
Antiviral Potential of Plants against Noroviruses

 , , and
, , and
Abstract
:1. Introduction
2. Characteristics of Human Norovirus
3. Methodology of Research Regarding the Antiviral Activity of Phytochemicals
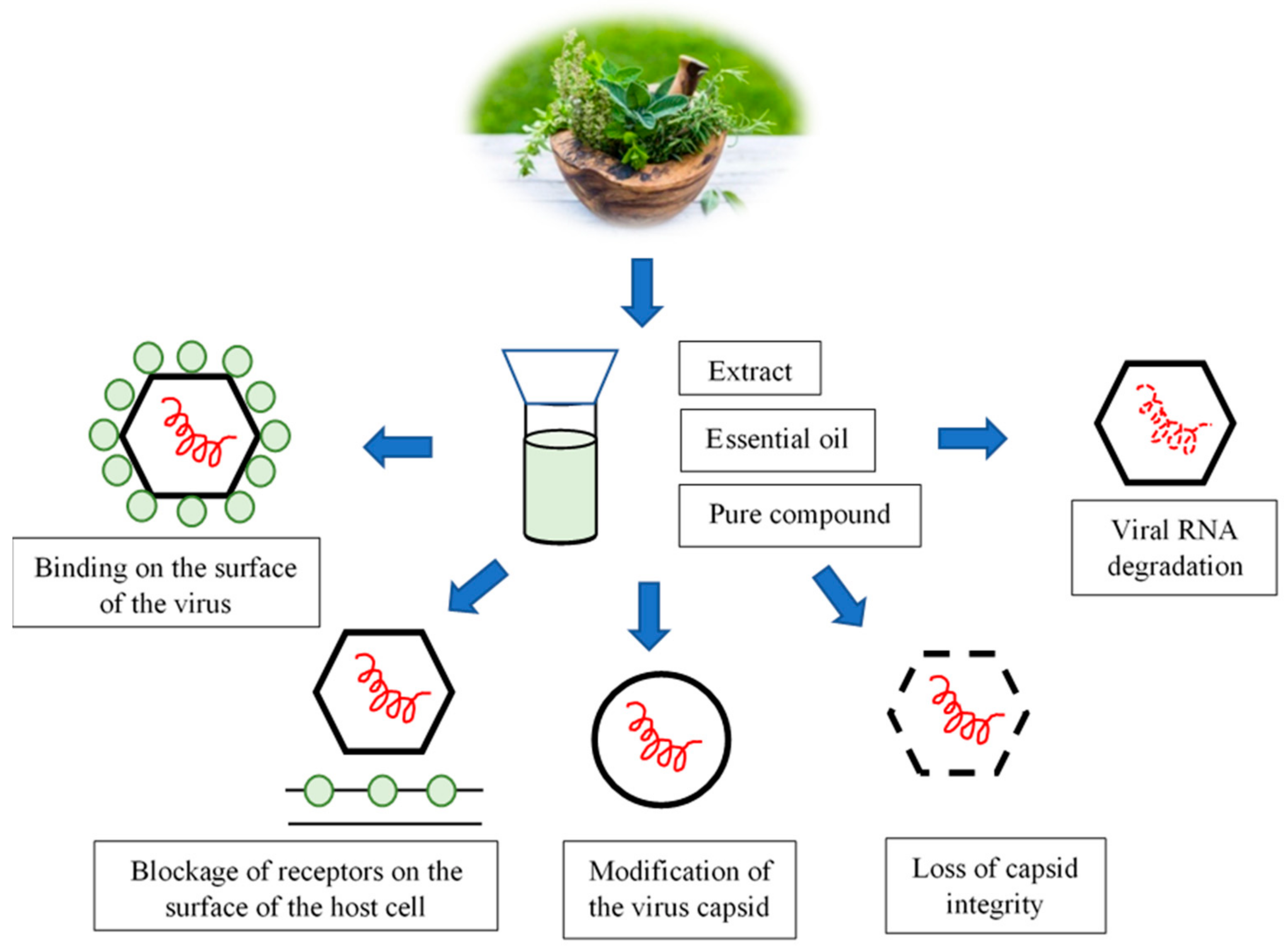
4. Mechanism of the Antiviral Action of Compounds of Plant Origin
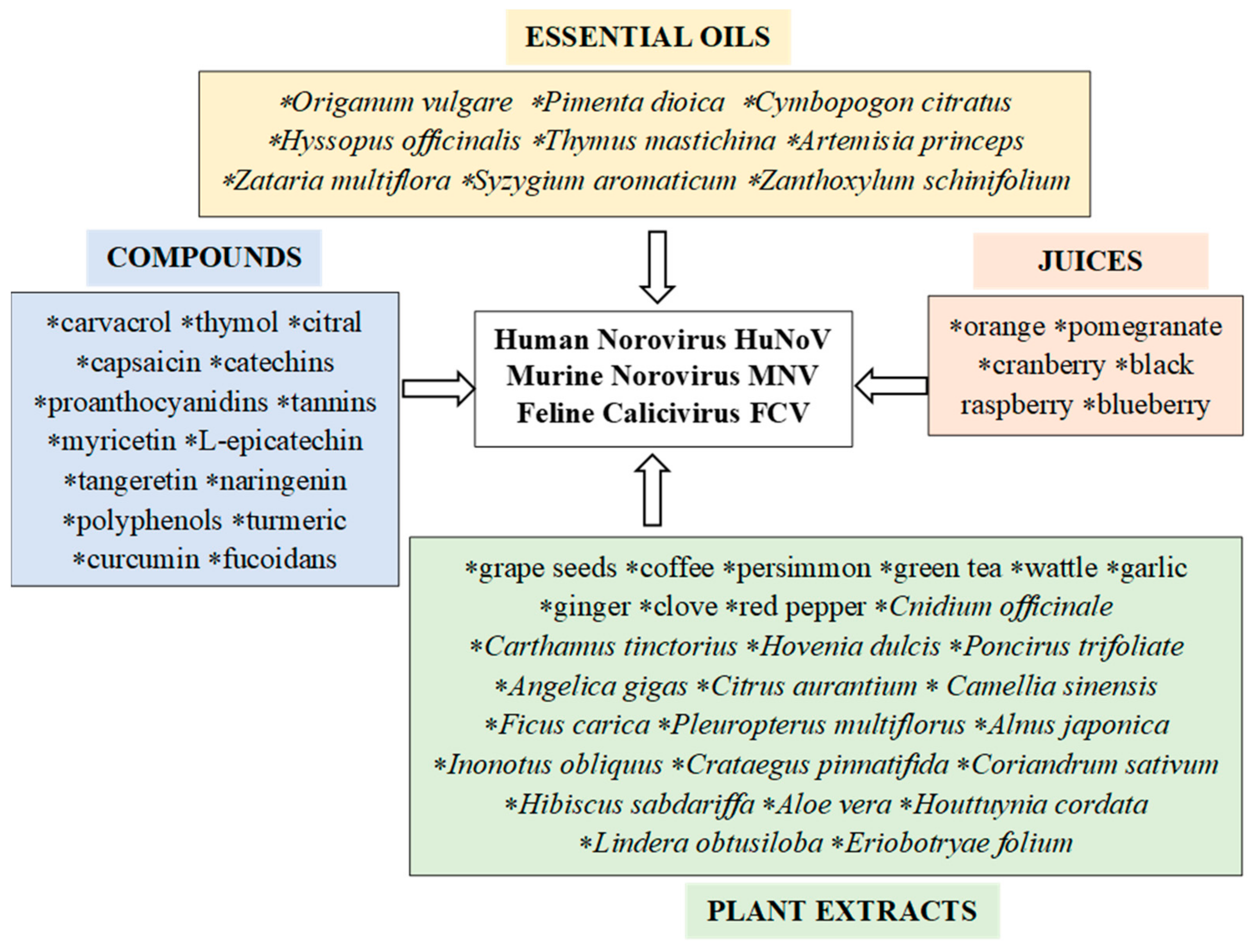
5. Plant Preparations as Antiviral Agents against Noroviruses
5.1. Effect of Essential Oils on Noroviruses
5.2. Effect of Plant Extracts on Noroviruses
5.3. Effect of Bioactive Plant Compounds on Noroviruses
5.4. The Effect of Juices on Noroviruses
6. Practical Application of Metabolites of Plant Origin in the Food Industry
7. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miranda, R.C.; Schaffner, D.W. Virus risk in the food supply chain. Curr. Opin. Food Sci. 2019, 30, 43–48. [Google Scholar] [CrossRef]
- Sanchez, G.; Bosch, A. Survival of enteric viruses in the environment and food. Viruses Foods 2016, 26, 367–392. [Google Scholar]
- Ahmed, S.M.; Hall, A.J.; Robinson, A.E.; Verhoef, L.; Premkumar, P.; Parashar, U.D.; Koopmans, M.; Lopman, B.A. Global prevalence of norovirus in cases of gastroenteritis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 725–730. [Google Scholar] [CrossRef] [Green Version]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; de Silva, N.R.; Gargouri, N.; et al. World Health Organization global estimates and regional comparisons of the burden of foodborne disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartsch, S.M.; Lopman, B.A.; Ozawa, S.; Hall, A.J.; Lee, B.Y. Global economic burden of norovirus gastroenteritis. PLoS ONE 2016, 11, e0151219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Estimates of the Global Burden of Foodborne Diseases; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Pires, S.M.; Fischer-Walker, C.L.; Lanata, C.F.; Devleesschauwer, B.; Hall, A.J.; Kirk, M.D.; Duarte, A.S.R.; Black, R.E.; Angulo, F.J. Aetiology-specific estimates of the global and regional incidence and mortality of diarrhoeal diseases commonly transmitted through food. PLoS ONE 2015, 10, e0142927. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Surveillance for Foodborne Disease Outbreaks United States, 2017: Annual Report. Available online: https://www.cdc.gov/fdoss/pdf/2017_FoodBorneOutbreaks_508.pdf (accessed on 6 June 2020).
- Neethirajan, S.; Ahmed, S.R.; Chand, R.; Buozis, J.; Nagy, E. Recent advances in biosensor development for foodborne virus detection. Nanotheranostics 2017, 1, 272–295. [Google Scholar] [CrossRef] [Green Version]
- EFSA (European Food Safety Authority). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, e5500. [Google Scholar]
- EFSA (European Food Safety Authority). The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, e05926. [Google Scholar]
- EFSA (European Food Safety Authority). The European Union One Health 2019 Zoonoses Report, European Food Safety Authority European Centre for Disease Prevention and Control. EFSA J. 2021, 19, e6406. [Google Scholar]
- Centers for Disease Control and Prevention. Norovirus Worldwide. 2020. Available online: https://www.cdc.gov/nceh/vsp/desc/about_inspections.htm (accessed on 6 June 2020).
- Bosch, A.; Gkogka, E.; Le Guyader, F.S.; Loisy-Hamon, F.; Lee, A.; van Lieshout, L.; Marthi, B.; Myrmel, M.; Sansom, A.; Schultz, A.C.; et al. Foodborne viruses: Detection, risk assessment, and control options in food processing. Int. J. Food Microbiol. 2018, 285, 110–128. [Google Scholar] [CrossRef]
- Pexara, A.; Govaris, A. Foodborne viruses and innovative non-thermal food-processing technologies. Food 2020, 9, 1520. [Google Scholar] [CrossRef] [PubMed]
- Robilotti, E.; Deresinski, S.; Pinsky, B.A. Norovirus. Clin. Microbiol. Rev. 2015, 28, 134–164. [Google Scholar] [CrossRef] [Green Version]
- Callejon, R.M.; Rodriguez-Naranjo, M.I.; Ubeda, C.; Hornedo-Ortega, R.; Garcia-Parrilla, M.C.; Troncoso, A.M. Reported foodborne outbreaks due to fresh produce in the United States and European Union: Trends and causes. Foodborne Pathog. Dis. 2015, 12, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Machado-Moreira, B.; Richards, K.; Brennan, F.; Abram, F.; Burgess, C.M. Microbial contamination of fresh produce: What, where, and how? Compr. Rev. Food Sci. Food Saf. 2019, 18, 1727–1750. [Google Scholar] [CrossRef] [Green Version]
- Lopman, B.A.; Reacher, M.H.; Vipond, I.B.; Sarangi, J.; Brown, D.W. Clinical manifestation of norovirus gastroenteritis in health care settings. Clin. Infect. Dis. 2004, 39, 318–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, M.D.; Goulter, R.M.; Jaykus, L.A. Human norovirus as a foodborne pathogen: Challenges and developments. Annu. Rev. Food Sci. Technol. 2015, 6, 411–433. [Google Scholar] [CrossRef] [PubMed]
- Amankwaah, C.; Li, J.; Lee, J.; Pascall, M.A. Antimicrobial activity of chitosan-based films enriched with green tea extracts on murine norovirus, Escherichia coli, and Listeria innocua. Int. J. Food Sci. 2020, 2, 3941924. [Google Scholar] [CrossRef] [PubMed]
- Rajiuddin, S.M.; Vigre, H.; Musavian, H.S.; Kohle, S.; Krebs, N.; Hansen, T.B.; Gantzer, C.; Schultz, A.C. Inactivation of hepatitis A virus and murine norovirus on surfaces of plastic, steel and raspberries using steam-ultrasound treatment. Food Environ. Virol. 2020, 12, 295–309. [Google Scholar] [CrossRef]
- Solis-Sanchez, D.; Rivera-Piza, A.; Lee, S.; Kim, J.; Kim, B.; Choi, J.B.; Kim, Y.W.; Ko, G.P.; Song, M.J.; Lee, S.J. Antiviral effects of Lindera obtusiloba leaf extract on murine norovirus-1 (MNV-1), a human norovirus surrogate, and potential application to model Foods. Antibiotics 2020, 9, 697. [Google Scholar] [CrossRef]
- Green, K.Y.; Ando, T.; Balayan, M.S.; Berke, T.; Clarke, I.N.; Estes, M.K. Taxonomy of the caliciviruses. J. Infect. Dis. 2000, 181, 322–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinjé, J. Advances in laboratory methods for detection and typing of norovirus. J. Clin. Microbiol. 2015, 53, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Hoa Tran, T.N.; Trainor, E.; Nakagomi, T.; Cunliffe, N.A.; Nakagomi, O. Molecular epidemiology of noroviruses associated with acute sporadic gastroenteritis in children: Global distribution of genogroups, genotypes and GII.4 variants. J. Clin. Virol. 2013, 56, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroneman, A.; Verhoef, L.; Harris, J.; Vennema, H.; Duizer, E.; van Duynhoven, Y.; Gray, J.; Iturriza, M.; Böttiger, B.; Falkenhorst, G.; et al. Analysis of integrated virological and epidemiological reports of norovirus outbreaks collected within the foodborne viruses in Europe network from 1 July 2001 to 30 June 2006. J. Clin. Microbiol. 2008, 46, 2959–2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teunis, P.F.; Moe, C.L.; Liu, P.; Miller, S.E.; Lindesmith, L.; Baric, R.S.; Le Pendu, J.; Calderon, R.L. Norwalk virus: How infectious is it? J. Med. Virol. 2008, 80, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Lysen, M.; Thorhagen, M.; Brytting, M.; Hjertqvist, M.; Andersson, Y.; Hedlund, K.O. Genetic diversity among food-borne and waterborne norovirus strains causing outbreaks in Sweden. J. Clin. Microbiol. 2009, 47, 2411–2418. [Google Scholar] [CrossRef] [Green Version]
- Vega, E.; Barclay, L.; Gregoricus, N.; Shirley, S.H.; Lee, D.; Vinje, J. Genotypic and epidemiologic trends of norovirus outbreaks in the United States, 2009 to 2013. J. Clin. Microbiol. 2014, 52, 147–155. [Google Scholar]
- Hassard, F.; Sharp, J.H.; Taft, H.; LeVay, L.; Harris, J.P.; McDonald, J.E.; Tuson, K.; Wilson, J.; Jones, D.L.; Malham, S.K. Critical review on the public health impact of norovirus contamination in shellfish and the environment: A UK perspective. Food Environ. Virol. 2017, 9, 123–141. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.; Chhabra, V.; Rawal, R.K.; Sharma, S. Chemometrics: A new scenario in herbal drug standardization. J. Pharm. Anal. 2014, 4, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Yang, M.; Lee, G.; Si, J.; Lee, S.J.; You, H.J.; Ko, G. Curcumin shows antiviral properties against norovirus. Molecules 2016, 21, 1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drevinskas, T.; Mickiene, R.; Maruska, A.; Stankevicius, M.; Tiso, N.; Salomskas, A.; Lelesius, R.; Karpovaite, A.; Ragazinskiene, O. Confirmation of antiviral properties of medicinal plants via chemical analysis, machine learning methods and antiviral tests: Methodological approach. Anal. Methods 2018, 10, 1875–1885. [Google Scholar] [CrossRef]
- Lee, H.Y.; Yum, J.H.; Rho, Y.K.; Oh, S.J.; Choi, H.S.; Chang, H.B.; Choi, D.H.; Leem, M.J.; Choi, E.J.; Ryu, J.M.; et al. Inhibition of HCV replicon cell growth by 2-arylbenzofuran derivatives isolated from Mori Cortex Radicis. Planta Med. 2007, 73, 1481–1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggers, M.; Schwebke, I.; Suchomel, M.; Fotheringham, V.; Gebel, J.; Meyer, B.; Morace, G.; Roedger, H.J.; Roques, C.; Visa, P.; et al. The European tiered approach for virucidal efficacy testing-rationale for rapidly selecting disinfectants against emerging and re-emerging viral diseases. Eurosurveillance 2021, 26, 2000708. [Google Scholar] [CrossRef]
- Musarra-Pizzo, M.; Pennisi, R.; Ben-Amor, I.; Mandalari, G.; Sciortino, M.T. Antiviral activity exerted by natural products against human viruses. Viruses 2021, 13, 828. [Google Scholar] [CrossRef]
- Mukherjee, P.K. Antiviral evaluation of herbal drugs. Qual. Control Eval. Herb. Drugs 2019, 599–628. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Lim, C.Y.; Lee, D.B.; Seok, J.H.; Kim, K.H.; Chung, M.S. Inhibitory effects of Laminaria japonica fucoidans against noroviruses. Viruses 2020, 12, 997. [Google Scholar] [CrossRef] [PubMed]
- Van Dycke, J.; Cuvry, A.; Knickmann, J.; Ny, A.; Rakers, S.; Taube, S.; de Witte, P.; Neyts, J.; Rocha-Pereira, J. Infection of zebrafish larvae with human norovirus and evaluation of the In Vivo efficacy of small-molecule inhibitors. Nat. Protoc. 2021, 16, 1830–1849. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural products for drug discovery in the 21st century: Innovations for novel drug discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.J.; Si, J.; Yun, H.S.; Ko, G.P. Effect of temperature and relative humidity on the survival of foodborne viruses during food storage. Appl. Environ. Microb. 2015, 81, 2075–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Lao, Y.; Pan, Y.; Chen, Y.; Zhao, H.; Gong, L.; Xie, N.; Mo, C.H. Synergistic antimicrobial effectiveness of plant essential oil and its application in seafood preservation: A review. Molecules 2021, 26, 307. [Google Scholar]
- Ni, Z.J.; Wang, X.; Shen, Y.; Thakur, K.; Han, J.; Zhang, J.G.; Hu, F.; Wei, Z.J. Recent updates on the chemistry, bioactivities, mode of action, and industrial applications of plant essential oils. Trends Food Sci. Technol. 2021, 110, 78–89. [Google Scholar] [CrossRef]
- Bertrand, I.; Schijven, J.F.; Sanchez, G.; Wyn-Jones, P.; Ottoson, J.; Morin, T.; Muscillo, M.; Verani, M.; Nasser, A.; de Roda Husman, A.M.; et al. The impact of temperature on the inactivation of enteric viruses in food and water: A review. J. Appl. Microbiol. 2012, 112, 1059–1074. [Google Scholar] [CrossRef] [PubMed]
- Ben-Shabat, S.; Yarmolinsky, L.; Porat, D.; Dahan, A. Antiviral effect of phytochemicals from medicinal plants: Applications and drug delivery strategies. Drug Deliv. Transl. Res. 2020, 10, 354–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andricoplo, A.D.; Ceron-Carrasco, J.P.; Mozzarelli, A. Bridging molecular docking to molecular dynamics in exploring ligand-protein recognition process: An overview. Front. Pharmacol. 2018, 9, 438. [Google Scholar]
- Perez, R.M. Antiviral activity of compounds isolated from plants. Pharm. Biol. 2003, 41, 107–157. [Google Scholar] [CrossRef]
- Seo, D.J.; Jeon, S.B.; Oh, H.; Lee, B.H.; Lee, S.Y.; Oh, S.H.; Jung, J.Y.; Choi, C. Comparison of the antiviral activity of flavonoids against murine norovirus and feline calicivirus. Food Control 2016, 60, 25–30. [Google Scholar] [CrossRef]
- Randazzo, W.; Falco, I.; Aznar, R.; Sanchez, G. Effect of green tea extract on enteric viruses and its application as natural sanitizer. Food Microbiol. 2017, 66, 150–156. [Google Scholar] [CrossRef]
- Gilling, D.H.; Kitajima, M.; Torrey, J.T.; Bright, K.R. Antiviral efficacy and mechanisms of action of oregano essential oil and its primary component carvacrol against murine norovirus. J. Appl. Microbiol. 2014, 116, 1149–1163. [Google Scholar] [CrossRef]
- Gilling, D.H.; Kitajima, M.; Torrey, J.T.; Bright, K.R. Mechanisms of antiviral action of plant antimicrobials against murine norovirus. Appl. Environ. Microbiol. 2014, 80, 4898–4910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, X.; Howell, A.B.; D’Souza, D.H. The effect of cranberry juice and cranberry proanthocyanidins on the infectivity of human enteric viral surrogates. Food Microbiol. 2010, 27, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Pilau, M.R.; Alves, S.H.; Weiblen, R.; Arenhart, S.; Cueto, A.P.; Lovato, L.T. Antiviral activity of the Lippia graveolens (Mexican oregano) essential oil and its main compound carvacrol against human and animal viruses. Braz. J. Microbiol. 2011, 42, 1616–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovač, K.; Diez-Valcarce, M.; Raspor, P.; Hernández, M.; Rodríguez-Lázaro, D. Natural plant essential oils do not inactivate non-enveloped enteric viruses. Food Environ. Virol. 2012, 4, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Cliver, D.O. Capsid and infectivity in virus detection. Food Environ. Virol. 2009, 1, 123–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tubiana, T.; Boulard, Y.; Bressanelli, S. Dynamics and asymmetry in the dimer of the norovirus major capsid protein. PLoS ONE 2017, 12, e0182056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, X.; Sangster, M.Y.; D’Souza, D.H. Time-dependent effects of pomegranate juice and pomegranate polyphenols on foodborne virus reduction. Foodborne Pathog. Dis. 2011, 8, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Baert, L.; Zhang, D.; Xia, M.; Zhong, W.; Van Coillie, E.; Xiang, J.; Uyttendaele, M. The effect of grape seed extract on human norovirus GII.4 and murine norovirus-1 in viral suspensions, on stainless steel discs, and in lettuce wash water. Appl. Environ. Microbiol. 2012, 78, 7572–7578. [Google Scholar] [CrossRef] [Green Version]
- Koch, C.; Reichling, J.; Schneele, J.; Schnitzler, P. Inhibitory effect of essential oils against herpes simplex virus type 2. Phytomedicine 2008, 15, 71–78. [Google Scholar] [CrossRef]
- Essoil Database. Available online: http://www.nipgr.ac.in/Essoildb/ (accessed on 6 May 2021).
- Leyva-López, N.; Gutiérrez-Grijalva, E.P.; Vazquez-Olivo, G.; Heredia, J.B. Essential oils of oregano: Biological activity beyond their antimicrobial properties. Molecules 2017, 22, 989. [Google Scholar] [CrossRef] [Green Version]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial properties of plant essential oils against human pathogens and their mode of action: An updated review. Evid. Based Complement. Altern. Med. 2016, 2016, 3012462. [Google Scholar] [CrossRef]
- Adam, K.; Sivropoulou, A.; Kokkini, S.; Lanaras, T.; Arsenakis, M. Antifungal activities of Origanum vulgare subsp. hirtum, Mentha spicata, Lavandula angustifolia, and Salvia fruticosa essential oils against human pathogenic fungi. J. Agric. Food Chem. 1998, 46, 1739–1745. [Google Scholar] [CrossRef]
- Elizaquivel, P.; Azizkhani, M.; Aznar, R.; Sanchez, G. The effect of essential oils on norovirus surrogates. Food Control 2013, 32, 275–278. [Google Scholar] [CrossRef]
- Cutillas, A.B.; Carrasco, A.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Thymus mastichina L. essential oils from Murcia (Spain): Composition and antioxidant, antienzymatic and antimicrobial bioactivities. PLoS ONE 2018, 13, e0190790. [Google Scholar]
- Rodrigues, M.; Lopes, A.C.; Vaz, F.; Filipe, M.; Alves, G.; Ribeiro, M.P.; Coutinho, P.; Araujo, A.R.T.S. Thymus mastichina: Composition and biological properties with a focus on antimicrobial activity. Pharmaceuticals 2020, 19, 479. [Google Scholar] [CrossRef]
- Fraternale, D.; Giamperi, L.; Ricci, D. Chemical composition and antifungal activity of essential oil obtained from In Vitro plants of Thymus mastichina L. J. Essent. Oil Res. 2003, 15, 278–281. [Google Scholar] [CrossRef]
- Borugă, O.; Jianu, C.; Mişcă, C.; Goleţ, I.; Gruia, A.T.; Horhat, F.G. Thymus vulgaris essential oil: Chemical composition and antimicrobial activity. J. Med. Life 2014, 7, 56–60. [Google Scholar]
- Kryvtsova, M.V.; Salamon, I.; Koscova, J.; Bucko, D.; Spivak, M. Antimicrobial, antibiofilm and biochemichal properties of Thymus vulgaris essential oil against clinical isolates of opportunistic infections. Biosyst. Divers. 2019, 27, 270–275. [Google Scholar] [CrossRef] [Green Version]
- Mahboubi, M.; Heidarytabar, R.; Mahdizadeh, E. Antibacterial activity of Zataria multiflora essential oil and its main components against Pseudomonas aeruginosa. Herba Pol. 2017, 63, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Saei-Dehkordi, S.S.; Tajik, H.; Moradi, M.; Khalighi-Sigaroodi, F. Chemical composition of essential oils in Zataria multiflora Boiss. from different parts of Iran and their radical scavenging and antimicrobial activity. Food Chem. Toxicol. 2010, 48, 1562–1567. [Google Scholar] [CrossRef] [PubMed]
- Eftekhar, F.; Zamani, S.; Yusefzadi, M.; Hadian, J.; Ebrahimi, S.N. Antibacterial activity of Zataria multiflora Boiss essential oil against extended spectrum β lactamase produced by urinary isolates of Klebsiella pneumoniae. Jundishapur J. Microbiol. 2011, 4, S43–S49. [Google Scholar]
- Mahboubi, M.; Bidgoli, F.G. Antistaphylococcal activity of Zataria multiflora essential oil and its synergy with vancomycin. Phytomedicine 2010, 17, 548–550. [Google Scholar] [CrossRef] [PubMed]
- Naeini, A.R.; Nazeri, M.; Shokri, H. Antifungal activity of Zataria multiflora, Pelargonium graveolens and Cuminum cyminum essential oils towards three species of Malassezia isolated from patients with pityriasis versicolor. J. Mycol. Med. 2011, 21, 87–91. [Google Scholar] [CrossRef]
- Selles, S.M.A.; Kouidri, M.; Belhamiti, B.T.; Amrane, A.A. Chemical composition, In-Vitro antibacterial and antioxidant activities of Syzygium aromaticum essential oil. J. Food Meas. Charact. 2020, 13, 1–7. [Google Scholar] [CrossRef]
- Saeed, A.; Shahwar, D. Evaluation of biological activities of the essential oil and major component of Syzygium aromaticum. J. Anim. Plant Sci. 2015, 25, 1095–1099. [Google Scholar]
- Kizil, S.; Hasimi, N.; Tolan, V.; Kilinc, E.; Karatas, H. Chemical composition, antimicrobial and antioxidant activities of hyssop (Hyssopus officinalis L.) essential oil. Not. Bot. Horti Agrobot. Cluj Napoca 2010, 38, 99–103. [Google Scholar]
- Mahboubi, M.; Haghi, G.; Kazempour, N. Antimicrobial activity and chemical composition of Hyssopus officinalis L. essential oil. J. Biol. Act. Prod. Nat. 2011, 1, 132–137. [Google Scholar]
- Oh, M.; Chung, M.S. Effects of oils and essential oils from seeds of Zanthoxylum schinifolium against foodborne viral surrogates. Evid. Based Complement. Altern. Med. 2014, 8, 135797. [Google Scholar]
- Diao, W.R.; Hu, Q.P.; Feng, S.S.; Li, W.Q.; Xu, J.G. Chemical composition and antibacterial activity of the essential oil from green huajiao (Zanthoxylum schinifolium) against selected foodborne pathogens. J. Agric. Food Chem. 2013, 61, 6044–6049. [Google Scholar] [CrossRef] [PubMed]
- Dharmadasa, R.M.; Abeysinghe, D.C.; Dissanayake, D.M.N.; Fernando, N.S. Leaf essential oil composition, antioxidant activity, total phenolic content and total flavonoid content of Pimenta dioica (L.) Merr (Myrtaceae): A superior quality spice grown in Sri Lanka. Univers. J. Agric. Res. 2015, 3, 49–52. [Google Scholar]
- Mérida-Reyes, M.S.; Muñoz-Wug, M.A.; Oliva-Hernández, B.E.; Gaitán-Fernández, I.C.; Simas, D.L.R.; Ribeiro da Silva, A.J.; Pérez-Sabino, J.F. Composition and antibacterial activity of the essential oil from Pimenta dioica (L.) Merr. from Guatemala. Medicines 2020, 7, 59. [Google Scholar] [CrossRef] [PubMed]
- Milenkovic, A.; Stanojević, J.; Stojanović-Radić, Z.; Pejčić, M.; Cvetkovic, D.; Zvezdanović, J.B.; Stanojević, L. Chemical composition, antioxidative and antimicrobial activity of allspice (Pimenta dioica (L.) Merr.) essential oil and extract. Adv. Technol. 2020, 9, 27–36. [Google Scholar] [CrossRef]
- Majewska, W.; Kozłowska, M.; Gruczyńska-Sękowska, E.; Kowalska, D.; Tarnowska, K. Lemongrass (Cymbopogon citratus) essential oil: Extraction, composition, bioactivity and uses for food preservation—A review. Pol. J. Food Nutr. Sci. 2019, 69, 327–341. [Google Scholar] [CrossRef]
- Premathilake, U.G.A.T.; Wathugala, D.L.; Dharmadasa, R.M. Evaluation of chemical composition and assessment of antimicrobial activities of essential oil of lemongrass (Cymbopogon citratus (DC.) Stapf). Int. J. Minor Fruits Med. Aromat. Plants 2018, 4, 13–19. [Google Scholar]
- Kim, Y.W.; You, H.J.; Lee, S.; Kim, B.; Kim, D.K.; Choi, J.B.; Kim, J.A.; Lee, H.J.; Joo, I.S.; Lee, J.S.; et al. Inactivation of norovirus by lemongrass essential oil using a norovirus surrogate system. J. Food Prot. 2017, 80, 1293–1302. [Google Scholar] [CrossRef] [Green Version]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia L. genus: A review of bioactive essential oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.S. The variation of the major compounds of Artemisia princeps var. orientalis (Pampan) Hara essential oil by harvest year. Korean J. Food Nutr. 2015, 28, 533–543. [Google Scholar] [CrossRef]
- Chung, M.S. Antiviral activities of Artemisia princeps var. orientalis essential oil and its α-thujone against norovirus surrogates. Food Sci. Biotechnol. 2017, 28, 1457–1461. [Google Scholar] [CrossRef]
- Su, X.; D’Souza, D.H. Grape seed extract for foodborne virus reduction on produce. Food Microbiol. 2013, 34, 1–6. [Google Scholar] [CrossRef]
- Joshi, S.S.; Su, X.; D’Souza, D.H. Antiviral effects of grape seed extract against feline calicivirus, murine norovirus, and hepatitis A virus in model food systems and under gastric conditions. Food Microbiol. 2015, 52, 1–10. [Google Scholar] [CrossRef]
- Oh, M.; Bae, S.Y.; Chung, M.S. Mulberry (Morus alba) seed extract and its polyphenol compounds for control of foodborne viral surrogates. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 655–660. [Google Scholar] [CrossRef]
- Ueda, K.; Kawabata, R.; Irie, T.; Nakai, Y.; Tohya, Y.; Sakaguchi, T. Inactivation of pathogenic viruses by plant-derived tannins: Strong effects of extracts from persimmon (Diospyros kaki) on a broad range of viruses. PLoS ONE 2013, 8, e55343. [Google Scholar] [CrossRef]
- Falco, I.; Randazzo, W.; Rodriguez-Diaz, J.; Gozalbo-Rovira, R.; Luque, D.; Aznar, R.; Sanchez, G. Antiviral activity of aged green tea extract in model food systems and under gastric conditions. Int. J. Food Microbiol. 2019, 2, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Falco, I.; Randazzo, W.; Gomez-Mascaraque, L.G.; Aznar, R.; Lopez-Rubio, A.; Sanchez, G. Fostering the antiviral activity of green tea extract for sanitizing purposes through controlled storage conditions. Food Control 2018, 84, 485–492. [Google Scholar] [CrossRef]
- Falco, I.; Díaz-Reolid, A.; Randazzo, W.; Sanchez, G. Green tea extract assisted low-temperature pasteurization to inactivate enteric viruses in juices. Int. J. Food Microbiol. 2020, 334, 108809. [Google Scholar] [CrossRef]
- Randazzo, W.; Costantini, V.; Morantz, E.K.; Vinje, J. Human intestinal enteroids to evaluate human norovirus GII.4 inactivation by aged-green tea. Front. Microbiol. 2020, 18, 1917. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.G.; Kim, K.L.; Shin, S.B.; Son, K.T.; Lee, H.J.; Kim, T.H.; Kim, Y.M.; Cho, E.J.; Kim, D.K.; Lee, E.W.; et al. Antiviral activity of green tea catechins against feline calicivirus as a surrogate for norovirus. Food Sci. Biotechnol. 2013, 22, 593–598. [Google Scholar] [CrossRef]
- Aboubakr, H.A.; Nauertz, A.; Luong, N.T.; Agrawal, S.; El-Sohaimy, S.A.; Youssef, M.M.; Goyal, S.M. In Vitro antiviral activity of clove and ginger aqueous extracts against feline calicivirus, a surrogate for human norovirus. J. Food Prot. 2016, 79, 1001–1012. [Google Scholar] [CrossRef]
- Seo, D.J.; Choi, C. Inhibition of murine norovirus and feline calicivirus by edible herbal extracts. Food Environ. Virol. 2017, 9, 35–44. [Google Scholar] [CrossRef]
- Park, S.Y.; Kang, S.; Ha, S.D. Antimicrobial effects of vinegar against norovirus and Escherichia coli in the traditional Korean vinegared green laver (Enteromorpha intestinalis) salad during refrigerated storage. Int. J. Food Microbiol. 2016, 5, 208–214. [Google Scholar]
- Lee, H.M.; Kim, S.J.; Lee, J.; Park, B.; Yang, J.S.; Ha, S.D.; Choi, C.; Ha, J.H. Capsaicinoids reduce the viability of a norovirus surrogate during kimchi fermentation. LWT 2019, 115, 108460. [Google Scholar] [CrossRef]
- Cheng, D.; Sun, L.; Zou, S.; Chen, J.; Mao, H.; Zhang, Y.; Liao, N.; Zhang, R. Antiviral effects of Houttuynia cordata polysaccharide extract on murine norovirus-1 (MNV-1)-a human norovirus surrogate. Molecules 2019, 24, 1835. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.S.; Dice, L.; D’Souza, D.H. Aqueous extracts of hibiscus sabdariffa calyces decrease hepatitis A virus and human norovirus surrogate titers. Food Environ. Virol. 2015, 7, 366–373. [Google Scholar] [CrossRef]
- Sanchez, C.; Aznar, R.; Sanchez, G. The effect of carvacrol on enteric viruses. Int. J. Food Microbiol. 2015, 192, 72–76. [Google Scholar] [CrossRef]
- Sanchez, G.; Aznar, R. Evaluation of natural compounds of plant origin for inactivation of enteric viruses. Food Environ. Virol. 2015, 7, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.C.; Kim, Y.W.; Ryu, S.; Lee, A.; Lee, J.S.; Song, M.J. Suppression of norovirus by natural phytochemicals from Aloe vera and Eriobotryae folium. Food Control 2017, 73, 1362–1370. [Google Scholar] [CrossRef]
- Su, X.; Howell, A.B.; D’Souza, D.H. Antiviral effects of cranberry juice and cranberry proanthocyanidins on foodborne viral surrogates—A time dependence study In Vitro. Food Microbiol. 2010, 27, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Sangster, M.Y.; D’Souza, D.H. In Vitro effects of pomegranate juice and pomegranate polyphenols on foodborne viral surrogates. Foodborne Pathog. Dis. 2010, 7, 1473–1479. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; D’Souza, D.H. Naturally occurring flavonoids against human norovirus surrogates. Food Environ. Virol. 2013, 5, 97–102. [Google Scholar] [CrossRef]
- Narayanan, A.; Kehn-Hall, K.; Senina, S.; Lundberg, L.; Duyne, R.V.; Guendel, I.; Das, R.; Baer, A.; Bethel, L.; Turell, M.; et al. Curcumin inhibits rift valley fever virus replication in human cells. J. Biol. Chem. 2012, 287, 33198–33214. [Google Scholar] [CrossRef] [Green Version]
- Randazzo, W.; Aznar, R.; Sanchez, G. Curcumin-mediated photodynamic inactivation of norovirus surrogates. Food Environ. Virol. 2016, 8, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Bae, S.Y.; Oh, M.; Seok, J.H.; Kim, S.; Chung, Y.B.; Gowda, K.G.; Mun, J.Y.; Chung, M.S.; Kim, K.H. Antiviral effects of black raspberry (Rubus coreanus) seed extract and its polyphenolic compounds on norovirus surrogates. Biosci. Biotechnol. Biochem. 2016, 80, 1196–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, S.; Howell, A.B.; D’Souza, D.H. Blueberry proanthocyanidins against human norovirus surrogates in model foods and under simulated gastric conditions. Food Microbiol. 2017, 63, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Horm, K.M.; D’Souza, D.H. Survival of human norovirus surrogates in milk, orange, and pomegranate juice, and juice blends at refrigeration (4 °C). Food Microbiol. 2011, 28, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Bae, S.Y.; Lee, J.H.; Cho, K.J.; Kim, K.H.; Chung, M.S. Antiviral effects of black raspberry (Rubus coreanus) juice on foodborne viral surrogates. Foodborne Pathog. Dis. 2012, 9, 915–921. [Google Scholar] [CrossRef]
- Joshi, S.S.; Howell, A.B.; D’Souza, D.H. Reduction of enteric viruses by blueberry juice and blueberry proanthocyanidins. Food Environ. Virol. 2016, 8, 235–243. [Google Scholar] [CrossRef]
- Lee, J.H.; Bae, S.Y.; Oh, M.; Kim, K.H.; Chung, M.S. Antiviral effects of mulberry (Morus alba) juice and its fractions on foodborne viral surrogates. Foodborne Pathog. Dis. 2014, 11, 224–229. [Google Scholar] [CrossRef]
- Ribeiro-Santos, R.; Andrade, M.; de Melo, N.R.; Sanches-Silva, A. Use of essential oils in active food packaging: Recent advances and future trends. Trends Food Sci. Technol. 2017, 61, 132–140. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kumar, P.; Singh, P.; Tripathi, N.N.; Bajpai, V.K. Essential oils: Sources of antimicrobials and food preservatives. Front. Microbiol. 2017, 16, 2161. [Google Scholar] [CrossRef] [Green Version]
- Fabra, M.J.; Falco, I.; Randazzo, W.; Sanchez, G.; Lopez-Rubio, A. Antiviral and antioxidant properties of active alginate edible films containing phenolic extracts. Food Hydrocoll. 2018, 81, 96–103. [Google Scholar] [CrossRef]
- Falco, I.; Flores-Meraz, P.L.; Randazzo, W.; Sanchez, G.; Lopez-Rubio, A.; Fabra, M.J. Antiviral activity of alginate-oleic acid based coatings incorporating green tea extract on strawberries and raspberries. Food Hydrocoll. 2019, 87, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Ju, J.; Chen, X.; Xie, Y.; Yu, H.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. Application of essential oil as a sustained release preparation in food packaging. Trends Food Sci. Technol. 2019, 92, 22–32. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Essential Oil | Plant | Main Constituents | Group of Chemical Compounds | Content (%) | Viruses | References |
|---|---|---|---|---|---|---|
| Oregano | Origanum vulgare | Carvacrol | Monoterpene | 0.3–80.8 | MNV FCV | [53,63,64,65,66,67] |
| Thymol | Sesquiterpene lactone | 0.96–63.7 | ||||
| P-cymene | Related to monoterpene | <0.1–16.94 | ||||
| Gamma-terpinene | Monoterpene | 0.8–21.0 | ||||
| Alpha-terpineol | Monoterpene alcohol | <0.09–12.0 | ||||
| Limonene | Monoterpene | 0.3–0.7 | ||||
| Marjoram | Thymus mastichina | Linalool | Monoterpene alcohol | 24.5–73.5 | MNV-1 | [57,63,68,69,70] |
| 1,8-cineole | Monoterpene deriv. | 9.4–55.6 | ||||
| Beta-pinene | Monoterpene | 0.6–5.9 | ||||
| Alpha-pinene | Monoterpene | 0.9–4.3 | ||||
| Alpha-terpineol | Monoterpene alcohol | 0.9–3.0 | ||||
| Camphor | Monoterpene deriv. | 0.00001–3.0 | ||||
| Limonene | Monoterpene | 0.4–2.1 | ||||
| Thyme | Thymus vulgaris | Thymol | Sesquiterpene lactone | 27.6–100 | MNV-1 | [57,63,71,72] |
| Trans-sabinene hydrate | Monoterpene hydrate | 0.43–39.4 | ||||
| Menthol | Monoterpene alcohol | 1.3–39 | ||||
| Bornyl acetate | Monoterpene | 0.2–25.57 | ||||
| Limonene | Monoterpene | 0.4–24.2 | ||||
| Carvacrol | Monoterpene | 2.0–20.5 | ||||
| Gamma-terpinene | Monoterpene | 0.6–14.9 | ||||
| Zataria multiflora | Zataria multiflora Boiss. | Thymol | Sesquiterpene lactone | 40.8 | MNV FCV | [63,67,73,74,75,76,77] |
| Carvacrol | Monoterpene | 27.8 | ||||
| Ρ-cymene | Related to monoterpene | 8.4 | ||||
| Gamma-terpinene | Monoterpene | 4.0 | ||||
| Beta-caryophyllene | Sesquiterpene | 2.0 | ||||
| Linalol | Monoterpene alcohol | 1.7 | ||||
| Alpha-terpinolene | Monoterpene | 1.3 | ||||
| Clove | Syzygium aromaticum (Eugenia caryophyllus) | Eugenol | Monoterpene deriv. | 86.7 | MNV FCV | [63,67,78,79] |
| Beta-caryophyllene | Sesquiterpene | 3.2 | ||||
| Allo-aromadendrene | Sesquiterpene | 1.3 | ||||
| Alpha-humulene | Sesquiterpene | 0.9 | ||||
| Hyssop | Hyssopus officinalis | Linalool | Monoterpene alcohol | 49.6 | MNV-1 | [57,63,80,81] |
| 1,8-cineole | Monoterpene deriv. | 13.3 | ||||
| Limonene | Monoterpene | 5.4–12.2 | ||||
| Beta-pinene | Monoterpene | 3.0–11.1 | ||||
| Beta-caryophyllene | Sesquiterpene | 1.5–2.8 | ||||
| Isopinocamphone | Bicyclic monoterpenoids | 1.3–43.3 | ||||
| Zanthoxylum schinifolium | Zanthoxylum schinifolium | Estragole | Phenylpropene | 42.0 | MNV-1 FCV-F9 | [63,82,83] |
| Oleic acid | Monounsaturated omega-9 fatty acid | 20.97 | ||||
| Palmitic acid | Saturated fatty acid | 19.86 | ||||
| 2,4-Decadienal | Polyunsaturated fatty aldehyde | 4.87 | ||||
| 2-Undecenal | Aldehyde | 3.81 | ||||
| Allspice | Pimenta dioica | Eugenol | Monoterpene deriv. | 45.4–83.68 | MNV | [54,63,84,85,86] |
| Beta-caryophyllene | Sesquiterpene | 2.3–8.9 | ||||
| P-cymene | Related to monoterpene | 1.77–1.78 | ||||
| Terpinolene | Monocyclic monoterpene | 1.23–2.35 | ||||
| Alpha-cadinol | Pseudoguaianolide | 1.0–5.9 | ||||
| Alpha-humulene | Sesquiterpene | 0.88–5.4 | ||||
| Lemongrass | Cymbopogon citratus | Geranial | Monoterpene aldehyde | 32.7–49.9 | MNV-1 | [54,63,87,88,89] |
| Neral | Monoterpene aldehyde | 26.5–38.2 | ||||
| Myrcene | Monoterpene | 1.7–25.3 | ||||
| Nerol | Monoterpene | 0.2–12.5 | ||||
| Geraniol | Monoterpene deriv. | 0.2–10.4 | ||||
| 1,8-cineole | Monoterpene deriv. | 0.2–2.9 | ||||
| Tea tree | Artemisia princeps var. orientalis | 1,8-cineole | Monoterpene deriv. | 2.2–24.3 | MNV-1 FCV-F9 | [63,90,91,92] |
| Borneol | Monoterpene alcohol | 2.1–5.6 | ||||
| Camphor | Monoterpene deriv. | 1.4–38.7 | ||||
| α-terpineol | Monoterpene alcohol | 1.1–9.8 | ||||
| Beta-pinene | Monoterpene | 0.6–11.7 | ||||
| Alpha-pinene | Monoterpene | 0.5–9.7 | ||||
| Beta-caryophyllene | Sesquiterpene | 0.4–10.6 | ||||
| Isoborneol | Monoterpene deriv. | 0.1–20.9 | ||||
| Alpha-thujone | Monoterpene | 0.1–16.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarowska, J.; Wojnicz, D.; Jama-Kmiecik, A.; Frej-Mądrzak, M.; Choroszy-Król, I. Antiviral Potential of Plants against Noroviruses. Molecules 2021, 26, 4669. https://doi.org/10.3390/molecules26154669
Sarowska J, Wojnicz D, Jama-Kmiecik A, Frej-Mądrzak M, Choroszy-Król I. Antiviral Potential of Plants against Noroviruses. Molecules. 2021; 26(15):4669. https://doi.org/10.3390/molecules26154669
Chicago/Turabian StyleSarowska, Jolanta, Dorota Wojnicz, Agnieszka Jama-Kmiecik, Magdalena Frej-Mądrzak, and Irena Choroszy-Król. 2021. "Antiviral Potential of Plants against Noroviruses" Molecules 26, no. 15: 4669. https://doi.org/10.3390/molecules26154669
APA StyleSarowska, J., Wojnicz, D., Jama-Kmiecik, A., Frej-Mądrzak, M., & Choroszy-Król, I. (2021). Antiviral Potential of Plants against Noroviruses. Molecules, 26(15), 4669. https://doi.org/10.3390/molecules26154669