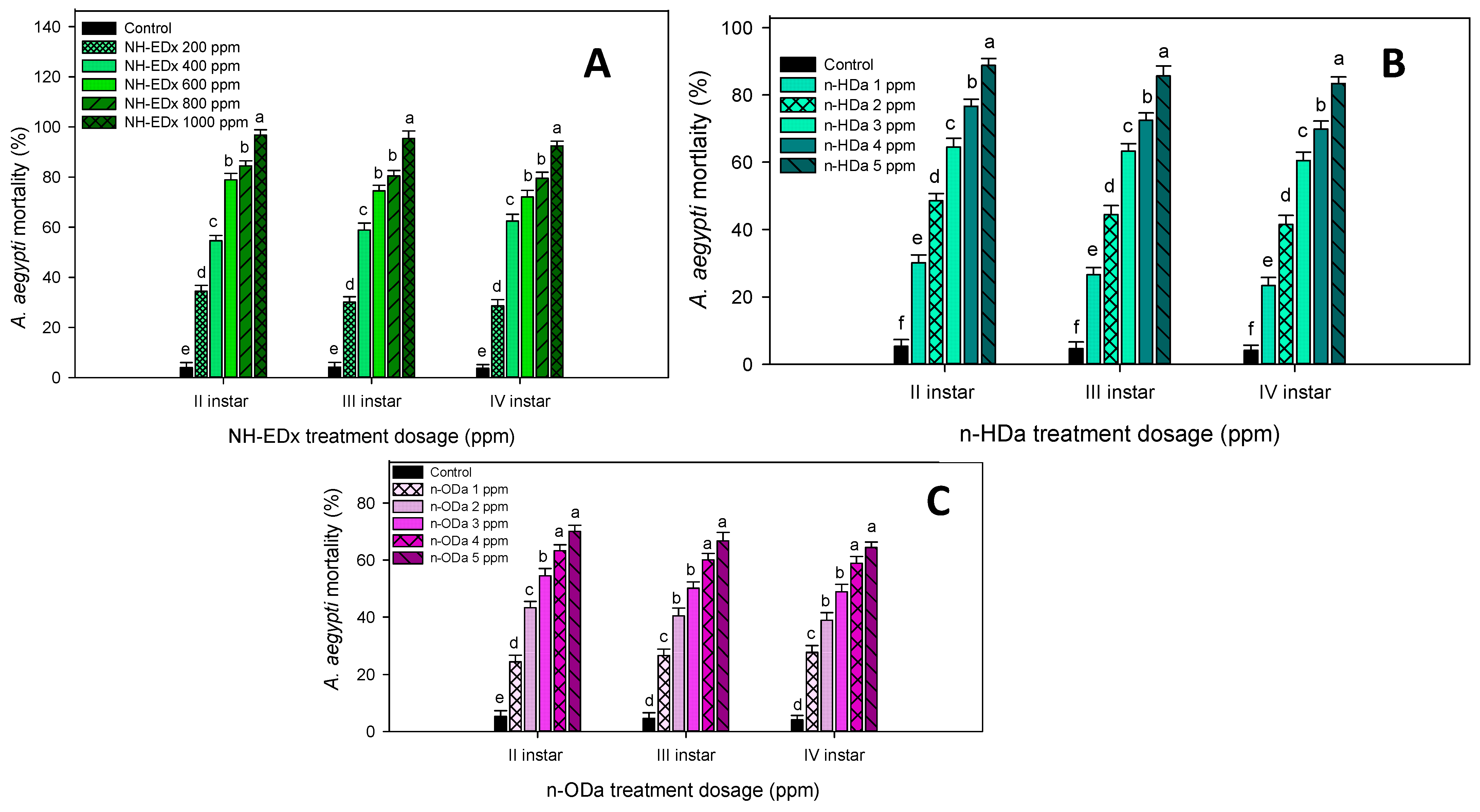
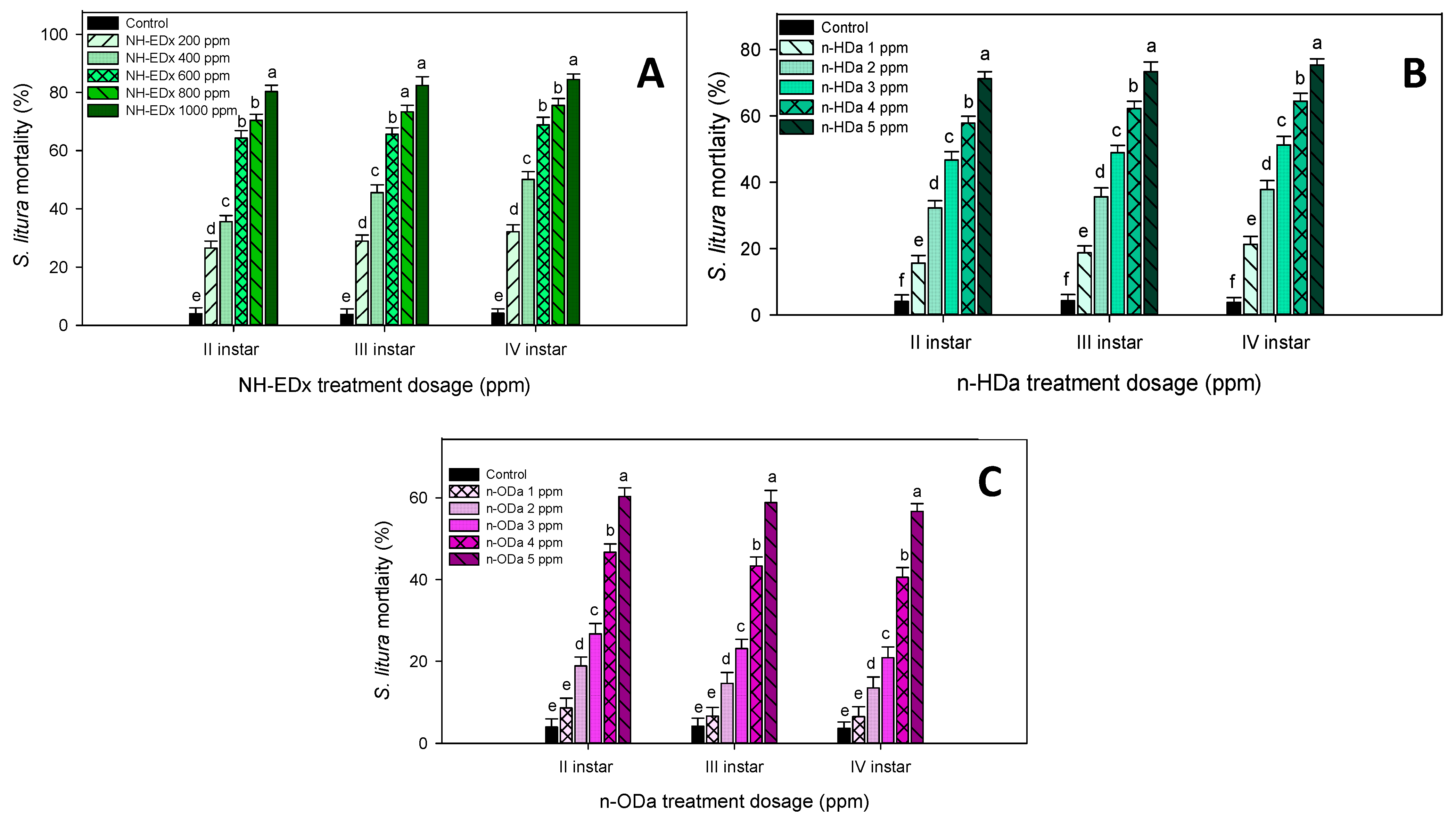
3.2. Larvicidal Activity
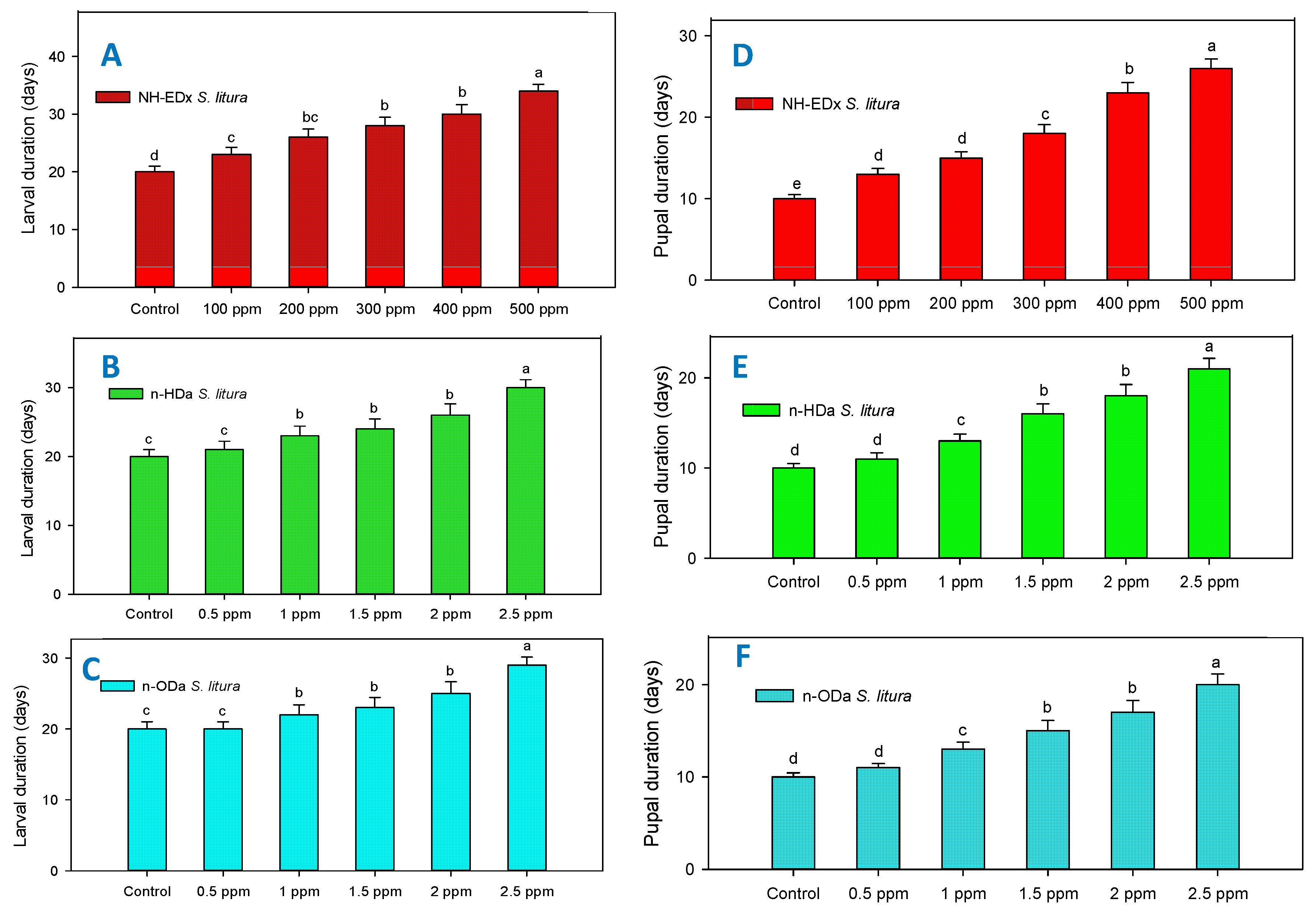
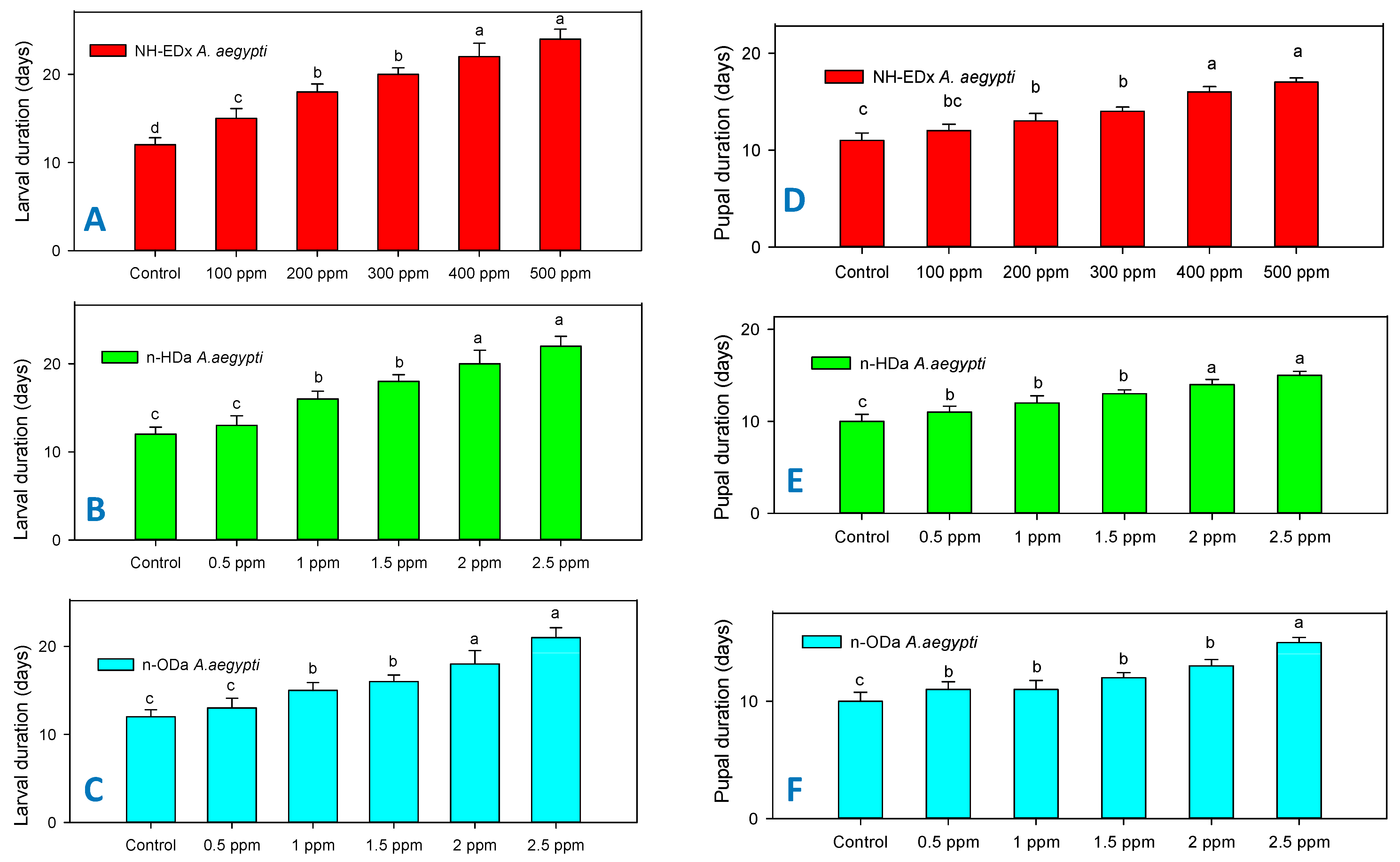
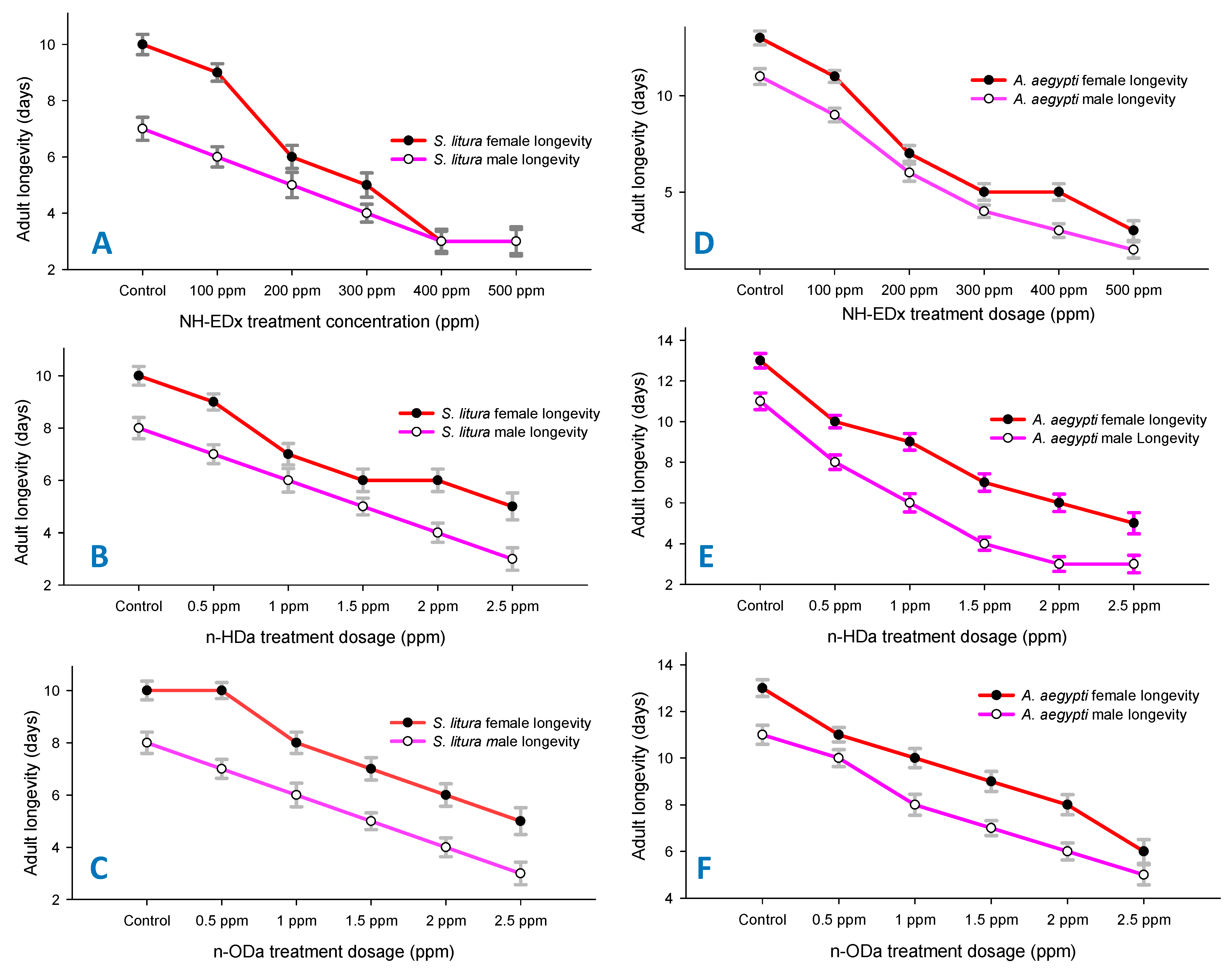
3.3. Developmental Changes
3.4. Enzyme Activity
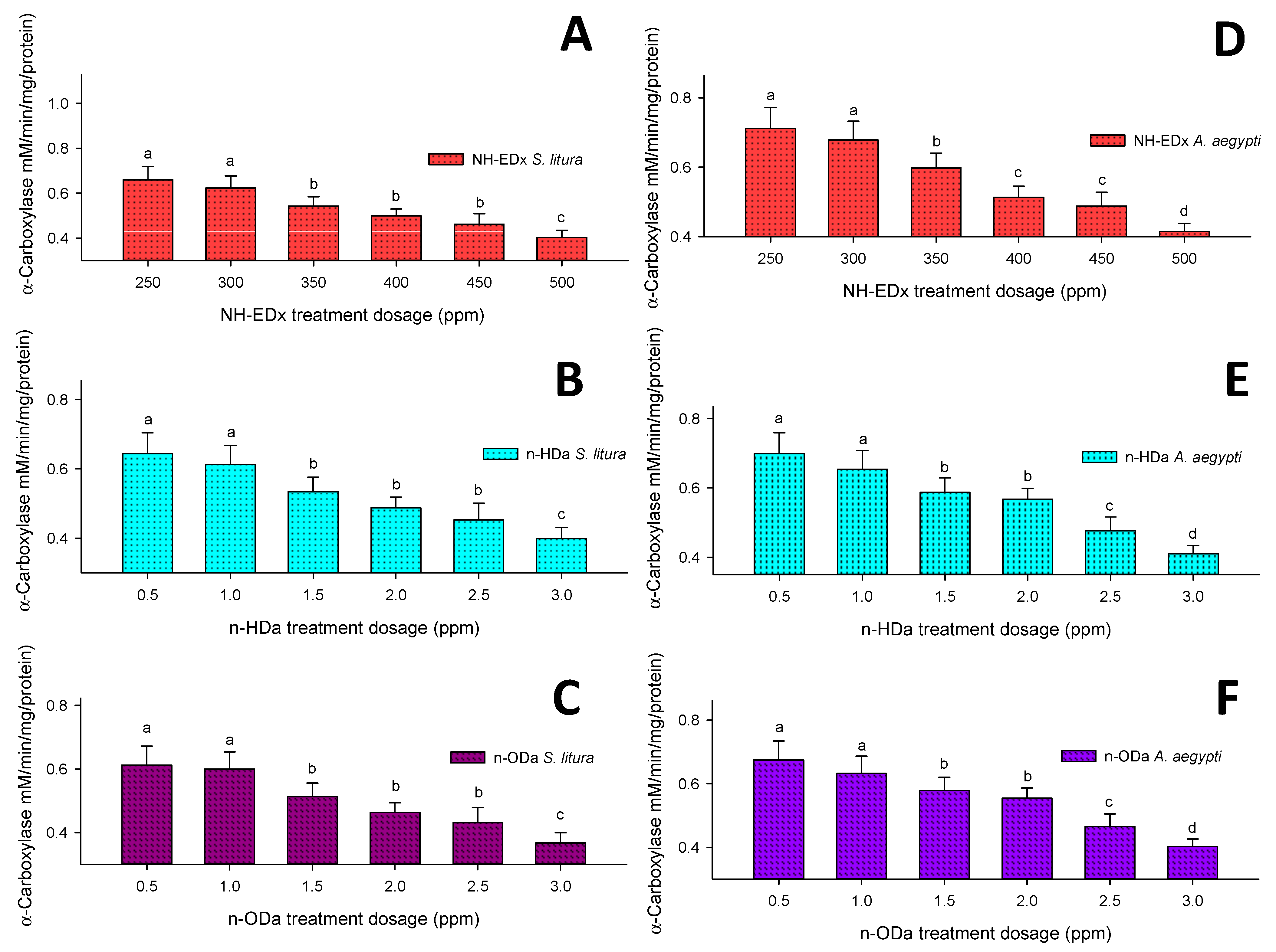
3.4.1. α-Carboxylesterase
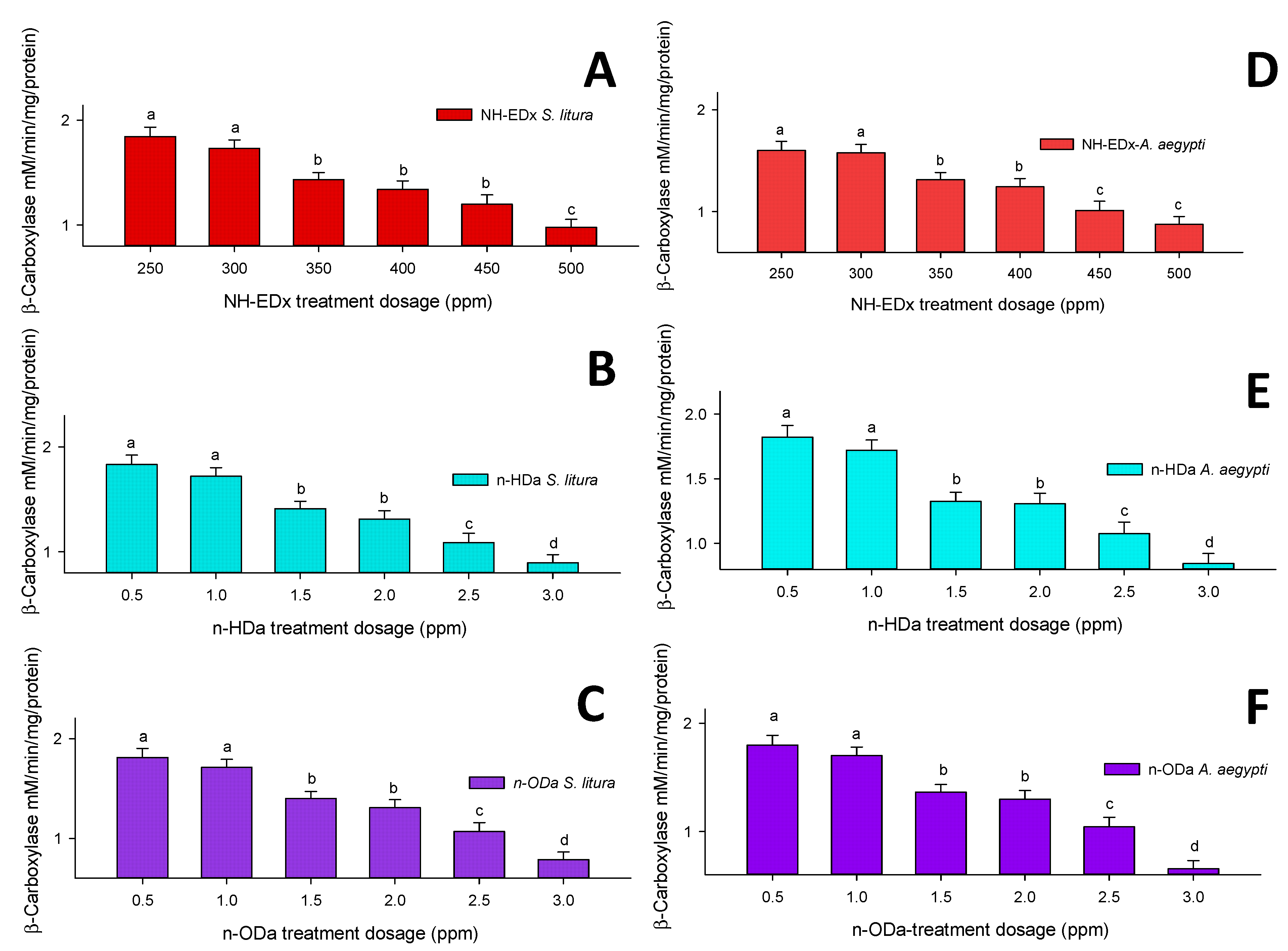
3.4.2. β-Carboxylesterase
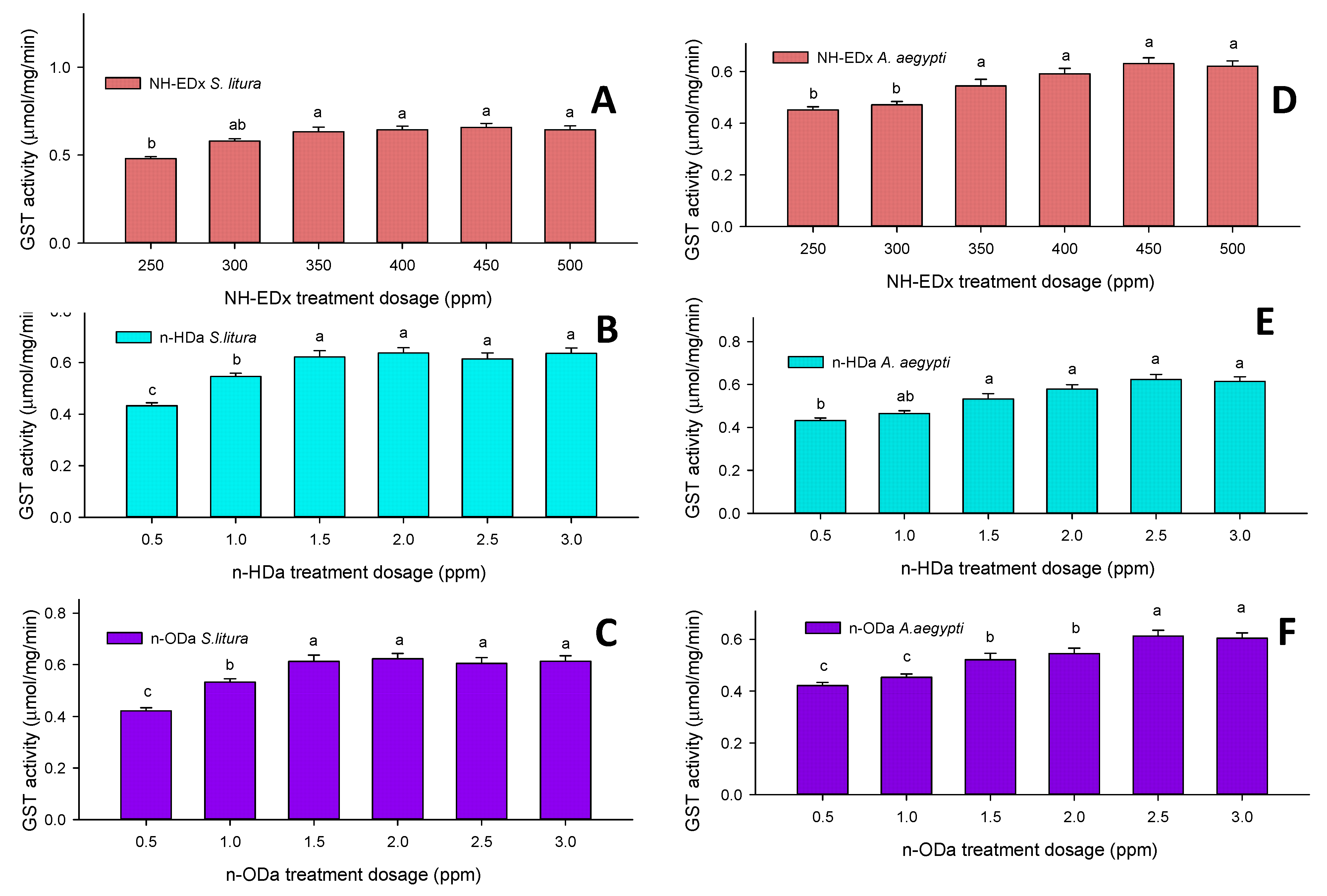
3.4.3. Glutathione S-Transferase (GST)
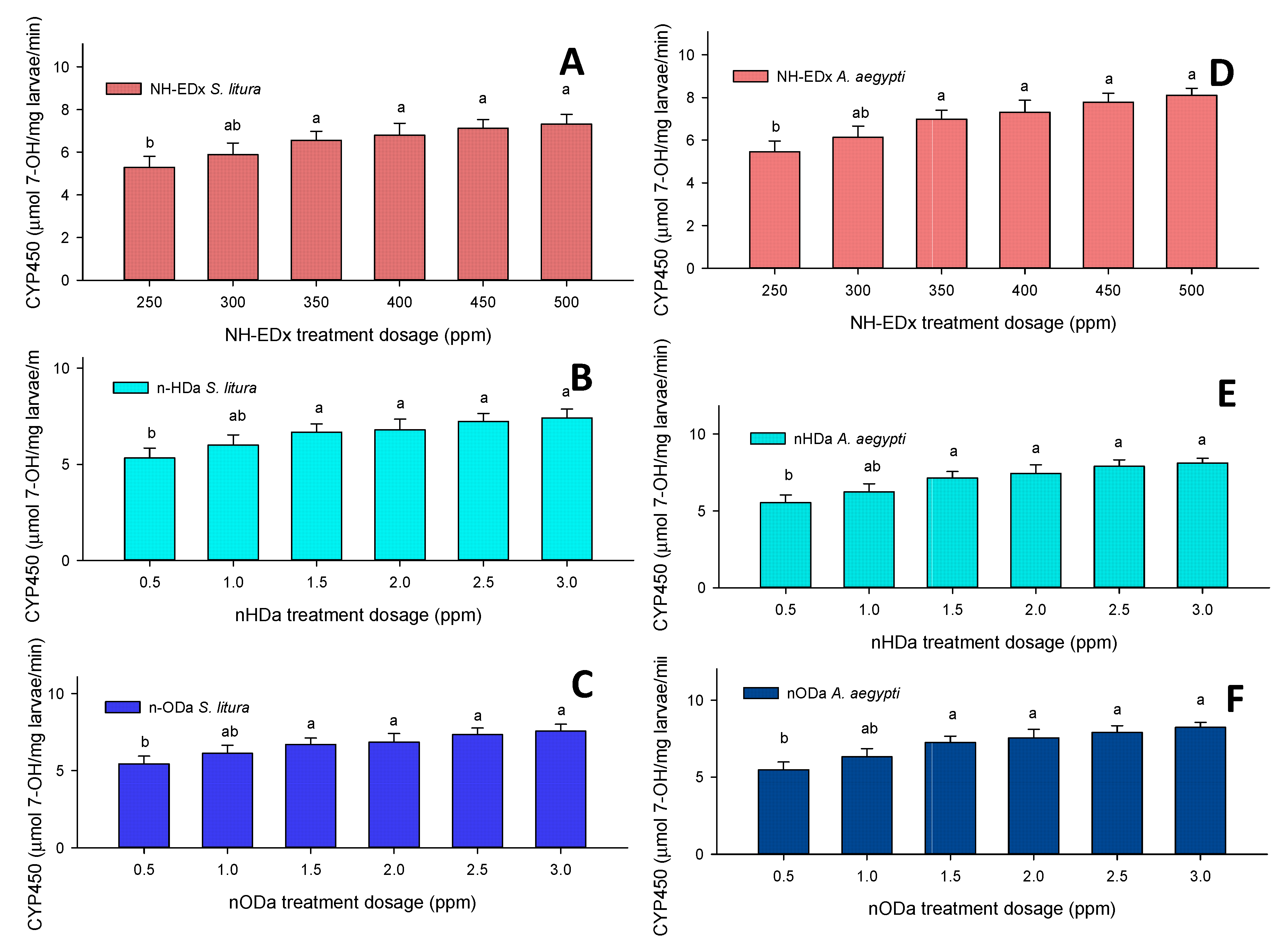
3.4.4. Cytochrome P-450 (CYP450)
3.5. Gut-Histological Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Senthil-Nathan, S. Physiological and biochemical effect of Neem and other Meliaceae plants secondary metabolites against Lepidopteran insects. Front. Physiol. 2013, 4, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Murfadunnisa, S.; Vasantha-Srinivasan, P.; Ganesan, R.; Senthil-Nathan, S.; Kim, T.J.; Ponsankar, A.; Kumar, S.D.; Chandramohan, D.; Krutmuang, P. Larvicidal and enzyme inhibition of essential oil from Spheranthus amaranthroids (Burm.) against lepidopteran pest Spodoptera litura (Fab.) and their impact on non-target earthworms. Biocatal. Agri. Biotechnol. 2019, 21, 101324. [Google Scholar] [CrossRef]
- Karthi, S.; Uthirarajan, K.; Manohar, V.; Venkatesan, M.; Chinnaperumal, K.; Vasantha-Srinivasan, P.; Krutmuang, P. Larvicidal enzyme inhibition and repellent activity of red mangrove Rhizophora mucronata (Lam.) leaf extracts and their biomolecules against three medically challenging arthropod vectors. Molecules 2020, 25, 3844. [Google Scholar] [CrossRef]
- Ponsakar, A.; Sahayaraj, K.; Senthil-Nathan, S.; Vasantha-Srinivasan, P.; Karthi, S.; Thanigaivel, A.; Petchidurai, G.; Madasamy, M.; Hunter, W.B. Toxicity and developmental effect of cucurbitacin E from Citrullus colocynthis L. (Cucurbitales: Cucurbitaceae) against Spodoptera litura Fab. and a non-target earthworm Eisenia fetida Savigny. Environ. Sci. Pollut. Res. 2020, 27, 23390–23401. [Google Scholar] [CrossRef]
- Chellappandian, M.; Senthil-Nathan, S.; Vasantha-Srinivasan, P.; Karthi, S.; Thanigaivel, A.; Kalaivani, K.; Sivanesh, H.; Stanley-Raja, V.; Chanthini, K.M.; Shyam-Sundar, N. Target and non-target botanical pesticides effect of Trichodesma indicum(Linn) R. Br. and their chemical derivatives against the dengue vector, Aedes aegypti L. Environ. Sci. Pollut. Res. 2019, 26, 16303–16315. [Google Scholar] [CrossRef]
- Senthil-Nathan, S.; Choi, M.Y.; Paik, C.H.; Seo, H.Y.; Kalivani, K.; Kim, J.D. Effect of azadirachtin on acetylcholinesterase (AChE) activity and histology of the brown plant hopper Nilaparvata lugens (Stal). Ecotoxicol. Environ. Saf. 2008, 70, 244–250. [Google Scholar] [CrossRef]
- Thanigaivel, A.; Chandrasekaran, R.; Revathi, K.; Nisha, S.; Sathish-Narayanan, S.; Kirubakaran, S.A.; Senthil-Nathan, S. Larvicidal efficacy of Adhatoda vasica (L.) Nees against the bancroftian filariasis vector Culex quinquefasciatus Say and dengue vector Aedes aegypti L. in in vitro condition. Parasitol. Res. 2012, 110, 1993–1999. [Google Scholar] [CrossRef] [PubMed]
- Lija-Escaline, J.; Senthil-Nathan, S.; Thanigaivel, A.; Pradeepa, V.; Vasantha-Srinivasan, P.; Ponsankar, A.; Edwin, E.; Selin-Rani, S.; Abdel-Megeed, A. Physiological and biochemical effects of chemical constituents from Piper nigrum Linn (Piperaceae) against the dengue vector Aedes aegypti Liston (Diptera: Culicidae). Parasitol. Res. 2015, 114, 4239–4249. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G. Plant-mediated biosynthesis of nanoparticles as an emerging tool against mosquitoes of medical and veterinary importance: A review. Parasitol. Res. 2016, 115, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Senthil-Nathan, S. A review of bio pesticides and their mode of action against insect pests. In Environmental Sustainability- Role of Green Technologies; Springer: Bangalore, India, 2015; pp. 49–63. [Google Scholar]
- Selin-Rani, S.; Senthil-Nathan, S.; Thanigaivel, A.; Vasantha-Srinivasan, P.; Edwin, E.; Ponsankar, A.; Lija-Escaline, J.; Kalaivani, K.; Abdel-Megeed, A.; Hunter, W.B.; et al. Toxicity and physiological effect of quercetin on generalist herbivore, Spodoptera litura Fab. and a non-target earthworm Eisenia fetida Savigny. Chemosphere 2016, 165, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Senthil-Nathan, S. A review of resistance mechanisms of synthetic insecticides and botanicals, phytochemicals, and essential oils as alternative larvicidal agents against mosquitoes. Front. Physiol. 2020, 10, 1591. [Google Scholar] [CrossRef] [PubMed]
- Amala, K.; Ganesan, R.; Karthi, S.; Senthil-Nathan, S.; Chellappandian, M.; Krutmunag, P.; Radhakrishnan, R.; Mohammad, F.; Ponsankar, A.; Vasantha-Srinivasan, P. Larval and gut enzyme toxicity of n-hexane extract Epaltes pygmaea DC. against the arthropod vectors and its non-toxicity against aquatic predator. Toxin Rev. 2020. [Google Scholar] [CrossRef]
- Huang, S.H.; Xian, J.D.; Kong, S.Z.; Li, Y.C.; Xie, J.H.; Lin, J.; Chen, J.N.; Wang, H.F.; Su, Z.R. Insecticidal activity of pogostone against Spodoptera litura and Spodoptera exigua (Lepidoptera: Noctuidae). Pest. Manag. Sci. 2014, 70, 510–516. [Google Scholar] [CrossRef]
- Selin-Rani, S.; Senthil-Nathan, S.; Revathi, K.; Chandrasekaran, R.; Thanigaivel, A.; Vasantha-Srinivasan, P.; Ponsankar, A.; Edwin, E.; Pradeepa, V. Toxicity of Alangium salvifolium Wang chemical constituents against the tobacco cutworm Spodoptera litura Fab. Pest. Biochem. Physiol. 2016, 126, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.A.; Isman, M.B. Anti-feedant and toxic activity of Trichilia americana extract against the larvae of Spodoptera litura. Entomol. Exp. Appl. 2001, 98, 9–16. [Google Scholar] [CrossRef]
- Koou, S.; Chong, C.; Vythilingam, I.; Lee, C.; Ng, L. Insecticide resistance and its underlying mechanisms in field populations of Aedes aegypti adults (Diptera: Culicidae) in Singapore. Parasit. Vectors 2014, 7, 471. [Google Scholar] [CrossRef]
- Reegan, A.D.; Gandhi, M.R.; Paulraj, M.G.; Ignacimuthu, S. Ovicidal and oviposition deterrent activities of medicinal plant extracts against Aedes aegypti L. and Culex quinquefasciatus Say mosquitoes (Diptera: Culicidae). Osong Public Health Res. Perspect. 2015, 6, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Thanigaivel, A.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Edwin, E.; Ponsankar, A.; Chellappandian, M.; Selin-Rani, S.; Lija-Escaline, J.; Kalaivani, K. Impact of Terminalia chebula Retz. against Aedes aegypti L. and non-target aquatic predatory insects. Ecotoxicol. Environ. Saf. 2016, 137, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Sivan, A.; Shriram, A.N.; Sunish, I.P.; Vidhya, P.T. Studies on insecticide susceptibility of Aedes aegypti (Linn) and Aedes albopictus (Skuse) vectors of dengue and chikungunya in Andaman and Nicobar Islands, India. Parasitol. Res. 2015, 114, 4693–4702. [Google Scholar] [CrossRef]
- Senthil-Nathan, S.; Kalaivani, K. Combined effects of azadirachtin and nucleopolyhedrovirus (SpltNPV) on Spodoptera litura Fabricius (Lepidoptera: Noctuidae) larvae. Biol. Control. 2006, 39, 96–104. [Google Scholar] [CrossRef]
- Senthil-Nathan, S.; Kalaivani, K.; Sehoon, K. Effects of Dysoxylum malabaricum Bedd. (Meliaceae) extract on the malarial vector Anopheles stephensi Liston (Diptera: Culicidae). Bioresour. Technol. 2006, 97, 2077–2083. [Google Scholar] [CrossRef]
- Yogarajalakshmi, P.; Poonguzhali, T.N.; Ganesan, R.; Karthi, S.; Senthil-Nathan, S.; Krutmuang, P.; Radhakrishnan, N.; Mohammad, F.; Kim, T.; Vasantha-Srinivasan, S. Toxicological screening of marine red algae Champia parvula (C. Agardh) against the dengue mosquito vector Aedes aegypti (Linn.) and its non-toxicity against three beneficial aquatic predators. Aquat. Toxicol. 2020, 222, 105474. [Google Scholar] [CrossRef]
- Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Ponsankar, A.; Thanigaivel, A.; Edwin, E.S.; Selin-Rani, S.; Chellappandian, M.; Pradeepa, V.; Lija-Escaline, J.; Kalaivani, K.; et al. Comparative analysis of mosquito (Diptera: Culicidae: Aedes aegypti Liston) responses to the insecticide Temephos and plant derived essential oil derived from Piper betle L. Ecotoxicol. Environ. Saf. 2017, 139, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Thanigaivel, A.; Edwin, E.S.; Ponsankar, A.; Selin-Rani, S.; Pradeepa, V.; Sakthi-Bhagavathy, M.; Kalaivani, K.; Hunter, W.B.; et al. Developmental response of Spodoptera litura Fab. to treatments of crude volatile oil from Piper betle L. and evaluation of toxicity to earthworm, Eudrilus eugeniae Kinb. Chemosphere 2016, 155, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Edwin, E.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Thanigaivel, A.; Ponsankar, A.; Selin-Rani, S.; Kalaivani, K.; Hunter, W.B.; Duraipandiyan, V.; Al-Dhabi, N.A. Effect of andrographolide on phosphatases activity and cytotoxicity against Spodoptera litura. Invert. Surv. J. 2016, 13, 153–163. [Google Scholar]
- Annamalai, M.; Vasantha-Srinivasan, P.; Thanigaivel, A.; Chellappandian, M.; Karthi, S.; Mayabini, J.; Guru Pirasanna Pandi, G.; Totan, A.; Murugesan, A.G.; Senthil-Nathan, S. Effect of thiamethoxam on growth, biomass of rice varieties and its specialized herbivore, Scirpophaga incertulas Walker. Physiol. Mol. Plant. Pathol. 2017. [Google Scholar] [CrossRef]
- Dinesh-Kumar, A.; Srimaan, E.; Chellappandian, M.; Vasantha-Srinivasan, P.; Karthi, S.; Annamalai, M.; Thanigaivel, A.; Ponsankar, A.; Chanthini, K.; Shyam-Sundar, N.; et al. Target and non-target response of Swietenia Mahagoni Jacq. chemical constituents against tobacco cutworm Spodoptera litura Fab. and earthworm, Eudrilus eugeniae Kinb. Chemosphere 2018, 199, 35–43. [Google Scholar] [CrossRef]
- Shin, J.; Vasantha-Srinivasan, P.; Kim, K. The multi-faceted potential of plant-derived metabolites as antimicrobial agents against multidrug-resistant pathogens. Microbial pathog. 2018, 116, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Pradeepa, V.; Senthil-Nathan, S.; Sathish-Narayanan, S.; Selin-Rani, S.; Vasantha-Srinivasan, P.; Thanigaivel, A.; Ponsankar, A.; Edwin, E.-S.; Sakthi-Bagavathy, M.; Kalaivani, K.; et al. Potential mode of action of a novel plumbagin as a mosquito repellent against the malarial vector Anopheles stephensi, (Culicidae: Diptera). Pesticide Biochem. Physiol. 2016, 134, 84–93. [Google Scholar] [CrossRef]
- Senthil-Nathan, S.; Kalaivani, K.; Murugan, K. Effects of neem limonoids on the malaria vector Anopheles stephensi Liston (Diptera: Culicidae). Acta Trop. 2005, 96, 47–55. [Google Scholar] [CrossRef]
- Senthil-Nathan, S.; Chung, P.G.; Murugan, K. Effect of bio-pesticides applied separately or together on Nutritional Indices of the Rice Leaffolder Cnaphalocrocis medinalis. Phytoparasitica 2005, 33, 187–195. [Google Scholar] [CrossRef]
- Hewawasam, R.P.; Jayatilaka, K.A.P.W.; Pathirana, C.; Muddduwa, L.K.B. Hepatoprotective effect of Epaltes divaricata extract on carbon tetrachloride induced hepatotoxixity in mice. Ind. J. Med. Res. 2004, 120, 30–34. [Google Scholar]
- Amala, K.; Saraswathy, A.; Amerjothy, S. GC-MS analysis of N-Hexane extract of Epaltes divaricata (L.) Cass. J. Pharmacog. Phytochem. 2013, 2, 33–35. [Google Scholar]
- Chah, K.F.; Eze, C.A.; Emuelosi, C.E.; Esimone, C.O. Antibacterial and wound healing properties of methanolic extracts of some Nigerian medicinal plants. J. Ethnopharmacol. 2006, 104, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Glorybai, L.; Kannan, B.K.; Arasu, M.V.; Al-Dhabi, N.A.; Agastin, P. Some biological activities of Epaltes divaricata L.—An in vitro study. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasantha-Srinivasan, P.; Chellappandian, M.; Senthil-Nathan, S.; Ponsankar, A.; Thanigaivel, A.; Karthi, S.; Edwin, E.; Selin-Rani, S.; Kalaivani, K.; Maggi, F.; et al. A novel herbal product based on Piper betle and Sphaeranthus indicus essential oils: Toxicity, repellent activity and impact on detoxifying enzymes GST and CYP450 of Aedes aegypti Liston (Diptera: Culicidae). J. Asia-Pac. Entomol. 2018, 21, 1466–1472. [Google Scholar] [CrossRef]
- World Health Organization. Instruction for Determining the Susceptibility or Resistance of Mosquito Larvae to Insecticides; WHOVBC; 1981; Volume 81, pp. 1–6. Available online: https://apps.who.int/iris/handle/10665/69615 (accessed on 10 March 2021).
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Agra-Neto, A.C.; Napoleão, T.H.; Pontual, E.V.; Santos, N.D.L.; Luz, L.A.; Oliveira, C.M.F.; Melo-Santos, M.A.V.; Coelho, L.C.B.B.; Navarro, D.M.A.F.; Paiva, P.M.G. Effect of Moringa oleifera lectins on survival and enzyme activities of Aedes aegypti larvae susceptible and resistant to organophosphate. Parasitol. Res. 2015, 113, 175–184. [Google Scholar] [CrossRef]
- Thanigaivel, A.; Senthil-Nathan, S.; Vasantha-Srinivasan, P.; Edwin, E.; Ponsankar, A.; Selin-Rani, S.; Pradeepa, V.; Chellappandian, M.; Kalaivani, K.; Abdel-Megeed, A.; et al. Chemicals isolated from Justicia adhatoda Linn reduce fitness of the mosquito. Arch. Insect Biochem. Physiol. 2017, 94, e21384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef]
- Silva, W.J.; Doria, G.A.A.; Maia, R.T.; Nunes, R.S.; Carvalho, G.A.; Blank, A.F.; Alves, P.B.; Marcal, R.M.; Cavalcanti, S.C.H. Effects of essential oils on Aedes aegypti larvae: Alternatives to environmentally safe insecticides. Bioresour. Technol. 2008, 99, 3251–3255. [Google Scholar] [CrossRef]
- Polson, K.A.; Brogdon, W.G.; Rawlins, S.C.; Chadee, D.D. Characterization of insecticide resistance in Trinidadian strains of Aedes aegypti mosquitoes. Acta Trop. 2011, 117, 31–38. [Google Scholar] [CrossRef]
- Piesik, D.; Wenda-Piesik, A.; Weaver, D.K.; Morrill, W.L. Influence of Fusarium crown rot disease on semio-chemical production by wheat plants. J. phytopathol. 2007, 155, 488–496. [Google Scholar] [CrossRef]
- Piesik, D.; Delaney, K.J.; Wenda-Piesik, A.; Sendel, S.; Tabaka, P.; Buszewski, B. Meligethes aeneus pollen-feeding suppresses, and oviposition induces, Brassica napus volatiles: Beetle attraction/repellence to lilac aldehydes and veratrole. Chemoecology 2013, 23, 241–250. [Google Scholar] [CrossRef]
- Piesik, D.; Kalka, I.; Wenda-Piesik, A.; Bocianowski, J. Apion miniatum Germ. herbivory on the mossy sorrel, Rumex confertus Willd.: Induced plant volatiles and weevil orientation responses. Pol. J. Environ. Stud. 2014, 23, 2149–2156. [Google Scholar] [CrossRef]
- Vasantha-Srinivasan, P.; Karthi, S.; Ganesan, R.; Senthil-Nathan, S.; Krutmuang, P.; Chellappandian, M.; Radhakrishnan, N.; Ponsankar, A.; Karthick, K.; Nelofer, A. The efficacy of methanolic extract of Swietenia mahagoni Jacq. (Meliaceae) and a commercial insecticide against laboratory and field strains of Aedes aegypti (Linn.) and their impact on its predator Toxorhnchites splendens. Biocatal. Agri. Biotechnol. 2021, 31, 101915. [Google Scholar] [CrossRef]
- Thanigaivel, A.; Chanthini, M.; Karthi, S.; Vasantha-Srinivasan, P.; Ponsakar, A.; Sivanesh, H.; Stanley-Raja, V.; Shyam-Sundar, N.; Narayanan, K.R.; Senthil-Nathan, S. Toxic effect of essential oil and its compounds isolated from Sphaeranthus amaranthoides Burm. f. against dengue mosquito vector Aedes aegypti Linn. Pest. Biochem. Physiol. 2019, 160, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Chellappandian, M.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Karthi, S.; Thanigaivel, A.; Ponsankar, A.; Kalaivani, K.; Hunter, W.B. Botanical essential oils and uses as mosquitocides and repellents against dengue. Environ. Inter. 2018, 113, 214–230. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G. Plant-borne compounds and nanoparticles: Challenges for medicine, parasitology and entomology. Environ. Sci. Pollut. Res. 2018, 25, 10149–10150. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.H.; He, J.T.; Liu, Z.X.; Huang, J.G. Cytotoxicity of chemical constituents from Torricellia tiliifolia DC. on Spodoptera litura (SL-1) cells. Pest. Biochem. Physiol. 2018, 144, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Tewari, H.; Jyothi, K.N.; Kasana, V.K.; Prasad, A.R.; Prasuna, A.L. Insect attractant and oviposition enhancing activity of hexadecanoic acid ester derivatives for monitoring and trapping Caryedon serratus. J. Stored Prod. Res. 2015, 61, 32–38. [Google Scholar] [CrossRef]
- Harborne, J.B. Introduction to Ecological Biochemistry; Elsevier: Berkely, CA, USA, 2003; ISBN 0-12-324685-7. [Google Scholar]
- Edwin, E.; Vasantha-Srinivasan, P.; Senthil-Nathan, S.; Thanigaivel, A.; Ponsankar, A.; Pradeepa, V.; Selin-Rani, S.; Kalaivani, K.; Hunter, W.B.; Abdel-Megeed, A.; et al. Anti-dengue efficacy of bioactive andrographolide from Andrographis paniculata (Lamiales: Acanthaceae) against the primary dengue vector Aedes aegypti (Diptera: Culicidae). Acta Trop. 2016, 163, 167–178. [Google Scholar] [CrossRef]
- Shaalan, E.A.S.; Canyon, D.V.; Younes, M.W.F.; Abdel-Wahab, H.; Mansour, A. A review of botanical phytochemicals with mosquitocidal potential. Environ. Int. 2005, 31, 1149–1166. [Google Scholar] [CrossRef]
- Hugo, L.E.; Jeffery, J.A.L.; Trewin, B.J.; Wockner, L.F.; Yen, N.T.; Le, N.H.; Nghia, L.T.; Hine, E.; Ryan, P.A.; Kay, B.H. Adult survivorship of the dengue mosquito Aedes aegypti varies seasonally in Central Vietnam. PLoS Negl. Trop. Dis. 2014, 8, e2669. [Google Scholar] [CrossRef] [PubMed]
- Koodalingam, A.; Mullainadhan, P.; Arumugam, M. Effects of extract of soapnut Sapindus emarginatus on esterases and phosphatases of the vector mosquito, Aedes aegypti (Diptera: Culicidae). Acta Trop. 2011, 118, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Zibaee, A.; Alborzi, Z.; Hoda, H. A review on digestive TAG-lipases of insects. Arch. Phytopathol. Plant. Prot. 2014, 47, 2187–2192. [Google Scholar] [CrossRef]
- Ray, D.; Pautou, M.P.; Meyran, J.C. Histopathological effects of tannic acid on the midgut epithelium of some aquatic diptera larvae. J. Inver. Pathol. 1999, 73, 173–181. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Name | Formula | Weight (g/mol) | R/T (min) | PA (%) | Chemical Structure |
|---|---|---|---|---|---|
| Hexadecane | C16H34 | 226.4 | 9.033 | 0.75 |  |
| n-Eicosane | C20H42 | 282.5 | 13.23 | 1.13 |  |
| Tetratriacontane | C34H70 | 478.9 | 19.76 | 1.45 |  |
| Ethyl-Hexadecanoate | C18H36O2 | 284.5 | 21.46 | 1.32 |  |
| Neophytadiene | C20H38 | 278.5 | 21.97 | 0.83 |  |
| n-Hexatriacontane | C36H74 | 507 | 22.46 | 5.13 |  |
| Hexacosane | C26H54 | 366.7 | 24.67 | 6.13 |  |
| n-Octadecadienoic acid | C18H32O2 | 280.4 | 31.78 | 14.63 |  |
| n-Hexadecanoic acid | C16H32O2 | 256.4 | 33.45 | 6.73 |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amala, K.; Karthi, S.; Ganesan, R.; Radhakrishnan, N.; Srinivasan, K.; Mostafa, A.E.-Z.M.A.; Al-Ghamdi, A.A.; Alkahtani, J.; Elshikh, M.S.; Senthil-Nathan, S.; et al. Bioefficacy of Epaltes divaricata (L.) n-Hexane Extracts and Their Major Metabolites against the Lepidopteran Pests Spodoptera litura (fab.) and Dengue Mosquito Aedes aegypti (Linn.). Molecules 2021, 26, 3695. https://doi.org/10.3390/molecules26123695
Amala K, Karthi S, Ganesan R, Radhakrishnan N, Srinivasan K, Mostafa AE-ZMA, Al-Ghamdi AA, Alkahtani J, Elshikh MS, Senthil-Nathan S, et al. Bioefficacy of Epaltes divaricata (L.) n-Hexane Extracts and Their Major Metabolites against the Lepidopteran Pests Spodoptera litura (fab.) and Dengue Mosquito Aedes aegypti (Linn.). Molecules. 2021; 26(12):3695. https://doi.org/10.3390/molecules26123695
Chicago/Turabian StyleAmala, Kesavan, Sengodan Karthi, Raja Ganesan, Narayanaswamy Radhakrishnan, Kumaraswamy Srinivasan, Abd El-Zaher M. A. Mostafa, Abdullah Ahmed Al-Ghamdi, Jawaher Alkahtani, Mohamed Soliman Elshikh, Sengottayan Senthil-Nathan, and et al. 2021. "Bioefficacy of Epaltes divaricata (L.) n-Hexane Extracts and Their Major Metabolites against the Lepidopteran Pests Spodoptera litura (fab.) and Dengue Mosquito Aedes aegypti (Linn.)" Molecules 26, no. 12: 3695. https://doi.org/10.3390/molecules26123695
APA StyleAmala, K., Karthi, S., Ganesan, R., Radhakrishnan, N., Srinivasan, K., Mostafa, A. E. -Z. M. A., Al-Ghamdi, A. A., Alkahtani, J., Elshikh, M. S., Senthil-Nathan, S., Vasantha-Srinivasan, P., & Krutmuang, P. (2021). Bioefficacy of Epaltes divaricata (L.) n-Hexane Extracts and Their Major Metabolites against the Lepidopteran Pests Spodoptera litura (fab.) and Dengue Mosquito Aedes aegypti (Linn.). Molecules, 26(12), 3695. https://doi.org/10.3390/molecules26123695