
Biological Actions and Molecular Mechanisms of Sambucus nigra L. in Neurodegeneration: A Cell Culture Approach

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Antioxidant Activity
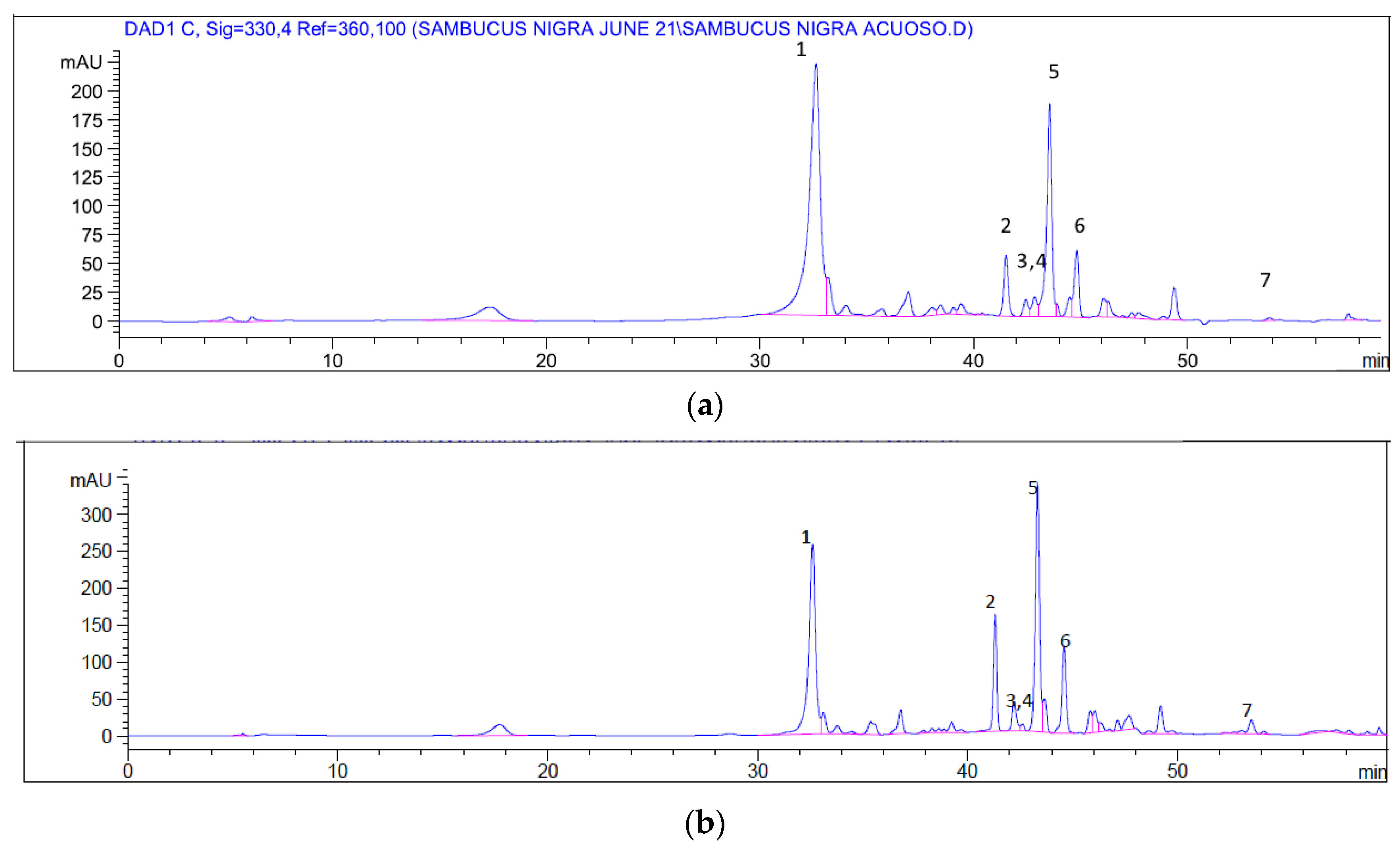
2.2. Chemical Profile
2.3. Cell Viability and mTORC1 Signaling
2.4. ROS Production
2.5. GSH Concentration
2.6. GPx Activity
2.7. GR Activity
2.8. Autophagy
3. Discussion
4. Materials and Methods
4.1. Reagents and Materials
4.2. Sample Preparation
4.3. High-Pressure Liquid Chromatography (HPLC) Analysis
4.4. Antioxidant Capacity and Oxygen Scavenging Activity
4.5. Cell Culture
4.6. Cell Treatment
4.7. Evaluation of Cell Viability
4.8. Determination of ROS
4.9. Determination of Reduced Glutathione (GSH) Concentration
4.10. Determination of Glutathione Peroxidase (GPx) and Glutathione Reductase (GR) Activity
4.11. Western Blotting
4.12. Antibodies
4.13. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- European Medicines Agency Monograph. Assessment Report on Sambucus nigra L., Flos (EMA/HMPC/611504/2016); Committee on Herbal Medicinal Products: London, UK, 2018. [Google Scholar]
- Charlebois, D.; Byers, P.L.; Finn, C.E.; Thomas, A.L. Elderberry: Botany, Horticulture, Potential. Hortic. Rev. 2010, 37, 213–280. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Samoticha, J.; Eler, K.; Stampar, F.; Veberic, R. Traditional Elderflower Beverages: A Rich Source of Phenolic Compounds with High Antioxidant Activity. J. Agric. Food Chem. 2015, 63, 1477–1487. [Google Scholar] [CrossRef]
- Manganelli, R.U.; Zaccaro, L.; Tomei, P. Antiviral activity in vitro of Urtica dioica L., Parietaria diffusa M. et K. and Sambucus nigra L. J. Ethnopharmacol. 2005, 98, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Młynarczyk, K.; Walkowiak-Tomczak, D.; Łysiak, G.P. Bioactive properties of Sambucus nigra L. as a functional ingredient for food and pharmaceutical industry. J. Funct. Foods 2018, 40, 377–390. [Google Scholar] [CrossRef]
- Izzo, A.; Di Carlo, G.; Biscardi, D.; De Fusco, R.; Mascolo, N.; Borrelli, F.; Capasso, F.; Fasulo, M.P.; Autore, G. Biological screening of Italian medicinal plants for antibacterial activity. Phytother. Res. 1995, 9, 281–286. [Google Scholar] [CrossRef]
- Harokopakis, E.; Albzreh, M.H.; Haase, E.; Scannapieco, F.A.; Hajishengallis, G. Inhibition of Proinflammatory Activities of Major Periodontal Pathogens by Aqueous Extracts From Elder Flower (Sambucus nigra). J. Periodontol. 2006, 77, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Gray, A.M.; Abdel-Wahab, Y.; Flatt, P. The Traditional Plant Treatment, Sambucus nigra (elder), Exhibits Insulin-Like and Insulin-Releasing Actions In Vitro. J. Nutr. 2000, 130, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Rebuelta, M.; Vivas, J.M.; Roman, L.S.; Fedez, M.-S. Étude de l’Effet Diurétique de Différentes Préparations des Fleurs du Sambucus Nigra L. Plantes Méd. Phytothér. 1983, 17, 173–181. [Google Scholar]
- Beaux, D.; Fleurentin, J.; Mortier, F. Effect of extracts of Orthosiphon stamineus Benth, Hieracium pilosella L., Sambucus nigra L. and Arctostaphylos uva-ursi (L.) Spreng. in rats. Phytother. Res. 1999, 13, 222–225. [Google Scholar] [CrossRef]
- Viapiana, A.; Wesolowski, M. The Phenolic Contents and Antioxidant Activities of Infusions of Sambucus nigra L. Plant Foods Hum. Nutr. 2017, 72, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Condon, K.J.; Sabatini, D.M. Nutrient regulation of mTORC1 at a glance. J. Cell Sci. 2019, 132, 132. [Google Scholar] [CrossRef]
- Thellung, S.; Corsaro, A.; Nizzari, M.; Barbieri, F.; Florio, T. Autophagy Activator Drugs: A New Opportunity in Neuro-protection from Misfolded Protein Toxicity. Int. J. Mol. Sci. 2019, 20, 901. [Google Scholar] [CrossRef] [Green Version]
- Spilman, P.; Podlutskaya, N.; Hart, M.J.; Debnath, J.; Gorostiza, O.; Bredesen, D.; Richardson, A.; Strong, R.; Galvan, V. Inhibition of mTOR by rapamycin abolishes cognitive deficits and reduces amyloid-beta levels in a mouse model of Alz-heimer’s disease. PLoS ONE 2010, 5, e9979. [Google Scholar] [CrossRef] [Green Version]
- Webb, J.L.; Ravikumar, B.; Atkins, J.; Skepper, J.N.; Rubinsztein, D.C. Alpha-Synuclein is degraded by both autophagy and the proteasome. J. Biol. Chem. 2003, 278, 25009–25013. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Rubinsztein, D.C. Huntington’s disease: Degradation of mutant huntingtin by autophagy. FEBS J. 2008, 275, 4263–4270. [Google Scholar] [CrossRef]
- Kello, M.; Drutovic, D.; Chripkova, M.; Pilatova, M.; Budovska, M.; Kulikova, L.; Urdzik, P.; Mojzis, J. ROS-Dependent Antiproliferative Effect of Brassinin Derivative Homobrassinin in Human Colorectal Cancer Caco2 Cells. Molecules 2014, 19, 10877–10897. [Google Scholar] [CrossRef] [Green Version]
- Schoeneberger, H.; Belz, K.; Schenk, B.; Fulda, S. Impairment of antioxidant defense via glutathione depletion sensitizes acute lymphoblastic leukemia cells for Smac mimetic-induced cell death. Oncogene 2014, 34, 4032–4043. [Google Scholar] [CrossRef] [Green Version]
- Hänsel, R.; Keller, K.; Rimpler, H. Hagers Handbuch der Pharmazeutischen Praxis: Drogen P-Z, 5th ed.; Springer: Berlin/Heidelberg, Germany, 1994; p. 580. [Google Scholar]
- Fleming, T. PDR for Herbal Medicines Medical Economics 2000, 2nd ed.; Medical Economics: North Olmsted, OH, USA, 2000. [Google Scholar]
- Tundis, R.; Ursino, C.; Bonesi, M.; Loizzo, M.R.; Sicari, V.; Pellicano, T.; Manfredi, I.L.; Figoli, A.; Cassano, A. Flower and Leaf Extracts of Sambucus nigra L.: Application of Membrane Processes to Obtain Fractions with Antioxidant and Antityro-sinase Properties. Membranes 2019, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Han, L.; Weng, J.; Wang, K.; Chen, T. Rapamycin inhibits proliferation and induces autophagy in human neuro-blastoma cells. Biosci. Rep. 2018, 38, BSR20181822. [Google Scholar] [CrossRef] [Green Version]
- García-Aguilar, A.; Guillén, C.; Nellist, M.; Bartolomé, A.; Benito, M. TSC2 N-terminal lysine acetylation status affects to its stability modulating mTORC1 signaling and autophagy. Biochim. Biophys. Acta Bioenerg. 2016, 1863, 2658–2667. [Google Scholar] [CrossRef]
- Dewi, F.R.P.; Jiapaer, S.; Kobayashi, A.; Hazawa, M.; Ikliptikawati, D.K.; Hartono; Sabit, H.; Nakada, M.; Wong, R.H.W. Nucleoporin TPR (translocated promoter region, nuclear basket protein) upregulation alters MTOR-HSF1 trails and sup-presses autophagy induction in ependymoma. Autophagy 2021, 17, 1001–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramiro-Puig, E.; Casadesus, G.; Lee, H.-G.; Zhu, X.; McShea, A.; Perry, G.; Pérez-Cano, F.J.; Smith, M.A.; Castell, M. Neuroprotective effect of cocoa flavonids on in vitro oxidative stress. Eur. J. Nutr. 2008, 48, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Bartolomé, A.; García-Aguilar, A.; Asahara, S.-I.; Kido, Y.; Guillén, C.; Pajvani, U.B.; Benito, M. MTORC1 Regulates both General Autophagy and Mitophagy Induction after Oxidative Phosphorylation Uncoupling. Mol. Cell. Biol. 2017, 37, e00441-17. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ramiro, I.; Martín, M.A.; Ramos, S.; Bravo, L.; Goya, L. Comparative effects of dietary flavanols on antioxidant defences and their response to oxidant-induced stress on Caco2 cells. Eur. J. Nutr. 2010, 50, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Martín, M.A.; Ramos, S.; Mateos, R.; Serrano, A.B.G.; Izquierdo-Pulido, M.; Bravo, L.; Goya, L. Protection of Human HepG2 Cells against Oxidative Stress by Cocoa Phenolic Extract. J. Agric. Food Chem. 2008, 56, 7765–7772. [Google Scholar] [CrossRef]
- León-González, A.J.; Mateos, R.; Ramos, S.; Martín, M.A.; Sarriá, B.; Cordero, C.M.; López-Lázaro, M.; Bravo, L.; Goya, L. Chemo-protective activity and characterization of phenolic extracts from Corema album. Food Res. Int. 2012, 49, 728–738. [Google Scholar] [CrossRef] [Green Version]
- Baeza, G.; Amigo-Benavent, M.; Sarriá, B.; Goya, L.; Mateos, R.; Bravo, L. Green coffee hydroxycinnamic acids but not caffeine protect human HepG2 cells against oxidative stress. Food Res. Int. 2014, 62, 1038–1046. [Google Scholar] [CrossRef] [Green Version]
- Martín, M.A.; Ramos, S.; Mateos, R.; Marais, J.P.; Bravo, L.; Khoo, C.; Goya, L. Chemical characterization and chemo-protective activity of cranberry phenolic powders in a model cell culture. Response of the antioxidant defenses and regulation of signaling pathways. Food Res. Int. 2015, 71, 68–82. [Google Scholar] [CrossRef]
- Azam, S.; Hadi, N.; Khan, N.U.; Hadi, S.M. Prooxidant property of green tea polyphenols epicatechin and epigallocate-chin-3-gallate: Implications for anticancer properties. Toxicol. Vitr. 2004, 18, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Farah, A.; Monteiro, M.; Donangelo, C.M.; Lafay, S. Chlorogenic Acids from Green Coffee Extract are Highly Bioavailable in Humans. J. Nutr. 2008, 138, 2309–2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, M.; Farah, A.; Perrone, D.; Trugo, L.C.; Donangelo, C. Chlorogenic Acid Compounds from Coffee Are Differentially Absorbed and Metabolized in Humans. J. Nutr. 2007, 137, 2196–2201. [Google Scholar] [CrossRef] [Green Version]
- Stalmach, A.; Mullen, W.; Barron, D.; Uchida, K.; Yokota, T.; Cavin, C.; Steiling, H.; Williamson, G.; Crozier, A. Metabolite Profiling of Hydroxycinnamate Derivatives in Plasma and Urine after the Ingestion of Coffee by Humans: Identification of Biomarkers of Coffee Consumption. Drug Metab. Dispos. 2009, 37, 1749–1758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M.A.; Serrano, A.B.; Ramos, S.; Pulido, M.I.; Bravo, L.; Goya, L. Cocoa flavonoids up-regulate antioxidant enzyme activity via the ERK1/2 pathway to protect against oxidative stress-induced apoptosis in HepG2 cells. J. Nutr. Biochem. 2010, 21, 196–205. [Google Scholar] [CrossRef]
- De Gouveia, N.M.; Ramos, S.; Martín, M.A.; Espindola, F.S.; Goya, L.; Palomino, O.M. Vochysia rufa Stem Bark Extract Protects Endothelial Cells against High Glucose Damage. Medicines 2017, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomino, O.; Gouveia, N.; Ramos, S.; Martín, M.A.; Goya, L. Protective Effect of Silybum marianum and Silibinin on Endothelial Cells Submitted to High Glucose Concentration. Planta Med. 2016, 83, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Martins, T.F.; Palomino, O.M.; Álvarez-Cilleros, D.; Martín, M.A.; Ramos, S.; Goya, L. Cocoa Flavanols Protect Human Endothelial Cells from Oxidative Stress. Plant Foods Hum. Nutr. 2020, 75, 161–168. [Google Scholar] [CrossRef]
- Castillo-Maldonado, I.; Moreno-Altamirano, M.M.B.; Serrano-Gallardo, L.B. Anti-dengue serotype-2 activity effect of Sam-bucus nigra leaves-and flowers-derived compounds. Virol. Res. Rev. 2017, 1, 1–5. [Google Scholar] [CrossRef]
- Feng, R.; Lu, Y.; Bowman, L.L.; Qian, Y.; Castranova, V.; Ding, M. Inhibition of Activator Protein-1, NF-κB, and MAPKs and Induction of Phase 2 Detoxifying Enzyme Activity by Chlorogenic Acid. J. Biol. Chem. 2005, 280, 27888–27895. [Google Scholar] [CrossRef] [Green Version]
- Pavlica, S.; Gebhardt, R. Protective effects of ellagic and chlorogenic acids against oxidative stress in PC12 cells. Free. Radic. Res. 2005, 39, 1377–1390. [Google Scholar] [CrossRef]
- Granado-Serrano, A.B.; Angeles Martin, M.; Bravo, L.; Goya, L.; Ramos, S. Time-course regulation of quercetin on cell sur-vival/proliferation pathways in human hepatoma cells. Mol. Nutr. Food Res. 2008, 52, 457–464. [Google Scholar] [CrossRef]
- Martin, M.A.; Ramos, S.; Cordero-Herrero, I.; Bravo, L.; Goya, L. Cocoa Phenolic Extract Protects Pancreatic Beta Cells against Oxidative Stress. Nutrients 2013, 5, 2955–2968. [Google Scholar] [CrossRef] [Green Version]
- Ramos, S.; Rodríguez-Ramiro, I.; Martín, M.A.; Goya, L.; Bravo, L. Dietary flavanols exert different effects on antioxidant defenses and apoptosis/proliferation in Caco-2 and SW480 colon cancer cells. Toxicol. Vitr. 2011, 25, 1771–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alía, M.; Ramos, S.; Mateos, R.; Granado-Serrano, A.B.; Bravo, L.; Goya, L. Quercetin protects human hepatoma HepG2 against oxidative stress induced by tert-butyl hydroperoxide. Toxicol. Appl. Pharm. 2006, 212, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhao, L.; Liu, J.; Liu, A.; Jia, C.; Ma, D.; Jiang, Y.; Bai, X. Multi-mechanisms are involved in reactive oxygen species regulation of mTORC1 signaling. Cell. Signal. 2010, 22, 1469–1476. [Google Scholar] [CrossRef]
- Maiese, K.; Chong, Z.Z.; Wang, S.; Shang, Y.C. Oxidant Stress and Signal Transduction in the Nervous System with the PI 3-K, Akt, and mTOR Cascade. Int. J. Mol. Sci. 2012, 13, 13830–13866. [Google Scholar] [CrossRef] [PubMed]
- Filip, G.A.; Florea, A.; Olteanu, D.; Clichici, S.; David, L.; Moldovan, B.; Cenariu, M.; Scrobota, I.; Potara, M.; Baldea, I. Biosynthesis of silver nanoparticles using Sambucus nigra L. fruit extract for targeting cell death in oral dysplastic cells. Mater. Sci. Eng. C 2021, 123, 111974. [Google Scholar] [CrossRef]
- Takahashi, R.; Ono, K.; Takamura, Y.; Mizuguchi, M.; Ikeda, T.; Nishijo, H.; Yamada, M. Phenolic compounds prevent the oligomerization of α-synuclein and reduce synaptic toxicity. J. Neurochem. 2015, 134, 943–955. [Google Scholar] [CrossRef] [Green Version]
- Dávalos, A.; Gómez-Cordovés, A.C.; Bartolomé, B. Extending Applicability of the Oxygen Radical Absorbance Capacity (ORAC−Fluorescein) Assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef]
- Sharma, O.P.; Bhat, T.K. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Browne, R.W.; Armstrong, D. Reduced Glutathione and Glutathione Disulfide. Free Radic. Antioxid. Protoc. 1998, 108, 347–352. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | ORAC µmol TE/mg | IC50 (mg/mL) | AE |
|---|---|---|---|
| Aqueous | 1.38 b ± 0.36 | 8.17 a ± 0.30 | 0.122 a ± 0.004 |
| Ethanolic | 1.13 b ± 0.28 | 7.93 a ± 0.55 | 0.126 a ± 0.009 |
| Methanolic | 0.66 a ± 0.03 | 11.66 b ± 2.05 | 0.089 b ± 0.022 |
| Compound | Retention Time (min) | Content (mg/mL Extract) | |
|---|---|---|---|
| Aqueous Extract | Ethanolic Extract | ||
| 1 Myricetin | 32.61 | 12.61 × 10−3 ± 0.76 | 9.05 × 10−3 ± 0.65 |
| 2. Quercetin | 40.15 | 0.45 × 10−3 ± 0.05 | 1.65 × 10−3 ± 0.08 |
| 3. Caffeic acid | 41.48 | 1.67 × 10−3 ± 0.09 | 0.53 × 10−3 ± 0.01 |
| 4. Chlorogenic acid | 42.42 | 0.91 × 10−3 ± 0.01 | 0.95 × 10−3 ± 0.03 |
| 5. Protocateuchic acid | 43.56 | 4.87 × 10−3 ± 0.08 | 7.01 × 10−3 ± 0.12 |
| 6. Rutin | 48.50 | 0.70 × 10−3 ± 0.01 | 1.12 × 10−3 ± 0.04 |
| 7. Kaempferol | 55.00 | 0.91 × 10−3 ± 0.02 | 2.50 × 10−3 ± 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palomino, O.; García-Aguilar, A.; González, A.; Guillén, C.; Benito, M.; Goya, L. Biological Actions and Molecular Mechanisms of Sambucus nigra L. in Neurodegeneration: A Cell Culture Approach. Molecules 2021, 26, 4829. https://doi.org/10.3390/molecules26164829
Palomino O, García-Aguilar A, González A, Guillén C, Benito M, Goya L. Biological Actions and Molecular Mechanisms of Sambucus nigra L. in Neurodegeneration: A Cell Culture Approach. Molecules. 2021; 26(16):4829. https://doi.org/10.3390/molecules26164829
Chicago/Turabian StylePalomino, Olga, Ana García-Aguilar, Adrián González, Carlos Guillén, Manuel Benito, and Luis Goya. 2021. "Biological Actions and Molecular Mechanisms of Sambucus nigra L. in Neurodegeneration: A Cell Culture Approach" Molecules 26, no. 16: 4829. https://doi.org/10.3390/molecules26164829
APA StylePalomino, O., García-Aguilar, A., González, A., Guillén, C., Benito, M., & Goya, L. (2021). Biological Actions and Molecular Mechanisms of Sambucus nigra L. in Neurodegeneration: A Cell Culture Approach. Molecules, 26(16), 4829. https://doi.org/10.3390/molecules26164829