
Malformin-A1 (MA1) Sensitizes Chemoresistant Ovarian Cancer Cells to Cisplatin-Induced Apoptosis

 ,
,  , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
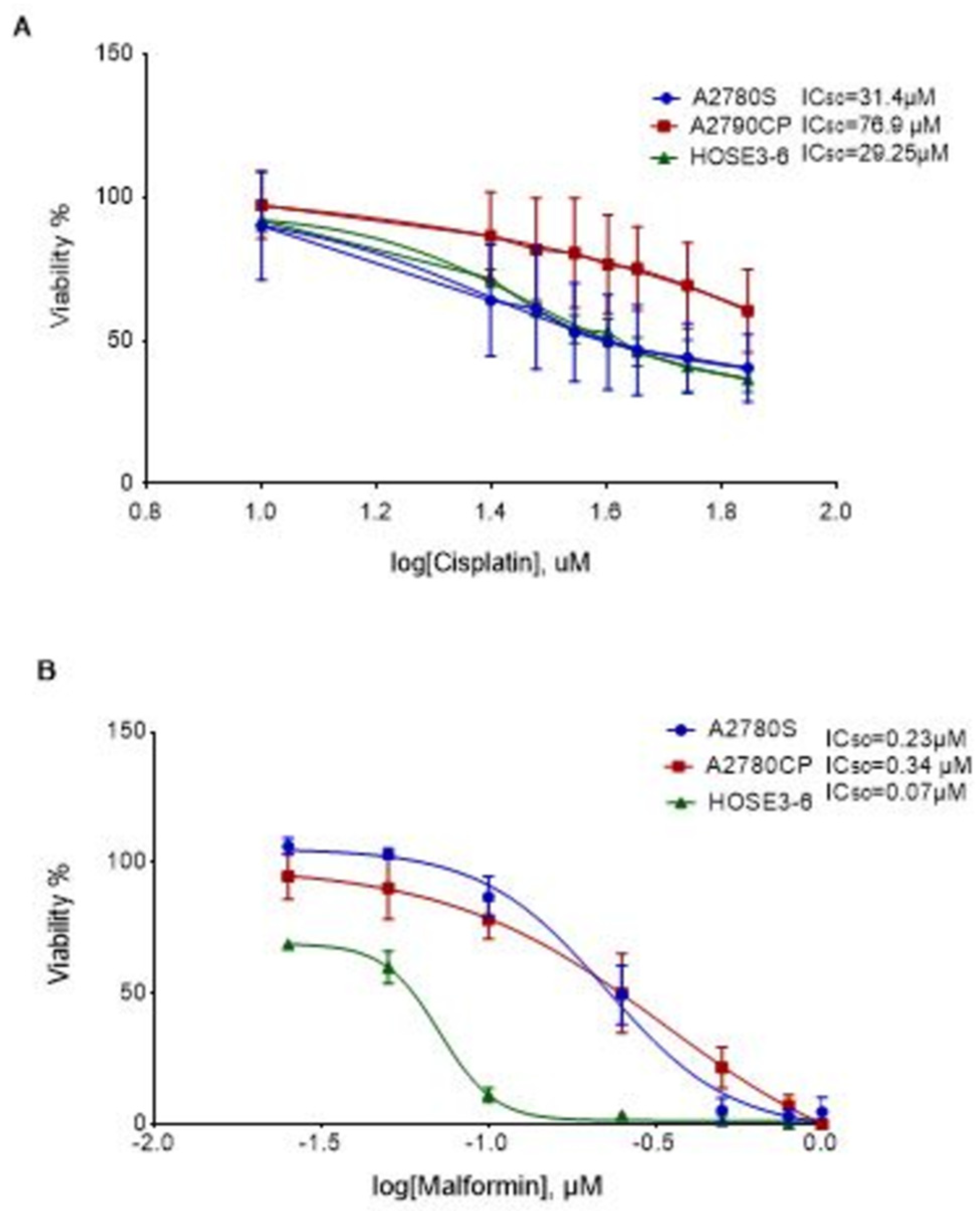
2.1. The Effect of Malformin A1 on Ovarian Cancer Cell Lines
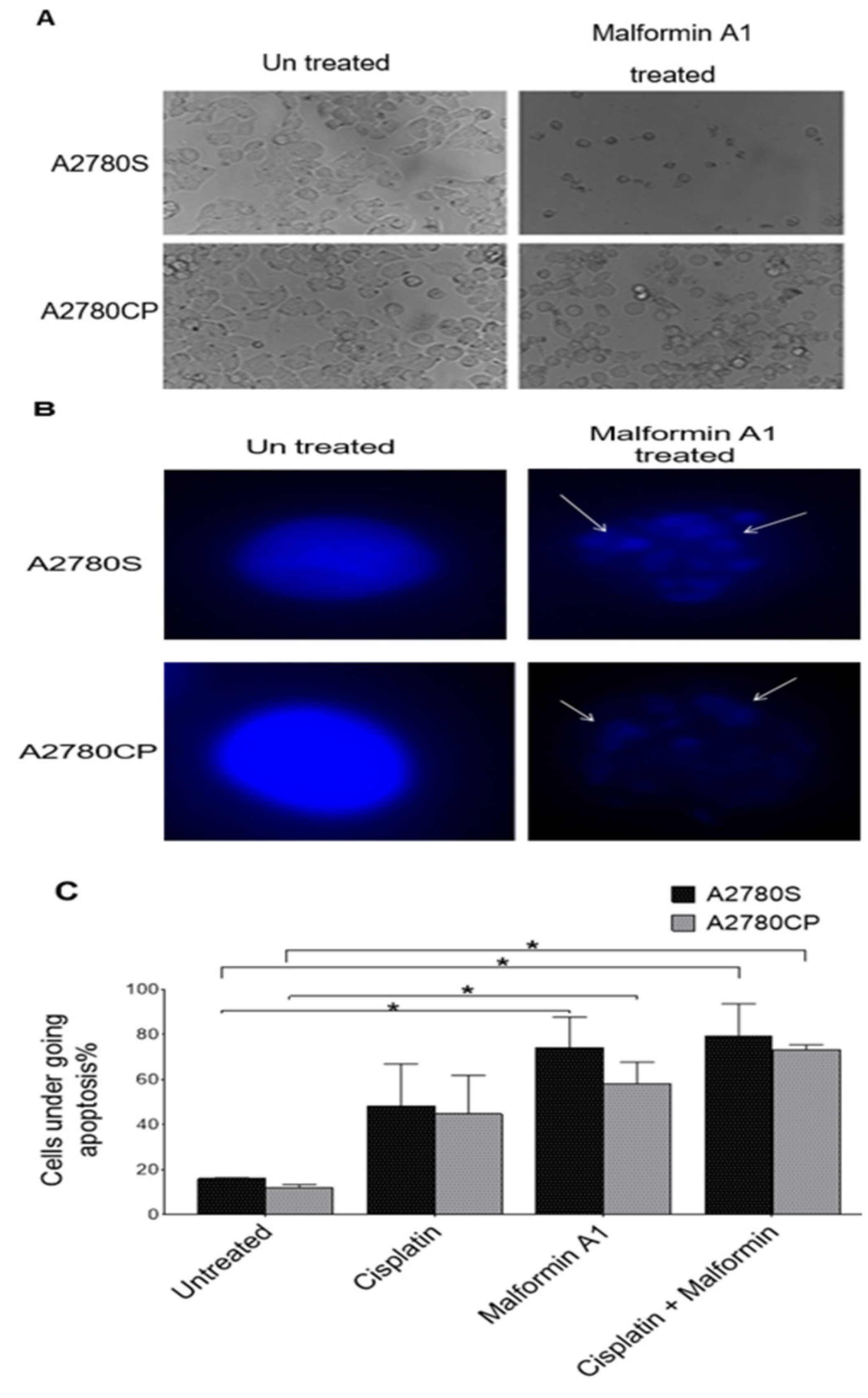
2.2. Morphological Hallmarks of Cell Death
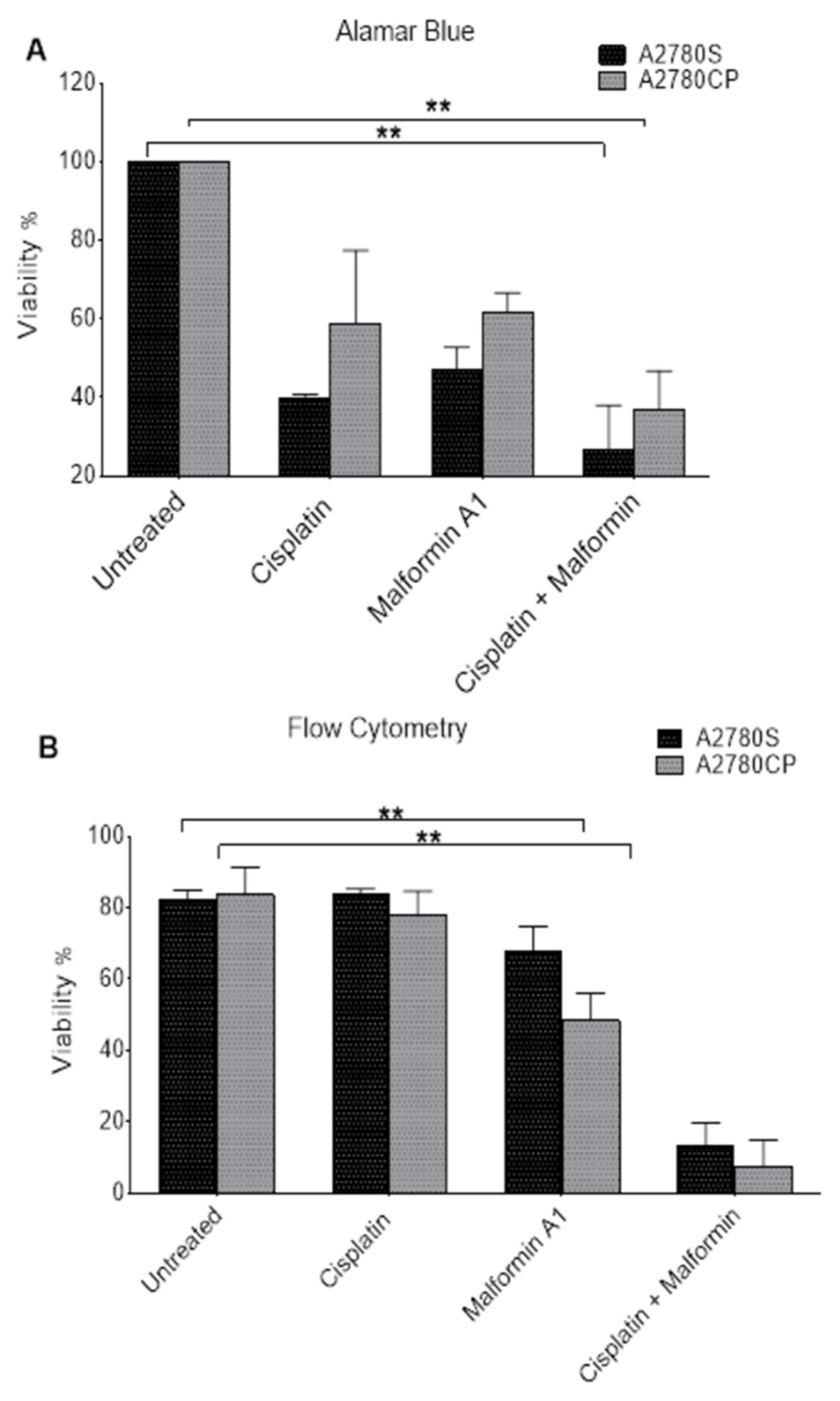
2.3. The Effect of Combined Malformin A1 to Cisplatin
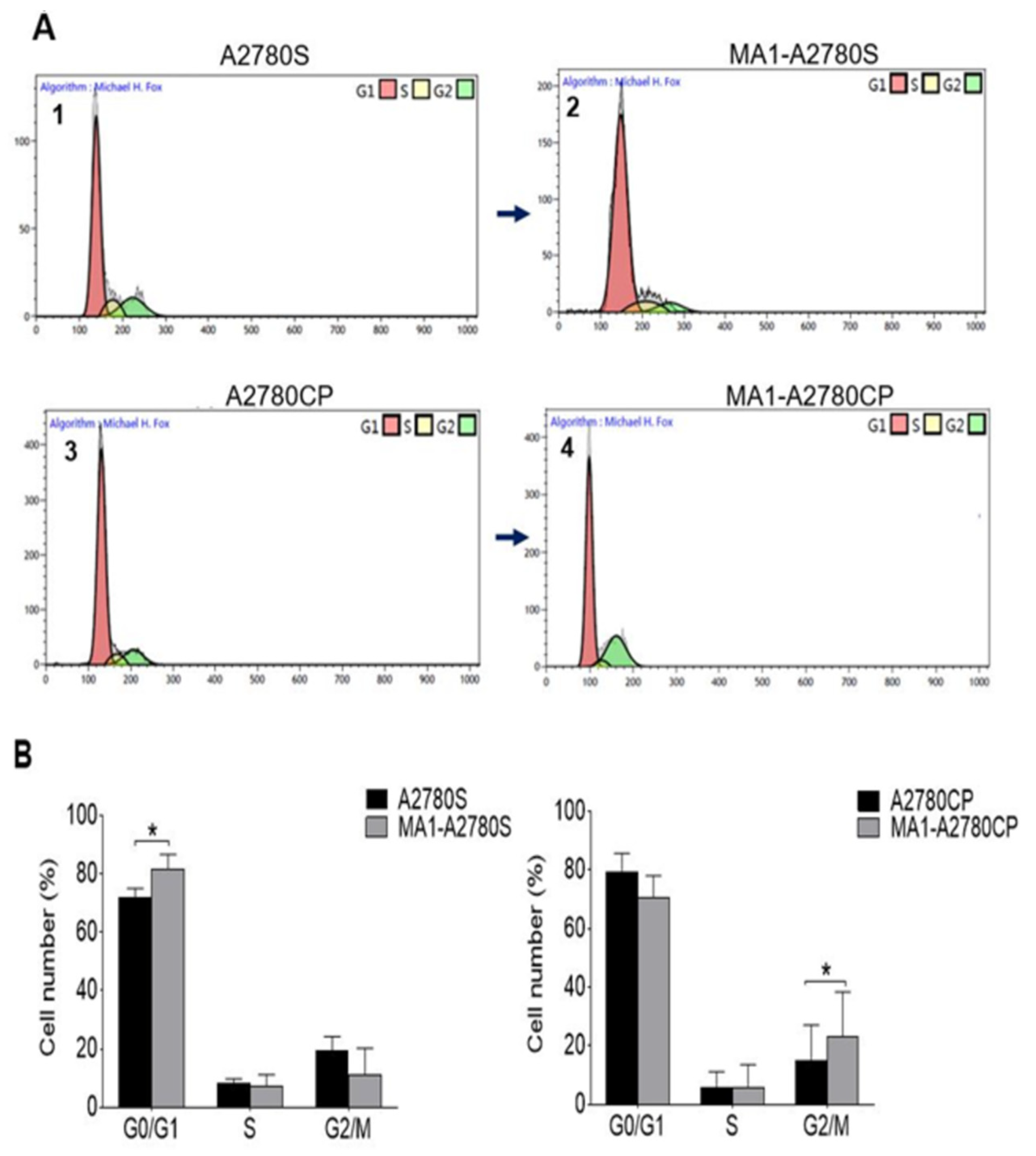
2.4. Cell Cycle
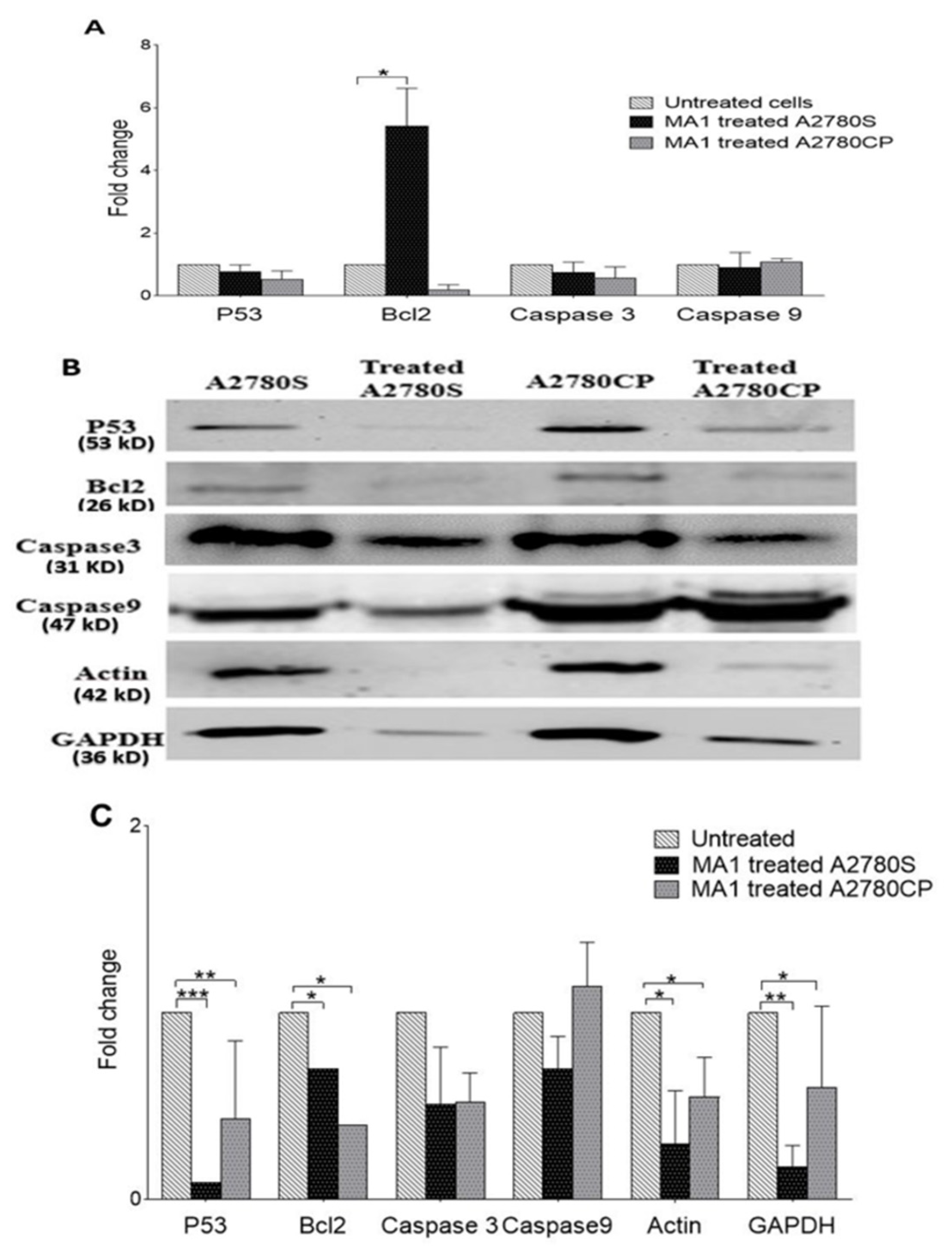
2.5. Expression of Apoptosis-Related Genes in Ovarian Cancer Cells
2.6. The Effect of MA1 Treatment on Proteins Expression
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Hoechst Dye Staining of Cells
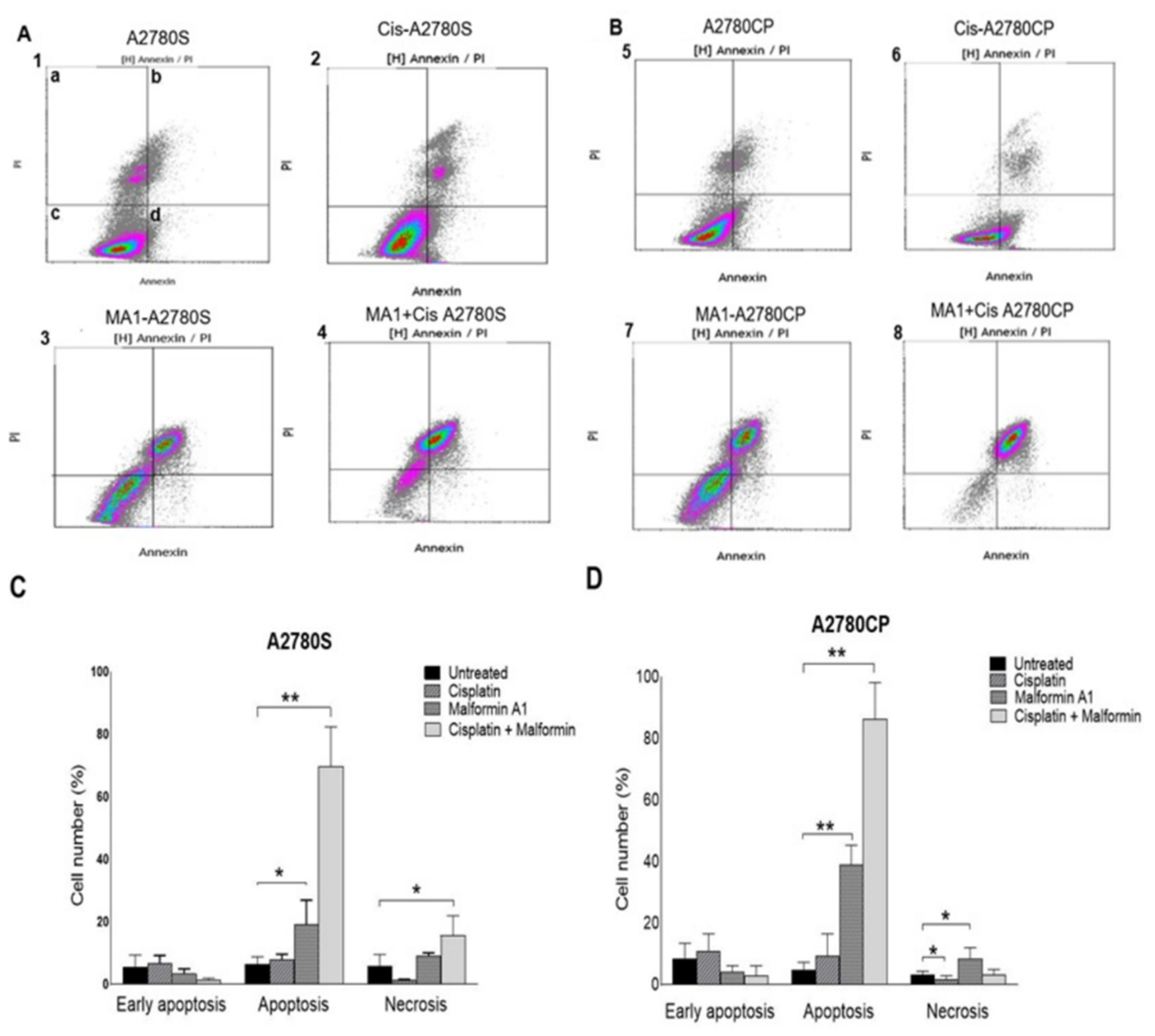
4.5. Annexin V-FITC Apoptosis and Cell Cycle Assays
4.6. Quantitative Real Time-PCR (qPCR)
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carcangiu, M.L.; Kurman, R.J.; Carcangiu, M.L.; Herrington, C.S. (Eds.) Tumours of the ovary. In WHO Classification of Tumours of Female Reproductive Organs, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2014; Volume 6, pp. 11–83. [Google Scholar]
- McGuire, W.; Markman, M. Primary ovarian cancer chemotherapy: Current standards of care. Br. J. Cancer 2003, 89, S3–S8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perren, T.J.; Swart, A.M.; Pfisterer, J.; Ledermann, J.A.; Pujade-Lauraine, E.; Kristensen, G.; Carey, M.S.; Beale, P.; Cervantes, A.; Kurzeder, C. A phase 3 trial of bevacizumab in ovarian cancer. N. Engl. J. Med. 2011, 365, 2484–2496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burger, R.A.; Brady, M.F.; Bookman, M.A.; Fleming, G.F.; Monk, B.J.; Huang, H.; Mannel, R.S.; Homesley, H.D.; Fowler, J.; Greer, B.E. Incorporation of bevacizumab in the primary treatment of ovarian cancer. N. Engl. J. Med. 2011, 365, 2473–2483. [Google Scholar] [CrossRef] [Green Version]
- Markman, M. Combination versus sequential cytotoxic chemotherapy in recurrent ovarian cancer: Time for an evidence-based comparison. Gynecol. Oncol. 2010, 118, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Konstantinopoulos, P.A.; Spentzos, D.; Karlan, B.Y.; Taniguchi, T.; Fountzilas, E.; Francoeur, N.; Levine, D.A.; Cannistra, S.A. Gene expression profile of BRCAness that correlates with responsiveness to chemotherapy and with outcome in patients with epithelial ovarian cancer. J. Clin. Oncol. 2010, 28, 3555. [Google Scholar] [CrossRef] [Green Version]
- Ledermann, J.; Harter, P.; Gourley, C.; Friedlander, M.; Vergote, I.; Rustin, G.; Scott, C.; Meier, W.; Shapira-Frommer, R.; Safra, T. Olaparib maintenance therapy in platinum-sensitive relapsed ovarian cancer. N. Engl. J. Med. 2012, 366, 1382–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peres, L.C.; Cushing-Haugen, K.L.; Köbel, M.; Harris, H.R.; Berchuck, A.; Rossing, M.A.; Schildkraut, J.M.; Doherty, J.A. Invasive epithelial ovarian cancer survival by histotype and disease stage. JNCI J. Natl. Cancer Inst. 2019, 111, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Chalamala, C. Potent anticancer compoundsfrom the ocean. Acad. J. 2013, 5, 455–460. [Google Scholar]
- Cragg, G.M.; Newman, D.J. Discovery and development of antineoplastic agents from natural sources. Cancer Investig. 1999, 19, 153–163. [Google Scholar] [CrossRef]
- Donoghue, M.; Lemery, S.J.; Yuan, W.; He, K.; Sridhara, R.; Shord, S.; Zhao, H.; Marathe, A.; Kotch, L.; Jee, J.; et al. Eribulin mesylate for the treatment of patients with refractory metastatic breast cancer: Use of a “physician’s choice” control arm in a randomized approval trial. Clin. Cancer Res. 2012, 18, 1496–1505. [Google Scholar] [CrossRef] [Green Version]
- Rinehart, K.L. Antitumor Compounds from Tunicates. Med. Res. Rev. 2000, 20, 1–27. [Google Scholar] [CrossRef]
- Swami, U.; Chaudhary, I.; Ghalib, M.H.; Goel, S. Eribulin—A review of preclinical and clinical studies. Crit. Rev. Oncol./Hematol. 2012, 81, 163–184. [Google Scholar] [CrossRef] [Green Version]
- Dobretsov, S.; Tamimi, Y.; Al-Kindi, M.A.; Burney, I. Screening for Anti-Cancer Compounds in Marine. Sultan Qaboos Univ. Med. J. 2016, 16, 168–174. [Google Scholar] [CrossRef]
- Curtis, R.W.; Stevenson, W.R.; Tuite, J. Malformin in Aspergillus niger-Infected Onion Bulbs (Allium cepa). Appl. Microbiol. 1974, 28, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.-M.; Liang, X.-A.; Zhang, H.-C.; Liu, R. Cytotoxic and antibiotic cyclic pentapeptide from an endophytic Aspergillus tamarii of Ficus carica. J. Agric. Food Chem. 2016, 64, 3789–3793. [Google Scholar] [CrossRef]
- Koizumi, Y.; Nagai, K.; Hasumi, K.; Kuba, K.; Sugiyama, T. Structure–activity relationship of cyclic pentapeptide malformins as fibrinolysis enhancers. Bioorganic Med. Chem. Lett. 2016, 26, 5267–5271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagimori, K.; Fukuda, T.; Hasegawa, Y.; Ōmura, S.; Tomoda, H. Fungal malformins inhibit bleomycin-induced G2 checkpoint in Jurkat cells. Biol. Pharm. Bull. 2007, 30, 1379–1383. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Oh, H.H.; Park, Y.L.; Yu, H.M.; Myung, D.S.; Cho, S.B.; Lee, W.S.; Park, D.; Joo, Y.E. Malformin A1 treatment alters invasive and oncogenic phenotypes of human colorectal cancer cells through stimulation of the p38 signaling pathway. Int. J. Oncol. 2017, 51, 959–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, M.; Wang, D.; Li, X.; Wang, W.; Lou, H.; Yuan, H. Malformin A1 promotes cell death through induction of apoptosis, necrosis and autophagy in prostate cancer cells. Cancer Chemother. Pharmacol. 2016, 77, 63–75. [Google Scholar] [CrossRef]
- Zhan, J.; Gunaherath, G.K.B.; Wijeratne, E.K.; Gunatilaka, A.L. Asperpyrone D and other metabolites of the plant-associated fungal strain Aspergillus tubingensis. Phytochemistry 2007, 68, 368–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notarte, K.; Nakao, Y.; Yaguchi, T.; Bungihan, M.; Suganuma, K.; dela Cruz, T.E. Trypanocidal activity, cytotoxicity and histone modifications induced by malformin A (1) isolated from the marine-derived fungus Aspergillus tubingensis IFM 63452. Mycosphere 2017, 8, 111–120. [Google Scholar] [CrossRef]
- Cornelison, R.; Llaneza, D.C.; Landen, C.N. Emerging therapeutics to overcome chemoresistance in epithelial ovarian cancer: A mini-review. Int. J. Mol. Sci. 2017, 18, 2171. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, K.; Foletta, V.; Bernard, O.; Itoh, K. A role for LIM kinase in cancer invasion. Proc. Natl. Acad. Sci. USA 2003, 100, 7247–7252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prudent, R.; Vassal-Stermann, E.; Nguyen, C.H.; Pillet, C.; Martinez, A.; Prunier, C.; Barette, C.; Soleilhac, E.; Filhol, O.; Beghin, A.; et al. Pharmacological inhibition of LIM kinase stabilizes microtubules and inhibits neoplastic growth. Cancer Res. 2012, 72, 4429–4439. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Doherty, J.; Antonipillai, J.; Chen, S.; Devlin, M.; Visser, K.; Baell, J.; Street, I.; Anderson, R.L.; Bernard, O. LIM kinase inhibition reduces breast cancer growth and invasiveness but systemic inhibition does not reduce metastasis in mice. Clin. Exp. Metastasis 2013, 30, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Colell, A.; Green, D.R.; Ricci, J.E. Novel roles for GAPDH in cell death and carcinogenesis. Cell Death Differ. 2009, 16, 1573–1581. [Google Scholar] [CrossRef] [Green Version]
- Revillion, F.; Pawlowski, V.; Hornez, L.; Peyrat, J.P. Glyceraldehyde-3-phosphate dehydrogenase gene expression inhuman breast cancer. Eur. J. Cancer 2000, 36, 1038–1042. [Google Scholar] [CrossRef]
- Huang, Q.; Lan, F.; Zheng, Z.; Xie, F.; Han, J.; Dong, L.; Xie, Y.; Zheng, F. Akt2 kinase suppresses glyceraldehyde-3-phosphate dehydrogenase (GAPDH)-mediated apoptosis in ovarian cancer cells via phosphorylating GAPDH at threonine 237 and decreasing its nuclear translocation. J. Biol. Chem. 2011, 286, 42211–42220. [Google Scholar] [CrossRef] [Green Version]
- Jung, D.W.; Kim, W.H.; Seo, S.; Oh, E.; Yim, S.H.; Ha, H.H.; Chang, Y.T.; Williams, D.R. Chemical targeting of GAPDH moonlighting function in cancer cells reveals its role in tubulin regulation. Chem. Biol. 2014, 21, 1533–1545. [Google Scholar] [CrossRef] [Green Version]
- Chleq-Deschamps, C.M.; LeBrun, D.P.; Huie, P.; Besnier, D.P.; Warnke, R.A.; Sibley, R.K.; Cleary, M.L. Topographical dissociation of BCL-2 messenger RNA and protein expression in human lymphoid tissues. Blood. 1993, 81, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Marks, J.R.; Davidoff, A.M.; Kerns, B.J.; Humphrey, P.A.; Pence, J.C.; Dodge, R.K.; Clarke-Pearson, D.L.; Iglehart, J.D.; Bast, R.C.; Berchuck, A. Overexpression and mutation of p53 in epithelial ovarian cancer. Cancer Res. 1991, 51, 2979–2984. [Google Scholar] [PubMed]
- Oren, M.; Rotter, V. Mutant p53 gain-of-function in cancer. Cold Spring Harb. Perspect. Biol. 2010, 2, a001107. [Google Scholar] [CrossRef]
- Tsujimoto, Y.; Cossman, J.; Jaffe, E. CM Croce, Involvement of the bcl-2 gene in human follicular lymphoma. Science 1985, 228, 1440–1443. [Google Scholar] [CrossRef]
- Tsujimoto, Y. Stress-resistance conferred by high level of bcl-2 alpha protein in human B lymphoblastoid cell. Europe MPC 1989, 11, 1331–1336. [Google Scholar]
- Huang, Q.; Li, F.; Liu, X.; Li, W.; Shi, W.; Liu, F.-F.; O’sullivan, B.; He, Z.; Peng, Y.; Tan, A.-C. Caspase 3–mediated stimulation of tumor cell repopulation during cancer radiotherapy. Nat. Med. 2011, 17, 860–866. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.; Leibowitz, B.; Chen, D.; He, K.; Zou, F.; Sobol, R.; Beer-Stolz, D.; Zhang, L.; Yu, J. Loss of Caspase-3 sensitizes colon cancer cells to genotoxic stress via RIP1-dependent necrosis. Cell Death Dis. 2015, 6, e1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdullah, N.; Tamimi, Y.; Dobretsov, S.; Balushi, N.A.; Alshekaili, J.; Al Balushi, H.; Al Kindi, M.; Hassan, S.I.; Bahlani, S.A.; Tsang, B.K.; et al. Malformin-A1 (MA1) Sensitizes Chemoresistant Ovarian Cancer Cells to Cisplatin-Induced Apoptosis. Molecules 2021, 26, 3624. https://doi.org/10.3390/molecules26123624
Abdullah N, Tamimi Y, Dobretsov S, Balushi NA, Alshekaili J, Al Balushi H, Al Kindi M, Hassan SI, Bahlani SA, Tsang BK, et al. Malformin-A1 (MA1) Sensitizes Chemoresistant Ovarian Cancer Cells to Cisplatin-Induced Apoptosis. Molecules. 2021; 26(12):3624. https://doi.org/10.3390/molecules26123624
Chicago/Turabian StyleAbdullah, Nada, Yahya Tamimi, Sergey Dobretsov, Najwa Al Balushi, Jalila Alshekaili, Hamed Al Balushi, Mahmood Al Kindi, Syed Imran Hassan, Shadia Al Bahlani, Benjamin K. Tsang, and et al. 2021. "Malformin-A1 (MA1) Sensitizes Chemoresistant Ovarian Cancer Cells to Cisplatin-Induced Apoptosis" Molecules 26, no. 12: 3624. https://doi.org/10.3390/molecules26123624
APA StyleAbdullah, N., Tamimi, Y., Dobretsov, S., Balushi, N. A., Alshekaili, J., Al Balushi, H., Al Kindi, M., Hassan, S. I., Bahlani, S. A., Tsang, B. K., & Burney, I. A. (2021). Malformin-A1 (MA1) Sensitizes Chemoresistant Ovarian Cancer Cells to Cisplatin-Induced Apoptosis. Molecules, 26(12), 3624. https://doi.org/10.3390/molecules26123624