
Phenolics Profiling of Carpobrotus edulis (L.) N.E.Br. and Insights into Molecular Dynamics of Their Significance in Type 2 Diabetes Therapy and Its Retinopathy Complication




Abstract
:1. Introduction
2. Results and Discussion
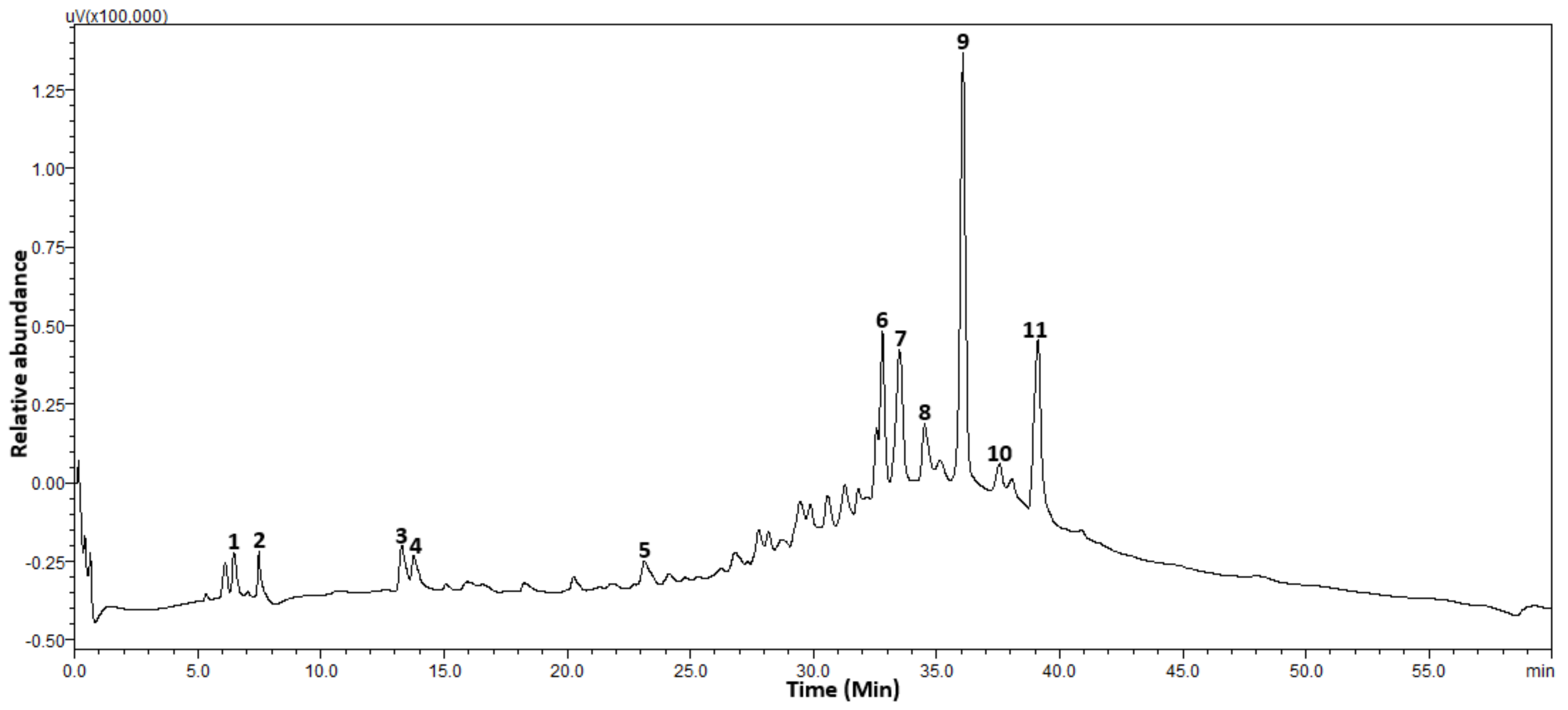
2.1. Phenolic Content
2.2. Antidiabetic Activity
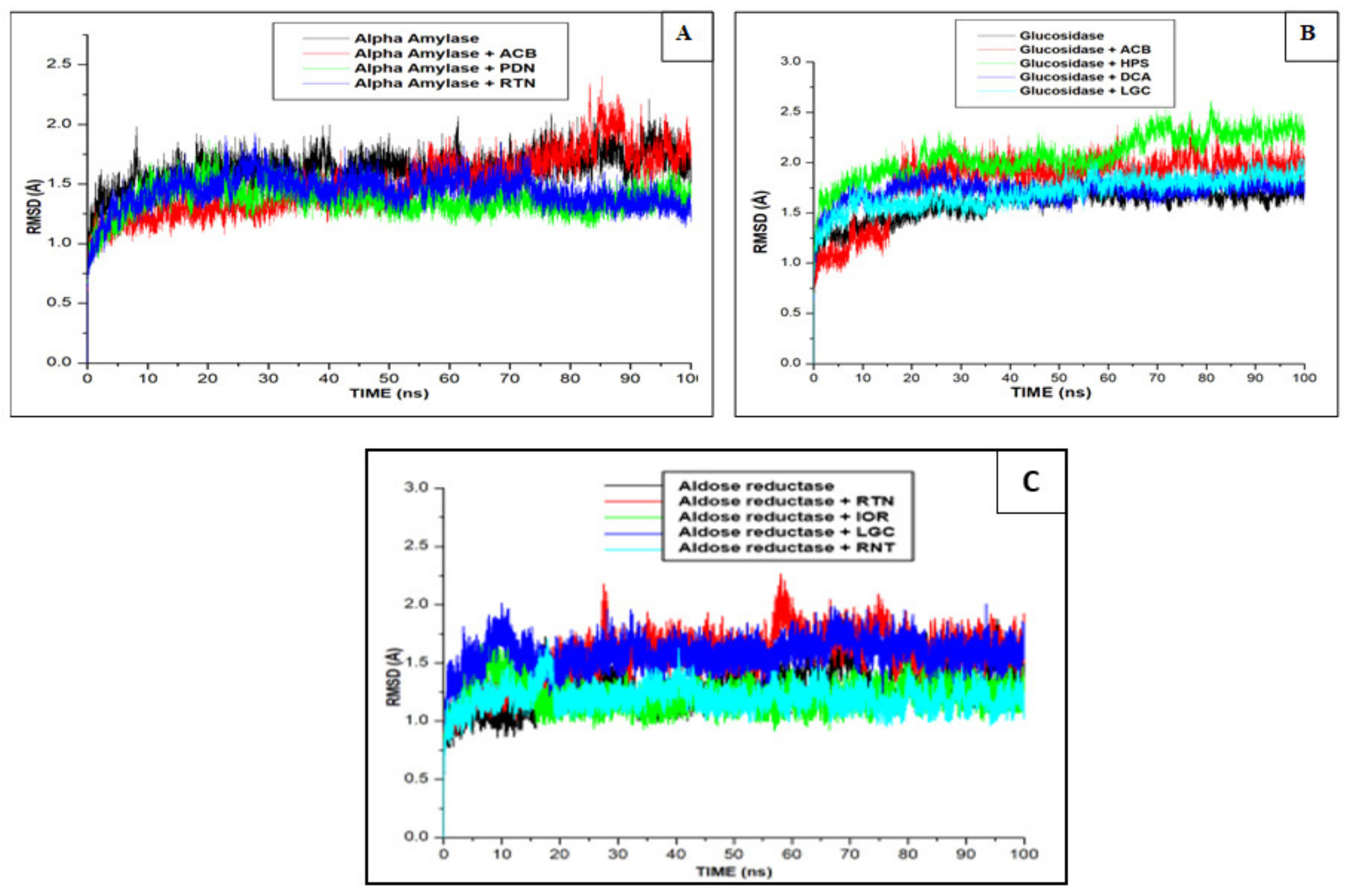
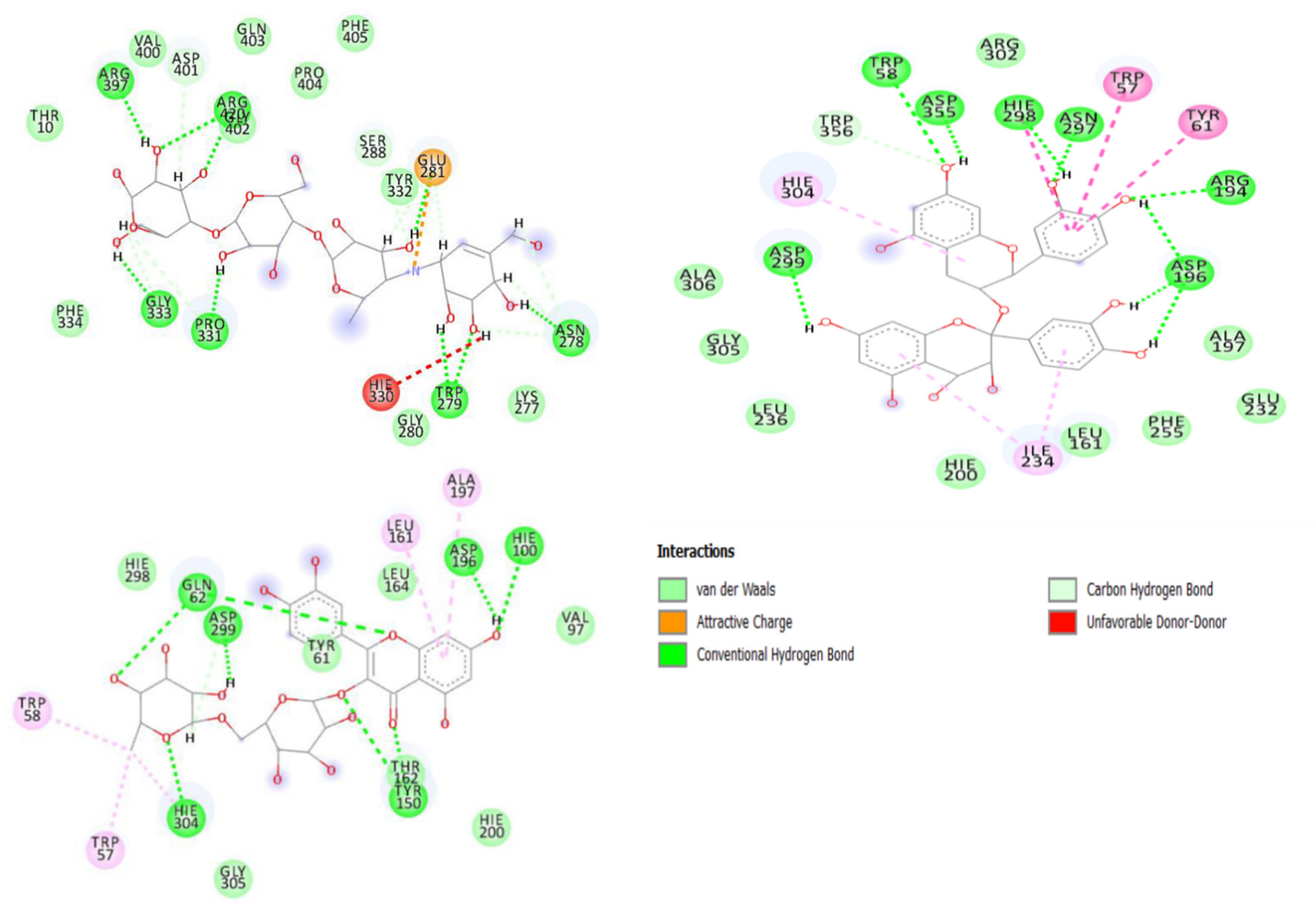
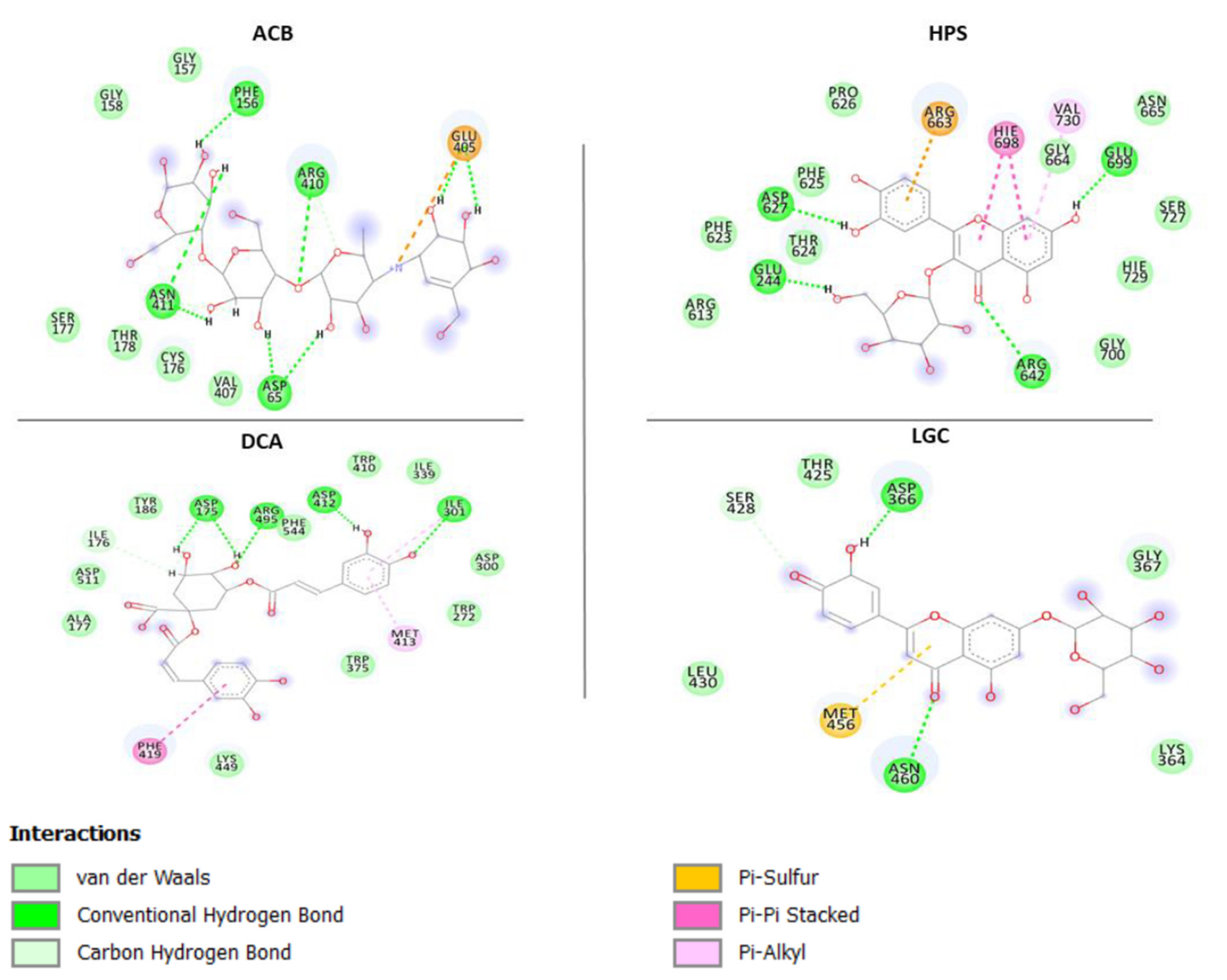
2.3. Molecular Docking and Dynamics
3. Materials and Methods
3.1. Phenolic Extract Preparation and Quantification
3.2. HPLC Analysis
3.3. In vitro Assays
3.3.1. Alpha-Amylase Inhibitory Assay
3.3.2. Alpha-Glucosidase Inhibitory Assay
3.3.3. Aldose Reductase Determination
3.4. In Silico Analysis
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of type 2 diabetes mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- WHO. The Top 10 Causes of Death. 2020. Available online: www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 3 August 2021).
- Sabiu, S.; Ajani, E.O.; Aladodo, R.A.; Garuba, T.; Agunbiade, M.O.; Alimi, A.A.; Lekena, N.; Abdulrazaq, M. Membrane stabilization and probable mechanisms of hypoglycemic activity of fruit extract of Solanum incanum L. (Solanaceae). Comp. Clin. Pathol. 2018, 27, 1611–1619. [Google Scholar] [CrossRef]
- Sabiu, S.; Madende, M.; Ajao, A.A.; Ogundeji, O.A.; Lekena, N.; Alayande, K.A. The scope of phytotherapy in southern African antidiabetic healthcare. Trans. R. Soc. S Afr. 2019, 74, 1–18. [Google Scholar] [CrossRef]
- Kalita, D.; Holm, D.G.; LaBarbera, D.V.; Petrash, J.M.; Jayanty, S.S. Inhibition of α-glucosidase, α-amylase, and aldose reductase by potato polyphenolic compounds. PLoS ONE 2018, 13, e0191025. [Google Scholar] [CrossRef]
- Karasu, C.; Cumaoğlu, A.; Gürpinar, A.R.; Kartal, M.; Kovacikova, L.; Milackova, I.; Stefek, M. Aldose reductase inhibitory activity and antioxidant capacity of pomegranate extracts. Interdiscip. Toxicol. 2012, 5, 15–20. [Google Scholar] [CrossRef]
- Asgar, M.D.A. Anti-diabetic potential of phenolic compounds: A review. Int. J. Food Prop. 2013, 16, 91–103. [Google Scholar] [CrossRef]
- Dias, T.R.; Alves, M.G.; Casal, S.; Oliveira, P.F.; Silva, B.M. Promising potential of dietary (poly) phenolic compounds in the prevention and treatment of diabetes mellitus. Curr. Med. Chem. 2017, 24, 334. [Google Scholar] [PubMed]
- Cavender, G.; Liu, M.; Fernandez-Salvador, J.; Hobbs, D.; Strik, B.; Frei, B.; Zhao, Y. Effect of different commercial fertilizers, harvest date, and storage time on two organically grown blackberry cultivars: Physicochemical properties, antioxidant properties, and sugar profiles. J. Food Qual. 2019, 2019, 1390358. [Google Scholar] [CrossRef]
- Mudimba, T.N.; Nguta, J.M. Traditional uses, phytochemistry and pharmacological activity of Carpobrotus edulis: A global perspective. J. Phytopharmacol. 2019, 8, 111–116. [Google Scholar] [CrossRef]
- Aryaeian, N.; Sedehi, S.K.; Arablou, T. Polyphenols and their effects on diabetes management: A review. Med. J. Islam. Repub. Iran 2017, 31, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WFO. Carpobrotus N.E. Br. 2021. Available online: http://www.worldfloraonline.org/taxon/wfo-4000006785 (accessed on 2 July 2021).
- Malan, C.; Notten, A. Carpobrotus edulis (L.) L.Bolus. SANBI PlantZAfrica.com. Available online: http://pza.sanbi.org/carpobrotus-edulis (accessed on 29 January 2021).
- Mulaudzi, R.B.; Aremu, A.O.; Rengasamy, K.R.R.; Adebayo, S.A.; McGaw, L.J.; Amoo, S.O.; Van Staden, J.; Du Plooy, C.P. Antidiabetic, anti-inflammatory, anticholinesterase and cytotoxicity determination of two Carpobrotus species. S Afr. J. Bot. 2019, 125, 142–148. [Google Scholar] [CrossRef]
- Rehana, D.; Mahendiran, D.; Kumar, R.S.; Rahiman, A.K. In vitro antioxidant and antidiabetic activities of zinc oxide nanoparticles synthesized using different plant extracts. Bioprocess Biosyst. Eng. 2017, 40, 943–957. [Google Scholar] [CrossRef]
- Balogun, F.O.; Ashafa, A.O.T. Aqueous roots extract of Dicoma anomala (Sond.) extenuates postprandial hyperglycaemia in vitro and its modulation against on the activities of carbohydrate-metabolizing enzymes in streptozotocin-induced diabetic Wistar Rats. S. Afr. J. Bot. 2017, 112, 102–111. [Google Scholar] [CrossRef]
- Ahmad, J.B.; Ajani, E.O.; Sabiu, S. Chemical group profiling, in vitro and in silico evaluation of Aristolochia ringens on α-amylase and α-glucosidase activity. Evid. Based Complement Altern. Med. 2021, 6679185. [Google Scholar] [CrossRef]
- Rasouli, H.; Hosseini-Ghazvini, S.M.; Adibi, H.; Khodarahmi, R. Differential α-amylase/α-glucosidase inhibitory activities of plant-derived phenolic compounds: A virtual screening perspective for the treatment of obesity and diabetes. Food Funct. 2017, 24, 1942–1954. [Google Scholar] [CrossRef] [PubMed]
- Grewal, A.S.; Sharma, N.; Singh, S.; Arora, S. Molecular docking evaluation of some natural phenolic compounds as aldose reductase inhibitors for diabetic complications. J. Pharm. Technol. Res. Mgt. 2017, 5, 135–148. [Google Scholar] [CrossRef]
- Aronson, J.K. Aldose reductase inhibitors. In Meyler’s Side Effects of Drugs, 16th ed.; Elsevier Publishers: Amsterdam, The Netherlands, 2016; p. 131. [Google Scholar]
- De la Fuente, J.A.; Manzanaro, S. Aldose reductase inhibitors from natural sources. J. Nat. Prod. Rep. 2003, 20, 243–251. [Google Scholar] [CrossRef]
- Lim, S.S.; Jung, Y.J.; Hyun, S.K.; Lee, Y.S.; Choi, J.S. Rat lens aldose reductase inhibitory constituents of Nelumbo nucifera stamens. J. Phytother. Res. 2006, 20, 825–830. [Google Scholar] [CrossRef]
- Oates, P.J. Aldose reductase, still a compelling target for diabetic neuropathy. Curr. Drug Targets 2008, 9, 14–36. [Google Scholar] [CrossRef]
- Jung, H.A.; Islam, M.N.; Kwon, Y.S.; Jin, S.E.; Son, Y.K.; Park, J.J.; Sohn, H.S.; Choi, J.S. Extraction and identification of three major aldose reductase inhibitors from Artemisia montana. Food Chem. Toxicol. 2011, 49, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Demir, Y.; Durmaz, L.; Taslimi., P.; Gulcin, I. Antidiabetic properties of dietary phenolic compounds: Inhibition effects on α-amylase, aldose reductase, and α-glycosidase. Biotechnol. Appl. Biochem. 2019, 66, 781–786. [Google Scholar] [CrossRef]
- Patel, D.K.; Kumar, R.; Sairam, K.; Hemalatha, S. Pharmacologically tested aldose reductase inhibitors isolated from plant sources—A concise report. Chin. Nat. Med. 2012, 10, 388400. [Google Scholar] [CrossRef]
- Idowu, K.; Ramharack, P.; Nlooto, M.; Gordon, M. Molecular dynamic mechanism(s) of inhibition of bioactive antiviral phytochemical compounds targeting cytochrome P450 3A4 and P-glycoprotein. J. Biomol. Struct. Dyn. 2020, 3, 1221–1233. [Google Scholar]
- Shode, F.O.; Idowu, A.S.K.; Uhomoibhi, O.J.; Sabiu, S. Repurposing drugs and identification of inhibitors of integral proteins (spike protein and main protease) of SARS-CoV-2. J. Biomol. Struct. Dyn. 2021. [Google Scholar] [CrossRef]
- Zabidi, N.A.; Ishak, N.A.; Hamid, M.; Ashari, S.E.; Mohammad Latif, M.A. Inhibitory evaluation of Curculigo latifolia on α-glucosidase, DPP (IV) and in vitro studies in antidiabetic with molecular docking relevance to type 2 diabetes mellitus. J. Enzym. Inhib. Med. Chem. 2021, 36, 109–121. [Google Scholar] [CrossRef]
- Salim, B.; Said, G.; Kambouche, N.; Kress, S. Identification of phenolic compounds from Nettle as new candidate inhibitors of main enzymes responsible on type-II diabetes. Curr. Drug Discov. Technol. 2020, 17, 197. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, R.; Binod, P.; Pandey, A. Biological pretreatment of lignocellulosic biomass—An overview. Bioresour. Technol. 2015, 199, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Hess, B. Convergence of sampling in protein simulations. Phys. Rev. 2002, 65, 031910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wu, S.; Zhang, Q.; Yin, Z.; Zhang, L. α-Glucosidase inhibitory effect of anthocyanins from Cinnamomum camphora fruit: Inhibition kinetics and mechanistic insights through in vitro and in silico studies. Int. J. Biol. Macromol. 2020, 143, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Hashim, A.; Khan, M.S.; Khan, M.; Mohd. Baig, M.; Ahmad, S. Antioxidant and α-amylase inhibitory property of Phyllanthus virgatus L.: An in vitro and molecular interaction study. BioMed Res. Int. 2013, 2013, 729393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babine, R.E.; Bender, S.L. Molecular recognition of protein-Ligand complexes: Applications to drug design. Chem. Rev. 1997, 97, 1359–1472. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Meth Enzymol. 1999, 299, 152–178. [Google Scholar]
- Peng, X.T.; Li, Y.N.; Xia, H.; Peng, L.J.; Feng, Y.Q. Rapid and sensitive detection of fipronil and its metabolites in edible oils by solid-phase extraction based on humic acid bonded silica combined with gas chromatography with electron capture detection. J. Sep. Sci. 2016, 39, 2196–2203. [Google Scholar] [CrossRef] [PubMed]
- Elsnoussi, A.H.M.; Mohammad, J.A.S.; Lee, F.A.; Amirin, S.; Sue, H.C.; Soo, C.T.; Mohd, Z.A.; Mun, F.Y. Potent α-glucosidase and α-amylase inhibitory activities of standardized 50% ethanolic extracts and sinensetin from Orthosiphon stamineus Benth as anti-diabetic mechanism. BMC Complement Altern. Med. 2012, 12, 176. [Google Scholar]
- Nakano, T.; Petrash, J.M. Kinetic and spectroscopic evidence for active site inhibition of human aldose reductase. Biochemistry 1996, 27, 11196. [Google Scholar] [CrossRef]
- Branden, G.; Sjogren, T.; Schnecke, V.; Xue, Y. Structure-based ligand design to overcome CYP inhibition in drug discovery projects. Drug Discov. Today 2014, 19, 905–911. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Bryant, S.H. PubChem substance and compound databases. Nucleic Acids Res. 2016, 44, 1202–1213. [Google Scholar] [CrossRef]
- Nair, P.C.; Miners, J.O. Molecular dynamics simulations: From structure function relationships to drug discovery. In Silico Pharmacol. 2014, 2, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonnet, P. P-SHAKE: A quadratically convergent SHAKE in. J. Comput. Phys. 2007, 220, 740–750. [Google Scholar] [CrossRef]
- Ylilauri, M.; Pentika¨inen, O.T. MMGBSA as a tool to understand the binding affinities of filamin–peptide interactions. J. Chem. Inform Model 2013, 53, 2626–2633. [Google Scholar] [CrossRef]
- Sabiu, S.; Idowu, K. An insight on the nature of biochemical interactions between glycyrrhizin, myricetin and CYP3A4 isoform. J. Food Biochem. 2021. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peaks | Retention Time (min) | m/z [M − H]− | Compound |
|---|---|---|---|
| 1 | 6.8 | 223.09 | Sinapic acid |
| 2 | 7.7 | 477.35 | Cacticin |
| 3 | 13.2 | 463.27 | Hyperoside |
| 4 | 13.8 | 515.44 | 1,3-Dicaffeoylquinic acid |
| 5 | 23.2 | 593.50 | Procyanidin |
| 6 | 33.0 | 609.45 | Rutin |
| 7 | 33.7 | 289.08 | Epicatechin |
| 8 | 34.5 | 623.56 | Isorhamnetin-3-O-rutinoside |
| 9 | 36.1 | 353.75 | Chlorogenic acid |
| 10 | 37.4 | 317.19 | Myricetin |
| 11 | 39.1 | 447.13 | Luteolin 7-O-beta-d-glucoside |
| Extract/Compound | Concentration (mg/mL) | ||
|---|---|---|---|
| Alpha-Amylase | Alpha-Glucosidase | Aldose Reductase | |
| C. edulis | 0.51 ± 0.07 a | 0.06 ± 0.01 a | 0.75 ± 0.05 a |
| Acarbose | 0.55 ± 0.09 a | 0.72 ± 0.05 b | NA |
| Ranirestat | NA | NA | 7.05 ± 0.05 b |
| Compounds | Aldose Reductase | α-Amylase | α-Glucosidase |
|---|---|---|---|
| Acarbose (ACB) | ND | −7.7 | −7.5 |
| Ranirestat (RNT) | −8.4 | ND | ND |
| 1,3-Dicaffeoylquinic acid (DCA) | −9.7 | −7.7 | −8.1 |
| Cacticin (CCT) | −7.9 | −8.1 | −7.6 |
| Chlorogenic acid (CGA) | −8.9 | −7.4 | −7.6 |
| Epicatechin (EPT) | −9.6 | −8.4 | −7.7 |
| Hyperoside (HPS) | −7.5 | −7.9 | −8.4 |
| Isorhamnetin-3-O-rutinoside(IOR) | −8.9 | −8.8 | −8.4 |
| Luteolin 7-O-beta-d-glucoside (LGC) | −9.4 | −9.2 | −8.6 |
| Myricetin (MYC) | −8.8 | −7.7 | 8.1 |
| Procyanidin (PDN) | −8.3 | −8.6 | −7.9 |
| Rutin (RTN) | −8.9 | −9.0 | −7.8 |
| Sinapic acid (SPA) | −6.6 | −5.7 | −6.4 |
| Complex | Energy Components (kcal/mol) | ||||
|---|---|---|---|---|---|
| Δ EvdW | ΔEelec | ΔGgas | ΔGsolv | ΔGbind | |
| α-Amylase | |||||
| ACB | −52.578 ± 4.803 | −93.386 ± 12.396 | −145.965 ± 11.568 | 91.286± 9.321 | −54.679 ± 4.890 |
| CCT | −42.042 ± 4.060 | −48.401 ± 2.379 | −90.443 ± 12.273 | 48.248 ± 5.903 | −42.195 ± 8.858 |
| PDN | −45.013 ± 5.091 | −111.131 ± 15.036 | −156.145 ± 13.931 | 86.310 ± 9.183 | −69.834 ± 6.574 |
| RTN | −43.268 ± 4.016 | −65.640 ± 5.205 | −108.908± 12.001 | 62.081 ± 9.760 | −46.826 ± 3.262 |
| α-Glucosidase | |||||
| ACB | −24.164 ± 5.955 | −396.679 ± 30.892 | −420.843 ± 31.177 | 385.092 ± 23.859 | −35.751 ± 9.641 |
| CCT | −19.542 ± 4.245 | −173.993 ± 21.584 | −198.343 ± 23.812 | 162.521 ± 19.321 | −30.857 ± 6.019 |
| HPS | −32.536± 4.673 | −65.783± 9.645 | −98.319± 7.684 | 60.127± 12.513 | −38.192± 6.407 |
| DCA | −34.367 ± 4.263 | −58.595 ± 11.108 | −92.962 ± 9.421 | 50.331 ± 7.343 | −42.630 ± 4.076 |
| LGC | −21.894 ± 3.942 | −183.993 ± 28.654 | −205.887 ± 28.876 | 172.531 ± 23.163 | −33.355 ± 7.119 |
| RTN | −24.254 ± 1.113 | −55.254 ± 5.548 | −87.478 ± 4.548 | 48.323 ± 4.453 | −31.012 ± 2.016 |
| Aldose reductase | |||||
| RNT | −45.149 ± 2.951 | −15.180 ± 3.971 | −60.330 ± 4.869 | 21.823 ± 2.876 | −38.506 ± 3.319 |
| CGA | −45.687 ± 2.949 | −30.610 ± 4.368 | −76.297 ± 5.030 | 34.866 ± 8.519 | −41.431 ± 7.470 |
| EPT | −41.078 ± 2.944 | −34.097 ± 6.898 | −75.177 ± 8.385 | 33.825 ± 5.961 | −41.351 ± 3.745 |
| IOR | −60.937 ± 3.431 | −29.525 ± 4.654 | −90.462 ± 9.270 | 45.064 ± 7.0224 | −45.398 ± 4.568 |
| LGC | −54.243 ± 3.435 | −58.854± 7.995 | −113.098 ± 8.049 | 67.995 ± 6.395 | −45.102 ± 4.024 |
| RTN | −56.737 ± 6.748 | −31.384 ± 5.681 | −88.122 ± 12.366 | 46.000 ± 9.981 | −42.122 ± 4.787 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabiu, S.; Balogun, F.O.; Amoo, S.O. Phenolics Profiling of Carpobrotus edulis (L.) N.E.Br. and Insights into Molecular Dynamics of Their Significance in Type 2 Diabetes Therapy and Its Retinopathy Complication. Molecules 2021, 26, 4867. https://doi.org/10.3390/molecules26164867
Sabiu S, Balogun FO, Amoo SO. Phenolics Profiling of Carpobrotus edulis (L.) N.E.Br. and Insights into Molecular Dynamics of Their Significance in Type 2 Diabetes Therapy and Its Retinopathy Complication. Molecules. 2021; 26(16):4867. https://doi.org/10.3390/molecules26164867
Chicago/Turabian StyleSabiu, Saheed, Fatai O. Balogun, and Stephen O. Amoo. 2021. "Phenolics Profiling of Carpobrotus edulis (L.) N.E.Br. and Insights into Molecular Dynamics of Their Significance in Type 2 Diabetes Therapy and Its Retinopathy Complication" Molecules 26, no. 16: 4867. https://doi.org/10.3390/molecules26164867
APA StyleSabiu, S., Balogun, F. O., & Amoo, S. O. (2021). Phenolics Profiling of Carpobrotus edulis (L.) N.E.Br. and Insights into Molecular Dynamics of Their Significance in Type 2 Diabetes Therapy and Its Retinopathy Complication. Molecules, 26(16), 4867. https://doi.org/10.3390/molecules26164867