
A Standardized Extract Prepared from Red Orange and Lemon Wastes Blocks High-Fat Diet-Induced Hyperglycemia and Hyperlipidemia in Mice

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of RLE
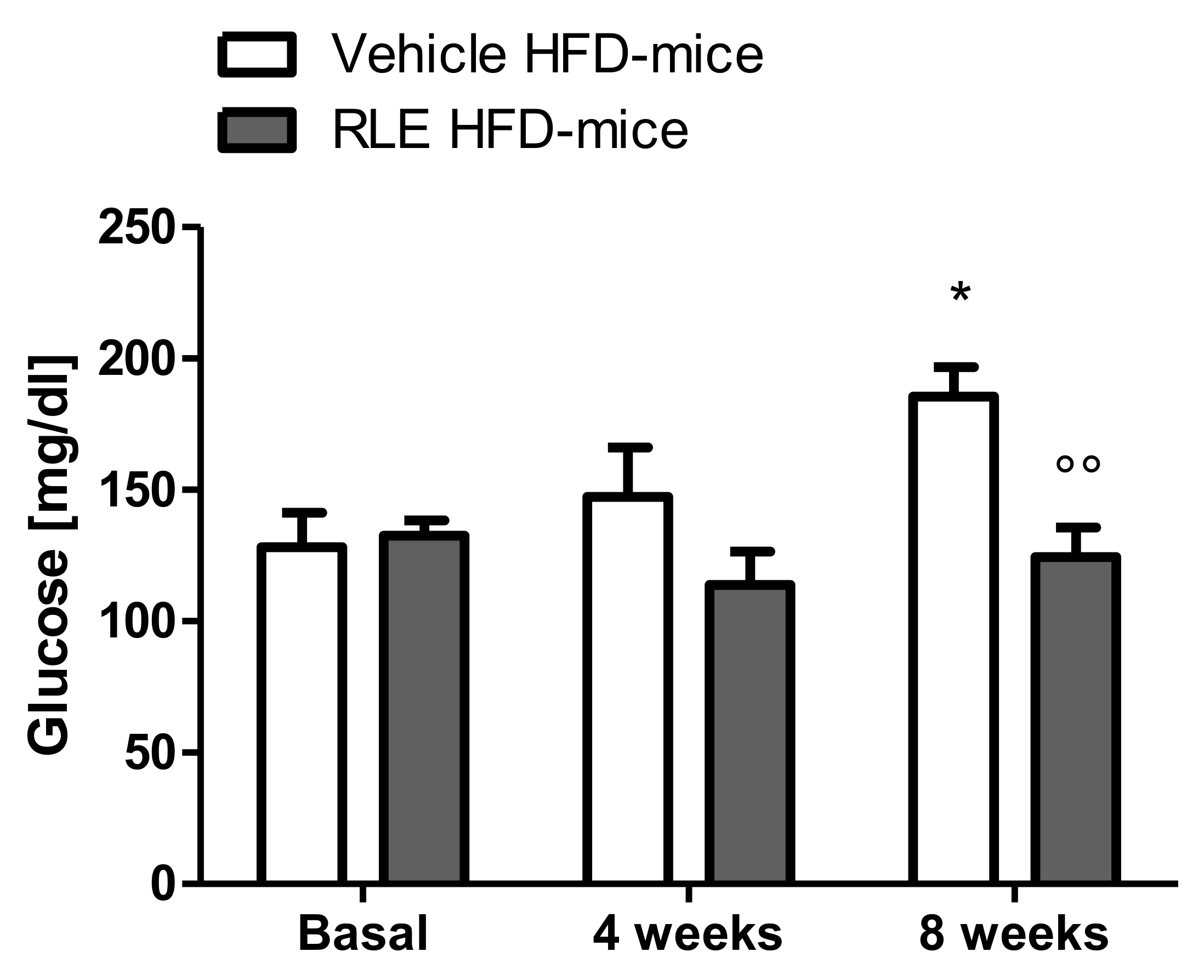
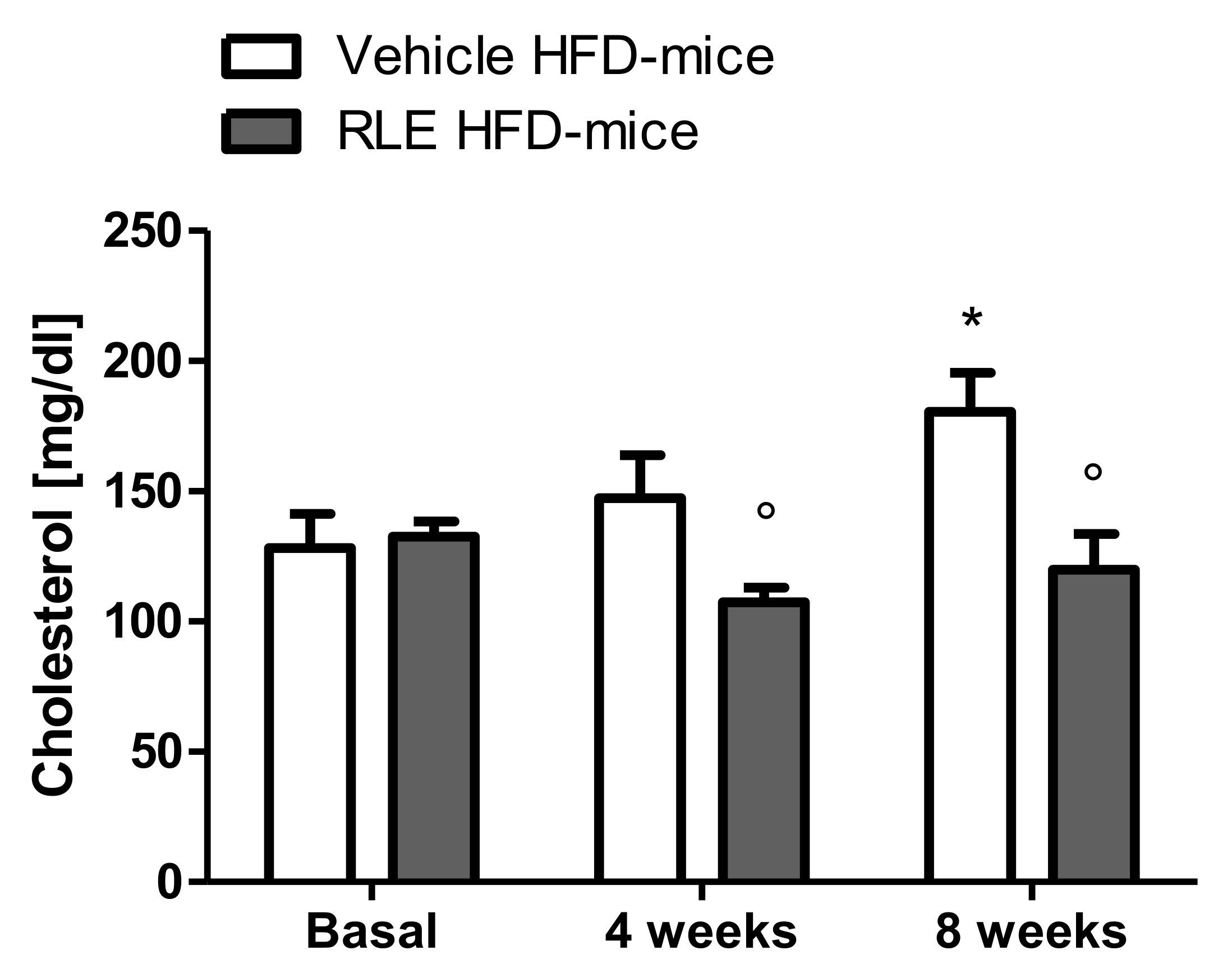
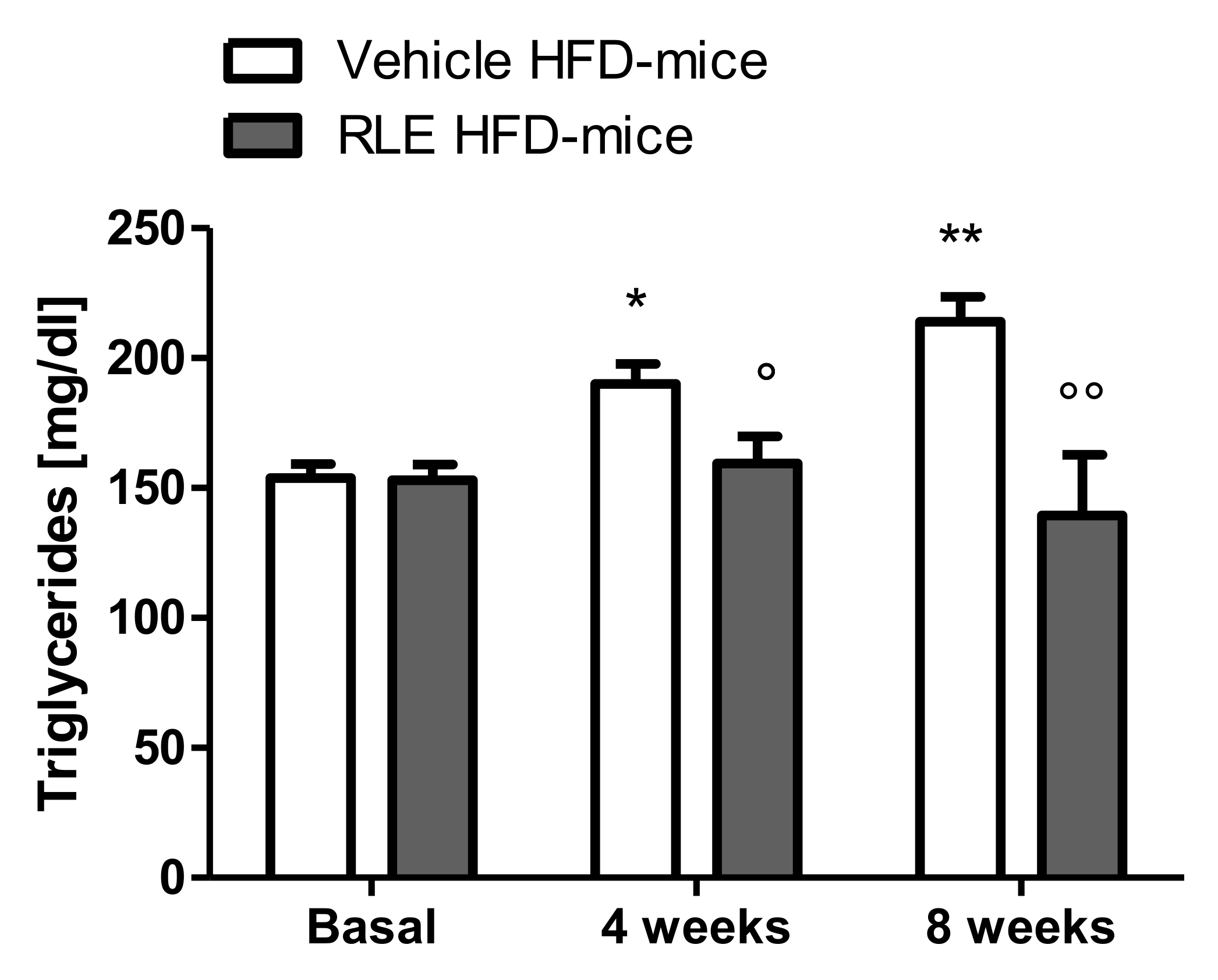
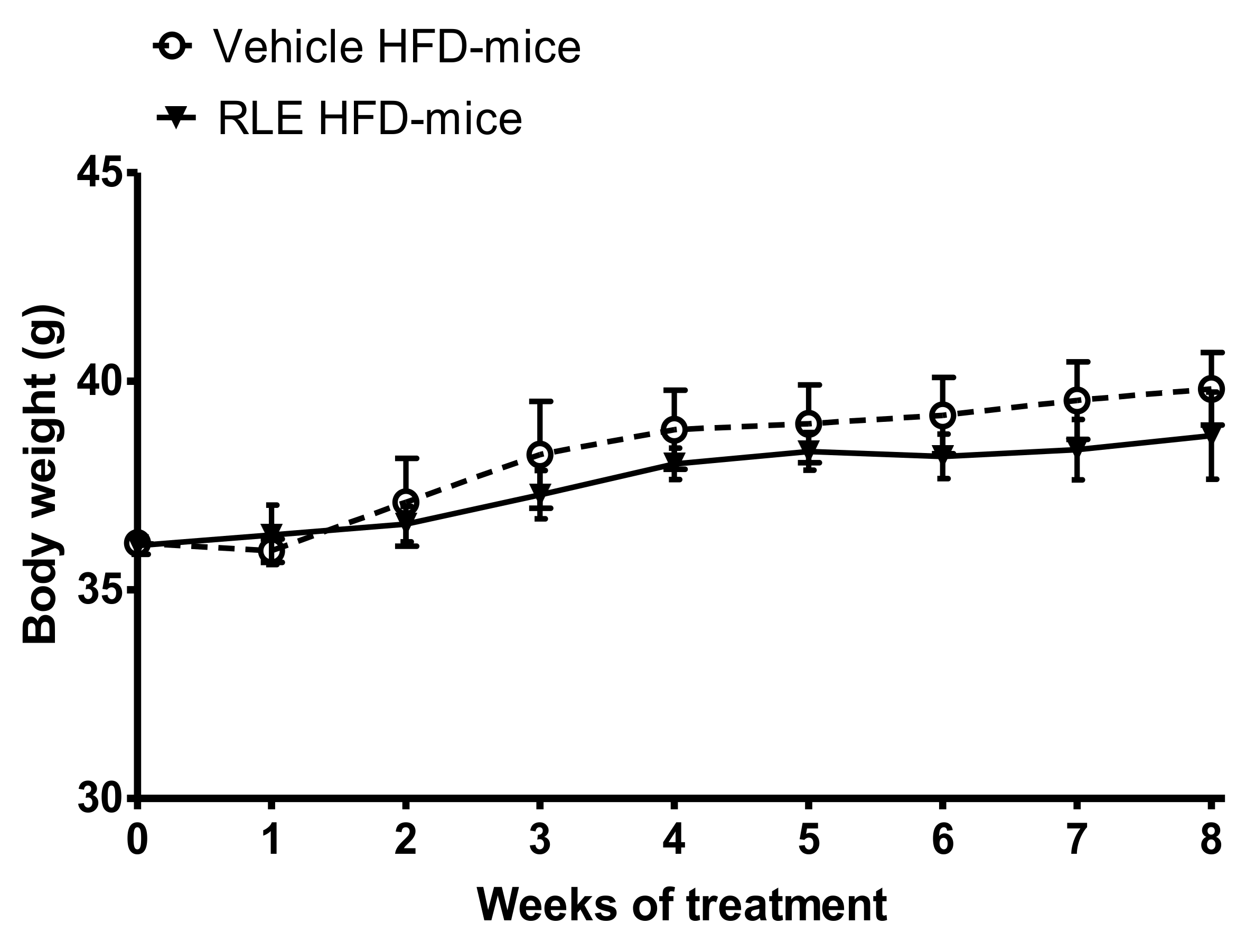
2.2. In Vivo Studies
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Chemical Composition of RLE
4.3. In Vivo Studies
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Food and Agriculture Organization of the United Nations. Database. Available online: http://www.fao.org/faostat/en/ (accessed on 1 July 2021).
- Rapisarda, P.; Tomaino, A.; Cascio, R.L.; Bonina, F.; de Pasquale, A.; Saija, A. Antioxidant Effectiveness As Influenced by Phenolic Content of Fresh Orange Juices. J. Agric. Food Chem. 1999, 47, 4718–4723. [Google Scholar] [CrossRef] [PubMed]
- Castañeda-Ovando, A.; Pacheco-Hernández, M.D.; Páez-Hernández, M.E.; Rodríguez, J.A.; Andrés, C. Galán-Vidal Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Titta, L.; Trinei, M.; Stendardo, M.; Berniakovich, I.; Petroni, K.; Tonelli, C.; Riso, P.; Porrini, M.; Minucci, S.; Pelicci, P.G.; et al. Blood orange juice inhibits fat accumulation in mice. Int. J. Obes. 2010, 34, 578–588. [Google Scholar] [CrossRef] [Green Version]
- Rapisarda, P.; Bellomo, S.; Intrigliolo, F. Recent research developments in agricultural & food chemistry. Res. Signpost 2000, 4, 217–230. [Google Scholar]
- Saija, A.; Tomaino, A.; Cascio, R.L.; Rapisarda, P.; Dederen, J.C. In vitro antioxidant activity and in vivo photoprotective effect of a red orange extract. Int. J. Cosmet. Sci. 1998, 20, 331–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emim, J.A.D.S.; Oliveira, A.B.; Lapa, A.J. Pharmacological Evaluation of the Anti-inflammatory Activity of a Citrus Bioflavonoid, Hesperidin, and the Isoflavonoids, Duartin and Claussequinone, in Rats and Mice. J. Pharm. Pharmacol. 1994, 46, 118–122. [Google Scholar] [CrossRef]
- González-Molina, E.; Domínguez-Perles, R.; Moreno, D.A.; García-Viguera, C. Natural bioactive compounds of Citrus limon for food and health. J. Pharm. Biomed. Anal. 2010, 51, 327–345. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Yamamotoi, K.; Osawa, T. Isolation of eriocitrin (eriodictyol 7-rutinoside) from lemon fruit (Citrus limon BURM. f.) and its antioxidative activity. Food Sci. Technol. Int. Tokyo 1997, 3, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Miyake, Y.; Yamamoto, K.; Morimitsu, Y.; Osawa, T. Isolation of C-Glucosylflavone from Lemon Peel and Antioxidative Activity of Flavonoid Compounds in Lemon Fruit. J. Agric. Food Chem. 1997, 45, 4619–4623. [Google Scholar] [CrossRef]
- Del Río, J.A.; Fuster, M.D.; Gómez, P.; Porras, I.; García-Lidón, A.; Ortuño, A. Citrus limon: A source of flavonoids of pharmaceutical interest. Food Chem. 2004, 84, 457–461. [Google Scholar] [CrossRef]
- Amenta, M.; Ballistreri, G.; Fabroni, S.; Romeo, F.V.; Spina, A.; Rapisarda, P. Qualitative and nutraceutical aspects of lemon fruits grown on the mountainsides of the Mount Etna: A first step for a protected designation of origin or protected geographical indication application of the brand name ‘Limone dell’Etna’. Food Res. Int. 2015, 74, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, H.; Liu, Y.; Yang, L. High fat diet induced obesity model using four strainsof mice: Kunming, C57BL/6, BALB/c and ICR. Exp. Anim. 2020, 69, 326–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, M.; Fabroni, S.; Emma, R.; Ballistreri, G.; Amenta, M.; Currenti, W.; Rinzivillo, C.; Rapisarda, P. A new standardized phytoextract from red orange and lemon wastes (red orange and lemon extract) reduces basophil degranulation and activation. Nat. Prod. Res. 2020. [CrossRef]
- Damiano, S.; Lauritano, C.; Longobardi, C.; Andretta, E.; Elagoz, A.M.; Rapisarda, P.; Di Iorio, M.; Florio, S.; Ciarcia, R. Effects of a Red Orange and Lemon Extract in Obese Diabetic Zucker Rats: Role of Nicotinamide Adenine Dinucleotide Phosphate Oxidase. J. Clin. Med. 2020, 9, 1600. [Google Scholar] [CrossRef]
- Damiano, S.; Lombari, P.; Salvi, E.; Papale, M.; Giordano, A.; Amenta, M.; Ballistreri, G.; Fabroni, S.; Rapisarda, P.; Capasso, G.; et al. A red orange and lemon by-products extract rich in anthocyanins inhibits the progression of diabetic nephropathy. J. Cell. Physiol. 2019, 234, 23268–23278. [Google Scholar] [CrossRef]
- Damiano, S.; Iovane, V.; Squillacioti, C.; Mirabella, N.; Prisco, F.; Ariano, A.; Amenta, M.; Giordano, A.; Florio, S.; Ciarcia, R. Red orange and lemon extract prevents the renal toxicity induced by ochratoxin A in rats. J. Cell. Physiol. 2020, 235, 5386–5393. [Google Scholar] [CrossRef]
- Salzano, A.; Damiano, S.; D’Angelo, L.; Ballistreri, G.; Claps, S.; Rufrano, D.; Maggiolino, A.; Neglia, G.; De Palo, P.; Ciarcia, R. Productive Performance and Meat Characteristics of Kids Fed a Red Orange and Lemon Extract. Animals 2021, 11, 809. [Google Scholar] [CrossRef]
- De Felice, E.; Giaquinto, D.; Damiano, S.; Salzano, A.; Fabroni, S.; Ciarcia, R.; Scocco, P.; de Girolamo, P.; D’angelo, L. Distinct pattern of npy in gastro–entero–pancreatic system of goat kids fed with a new standardized red orange and lemon extract (Rle). Animals 2021, 11, 449. [Google Scholar] [CrossRef]
- O’Keefe, J.H.; Bell, D.S. Postprandial hyperglycemia/hyperlipidemia (postprandial dysmetabolism) is a cardiovascular risk factor. Am. J. Cardiol. 2007, 100, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Nam, Y.; Chung, Y.H.; Kim, H.R.; Park, E.S.; Chung, S.J.; Kim, J.H.; Sohn, U.D.; Kim, H.C.; Oh, K.W.; et al. Beneficial effects of phosphatidylcholine on high-fat diet-induced obesity, hyperlipidemia and fatty liver in mice. Life Sci. 2014, 118, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Poirier, H.; Degrace, P.; Niot, I.; Bernard, A.; Besnard, P. Localization and regulation of the putative membrane fatty-acid transporter (FAT) in the small intestine. Comparison with fatty acid-binding proteins (FABP). Eur. J. Biochem. 1996, 238, 368–373. [Google Scholar] [CrossRef]
- Chen, M.; Yang, Y.; Braunstein, E.; Georgeson, K.E.; Harmon, C.M. Gut expression and regulation of FAT/CD36: Possible role in fatty acid transport in rat enterocytes. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E916–E923. [Google Scholar] [CrossRef] [Green Version]
- Vega, M.A.; Bragado, R. Localization of the lipid receptors CD36 and CLA-1/SR-BI in the human gastrointestinal tract: Towards the identification of receptors mediating the intestinal absorption of dietary lipids. J. Histochem. Cytochem. 2001, 49, 1253–1260. [Google Scholar]
- Goudriaan, J.R.; den Boer, M.A.; Rensen, P.C.; Febbraio, M.; Kuipers, F.; Romijn, J.A.; Havekes, L.M.; Voshol, P.J. CD36 deficiency in mice impairs lipoprotein lipase-mediated triglyceride clearance. J. Lipid Res. 2005, 46, 2175–2181. [Google Scholar] [CrossRef] [Green Version]
- Werder, M.; Han, C.H.; Wehrli, E.; Bimmler, D.; Schulthess, G.; Hauser, H. Role of scavenger receptors SR-BI and CD36 in selective sterol uptake in the small intestine. Biochemistry 2001, 40, 11643–11650. [Google Scholar] [CrossRef] [PubMed]
- Röder, P.V.; Geillinger, K.E.; Zietek, T.S.; Thorens, B.; Koepsell, H.; Daniel, H. The role of SGLT1 and GLUT2 in intestinal glucose transport and sensing. PLoS ONE 2014, 9, e89977. [Google Scholar] [CrossRef] [PubMed]
- Kao, E.S.; Tseng, T.H.; Lee, H.J.; Chan, K.C.; Wang, C.J. Anthocyanin extracted from Hibiscus attenuate oxidized LDL-mediated foam cell formation involving regulation of CD36 gene. Chem. Biol. Interact. 2009, 179, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Solverson, P. Anthocyanin Bioactivity in Obesity and Diabetes: The Essential Role of Glucose Transporters in the Gut and Periphery. Cells 2020, 9, 2515. [Google Scholar] [CrossRef]
- Rapisarda, P.; Lo Bianco, M.; Pannuzzo, P.; Timpanaro, N. Effect of cold storage on vitamin C, phenolics and antioxidant activity of five orange genotypes [Citrus sinensis (L.) Osbeck]. Postharvest Biol. Technol. 2008, 49, 348–354. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound No. | [M]+ (m/z) | MSn (m/z) | Anthocyanin | Relative Composition a (%) |
|---|---|---|---|---|
| 1 | 611 | 449/287 | cyanidin 3,5-diglucoside | 1.19 |
| 2 | 465 | 303 | delphinidin 3-glucoside | 2.69 |
| 3 | 611 | 287 | cyanidin 3-sophoroside | 0.39 |
| 4 | 449 | 287 | cyanidin 3-glucoside | 39.94 |
| 5 | 595 | 287 | cyanidin 3-rutinoside | 1.32 |
| 6 | 479 | 317 | petunidin 3-glucoside | 1.47 |
| 7 | 551 | 465/303 | delphinidin 3-(6″-malonyl)glucoside | 1.41 |
| 8 | 463 | 301 | peonidin 3-glucoside | 2.93 |
| 9 | 565 | 479/317 | petunidin 3-(6″-malonyl)glucoside | 1.48 |
| 10 | 535 | 449/287 | cyanidin 3-(6″-malonyl)glucoside | 21.86 |
| 11 | 593 | 449/287 | cyanidin 3-(6″-dioxalyl)glucoside | 5.70 |
| 12 | - | 271 | pelargonidin derivative | 1.34 |
| 13 | 549 | 463/301 | peonidin 3-(6″-malonyl)glucoside | 13.85 |
| 14 | - | 287 | cyanidin derivative | 2.29 |
| 15 | - | 301 | peonidin derivative | 1.85 |
| Total anthocyanins (g Cya-3-glu equivalents/100 g) b | 2.56 ± 0.08 | |||
| Compound No. | [M‒H]‒ (m/z) | MSn (m/z) | Flavanone | g/100 g a |
|---|---|---|---|---|
| 1 | 595 | 287 | eriocitrin | 11.02 ± 0.02 |
| 2 | 579 | 271 | narirutin | 1.23 ± 0.01 |
| 3 | 609 | 301 | hesperidin | 3.46 ± 0.01 |
| 4 | 593 | 285 | didymin | 0.15 ± 0.02 |
| Total flavanones (g Hesperidin equivalents/100 g) | 15.86 ± 0.01 | |||
| Compound No. | [M‒H]‒ (m/z) | MSn (m/z) | Hydroxycinnamic Acids | g/100 g a |
|---|---|---|---|---|
| 1 | 325 | 163/145 | p-coumaric | 0.366 ± 4.13 |
| 2 | 355 | 193/175 | ferulic | 0.970.± 3.72 |
| 3 | 385 | 223/205 | sinapic | 0.432 ± 1.45 |
| Total hydroxycinnamic acids (g/100 g) | 1.77 ± 0.009 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiechio, S.; Zammataro, M.; Barresi, M.; Amenta, M.; Ballistreri, G.; Fabroni, S.; Rapisarda, P. A Standardized Extract Prepared from Red Orange and Lemon Wastes Blocks High-Fat Diet-Induced Hyperglycemia and Hyperlipidemia in Mice. Molecules 2021, 26, 4291. https://doi.org/10.3390/molecules26144291
Chiechio S, Zammataro M, Barresi M, Amenta M, Ballistreri G, Fabroni S, Rapisarda P. A Standardized Extract Prepared from Red Orange and Lemon Wastes Blocks High-Fat Diet-Induced Hyperglycemia and Hyperlipidemia in Mice. Molecules. 2021; 26(14):4291. https://doi.org/10.3390/molecules26144291
Chicago/Turabian StyleChiechio, Santina, Magda Zammataro, Massimo Barresi, Margherita Amenta, Gabriele Ballistreri, Simona Fabroni, and Paolo Rapisarda. 2021. "A Standardized Extract Prepared from Red Orange and Lemon Wastes Blocks High-Fat Diet-Induced Hyperglycemia and Hyperlipidemia in Mice" Molecules 26, no. 14: 4291. https://doi.org/10.3390/molecules26144291
APA StyleChiechio, S., Zammataro, M., Barresi, M., Amenta, M., Ballistreri, G., Fabroni, S., & Rapisarda, P. (2021). A Standardized Extract Prepared from Red Orange and Lemon Wastes Blocks High-Fat Diet-Induced Hyperglycemia and Hyperlipidemia in Mice. Molecules, 26(14), 4291. https://doi.org/10.3390/molecules26144291