
Natural Cryoprotective and Cytoprotective Agents in Cryopreservation: A Focus on Melatonin

 ,
,
Abstract
:1. Introduction
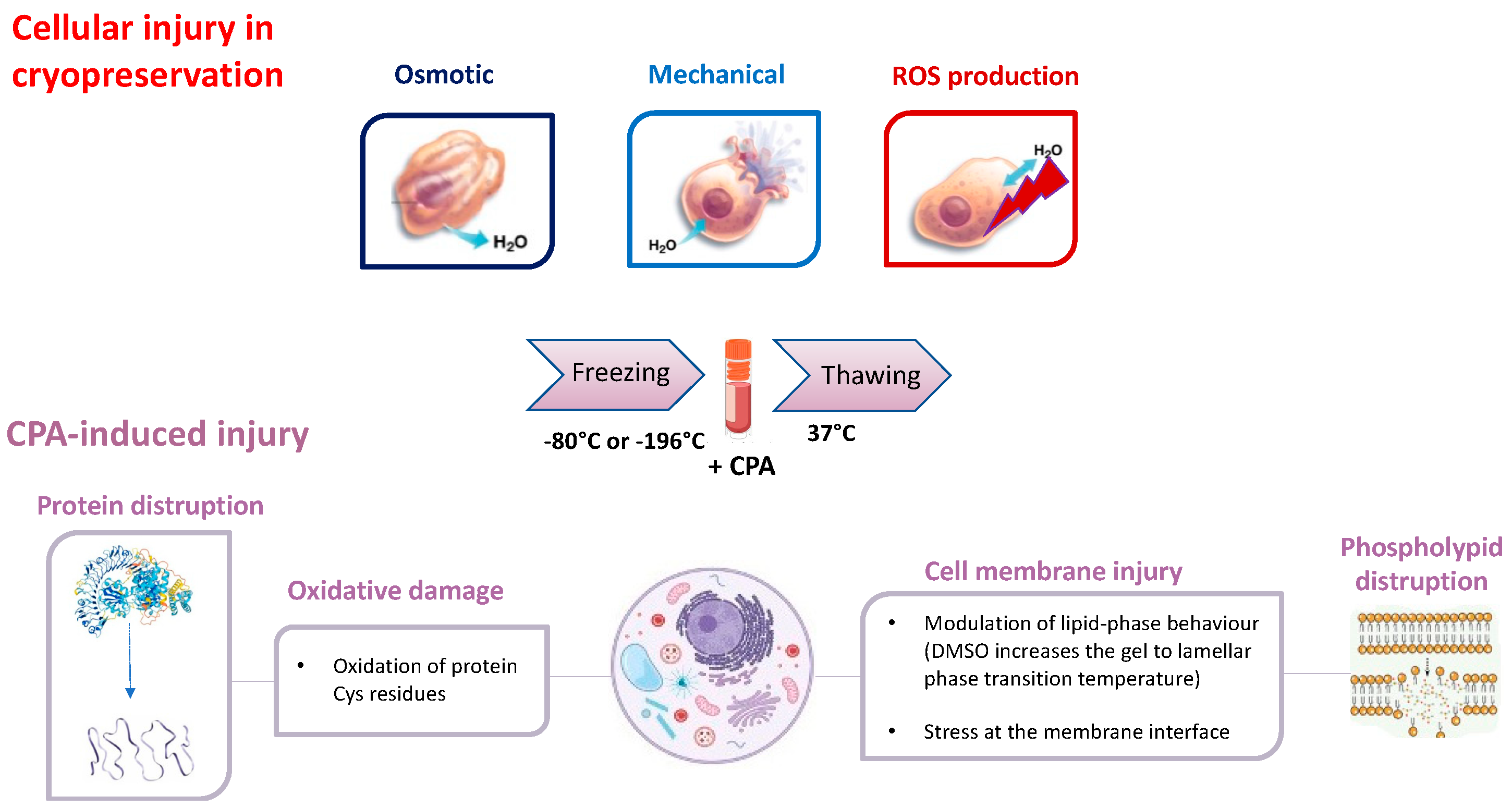
2. The Cryopreservation Process and the Role of CPA
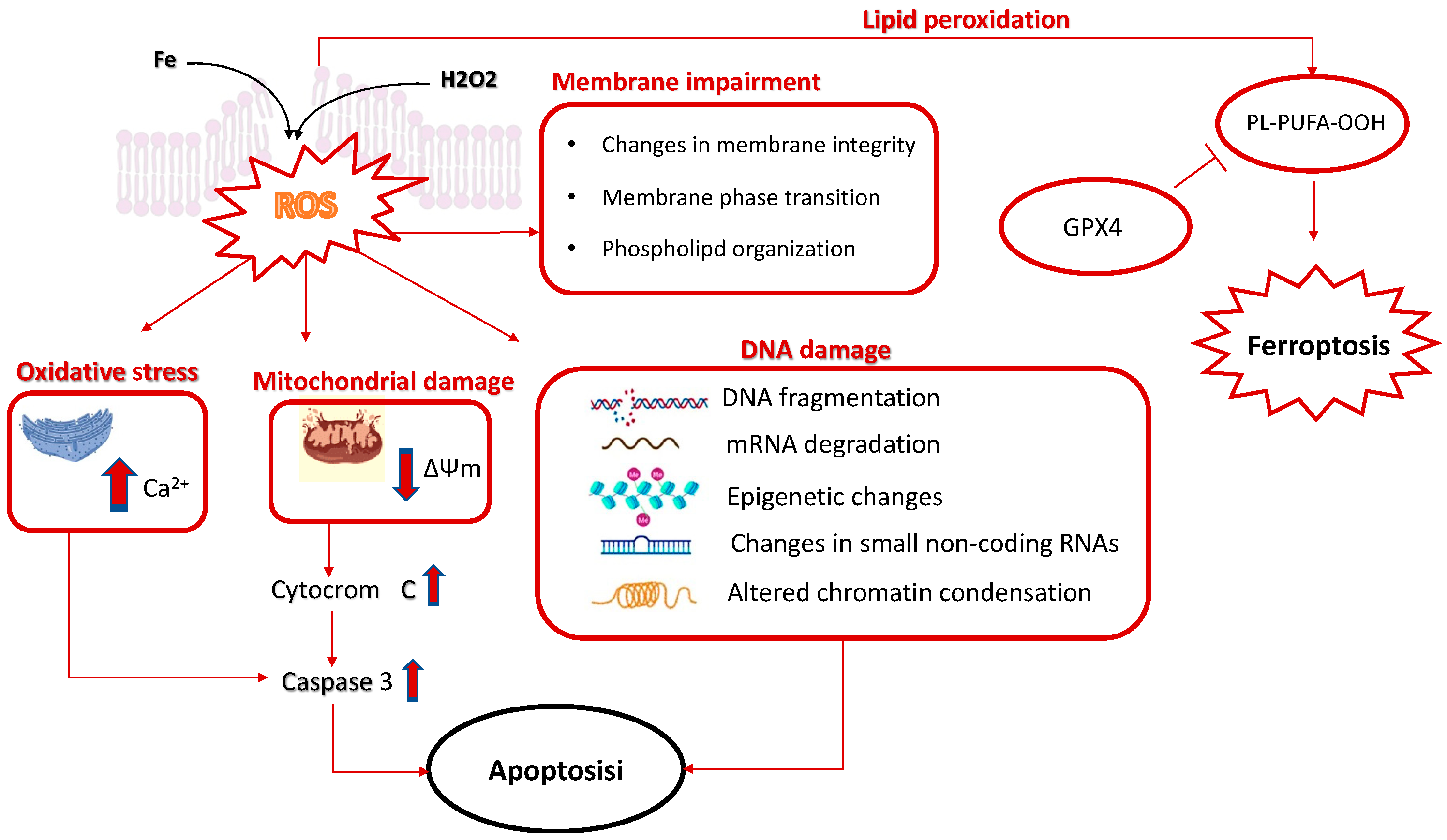
3. Challenges in the Utilization of CPAs
4. Natural Cryoprotective and Cytoprotective Agents
4.1. Antifreeze Proteins (AFPs)
4.2. Sugars
4.3. Natural Deep Eutectic Systems (NADESs)
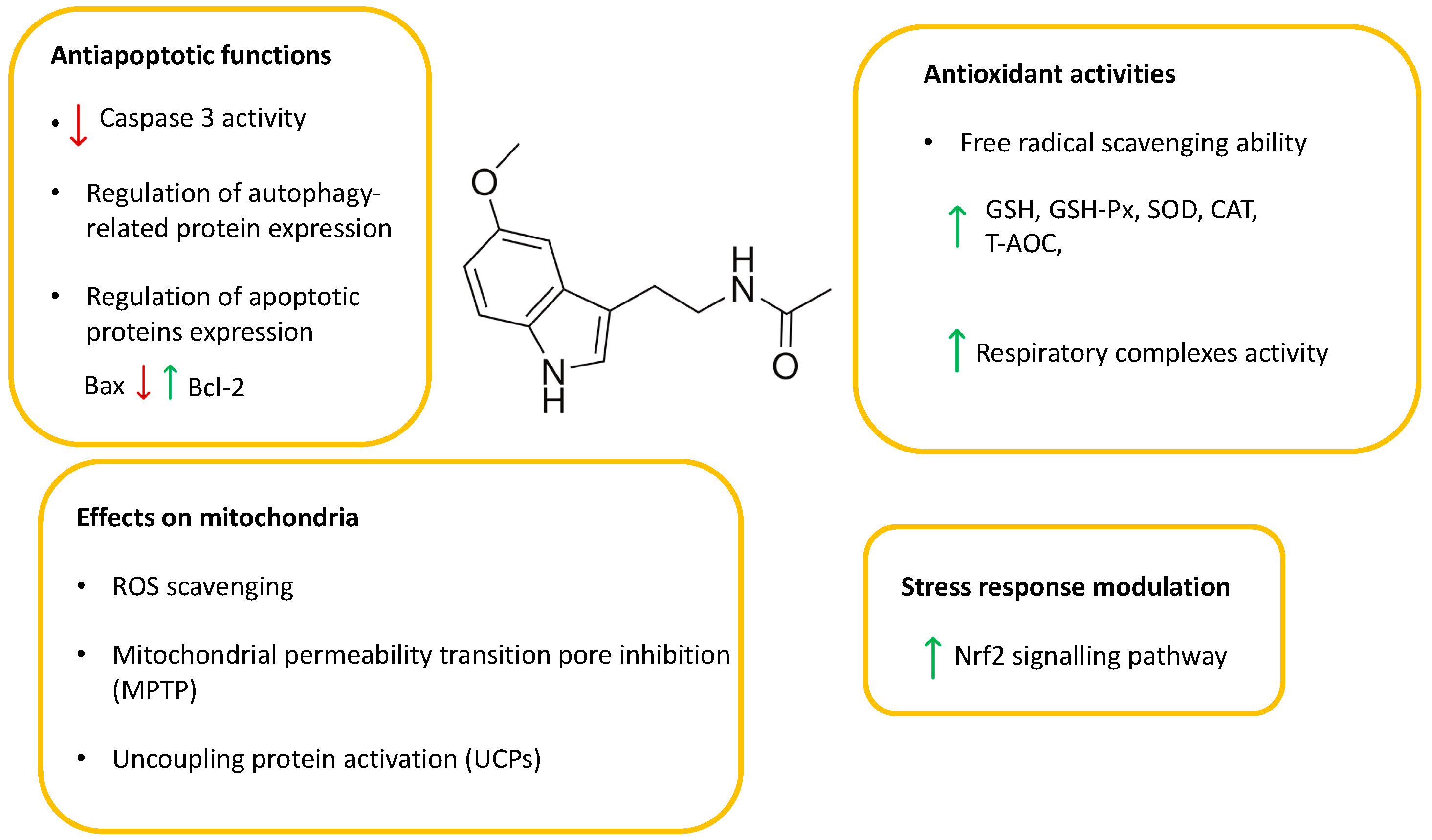
5. The Role of Melatonin (MLT) in Cytoprotection and Cryopreservation
5.1. Ovarian Tissue
5.2. Sperm Cells
6. Melatonin in Stem Cells Cytoprotection
Spermatogonial Stem Cells (SSCs)
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Torquato, P.; Giusepponi, D.; Bartolini, D.; Barola, C.; Marinelli, R.; Sebastiani, B.; Galarini, R.; Galli, F. Pre-analytical monitoring and protection of oxidizable lipids in human plasma (vitamin E and omega-3 and omega-6 fatty acids): An update for redox-lipidomics methods. Free Radic. Biol. Med. 2021, 176, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Pegg, D.E. Principles of cryopreservation. Methods Mol. Biol. 2007, 368, 39–57. [Google Scholar] [PubMed]
- Bojic, S.; Murray, A.; Bentley, B.L.; Spindler, R.; Pawlik, P.; Cordeiro, J.L.; Bauer, R.; de Magalhães, J.P. Winter is coming: The future of cryopreservation. BMC Biol. 2021, 19, 56. [Google Scholar] [CrossRef] [PubMed]
- Uchendu, E.E.; Leonard, S.W.; Traber, M.G.; Reed, B. M Vitamins C and E improve regrowth and reduce lipid peroxidation of blackberry shoot tips following cryopreservation. Plant Cell Rep. 2010, 29, 25–35. [Google Scholar] [CrossRef]
- Banday, M.N.; Lone, F.A.; Rasool, F.; Rashid, M.; Shikari, A. Use of antioxidants reduce lipid peroxidation and improve quality of crossbred ram sperm during its cryopreservation. Cryobiology 2017, 74, 25–30. [Google Scholar] [CrossRef]
- Evangelista-Vargas, S.; Santiani, A. Detection of intracellular reactive oxygen species (superoxide anion and hydrogen peroxide) and lipid peroxidation during cryopreservation of alpaca spermatozoa. Reprod. Domest. Anim. 2017, 52, 819–824. [Google Scholar] [CrossRef]
- Figueroa, E.; Farias, J.; Lee-Estevez, M.; Valdebenito, I.; Risopatrón, J.; Magnotti, C.; Romero, J.; Watanabe, I.; Oliveira, R. Sperm cryopreservation with supplementation of α-tocopherol and ascorbic acid in freezing media increase sperm function and fertility rate in Atlantic salmon (Salmo salar). Aquaculture 2018, 493, 1–8. [Google Scholar] [CrossRef]
- Ball, B.A. Oxidative stress, osmotic stress and apoptosis: Impacts on sperm function and preservation in the horse. Anim. Reprod. Sci. 2008, 107, 257–267. [Google Scholar] [CrossRef]
- Mostek, A.; Dietrich, M.A.; Słowińska, M.; Ciereszko, A. Cryopreservation of bull semen is associated with carbonylation of sperm proteins. Theriogenology 2017, 92, 95–102. [Google Scholar] [CrossRef]
- Ladeira, C.; Koppen, G.; Scavone, F.; Giovannelli, L. The comet assay for human biomonitoring: Effect of cryopreservation on DNA damage in different blood cell preparations. Mutat. Res. Toxicol. Environ. Mutagen. 2019, 843, 11–17. [Google Scholar] [CrossRef]
- Len, J.S.; Koh, W.S.D.; Tan, S.-X. The roles of reactive oxygen species and antioxidants in cryopreservation. Biosci. Rep. 2019, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, G.D.; Wang, S.; Fuller, B.J. Cryoprotectants: A review of the actions and applications of cryoprotective solutes that modulate cell recovery from ultra-low temperatures. Cryobiology 2017, 76, 74–91. [Google Scholar] [CrossRef] [PubMed]
- Crisol, M.; Wu, K.; Laouar, L.; Elliott, J.A.; Jomha, N.M. Antioxidant additives reduce reactive oxygen species production in articular cartilage during exposure to cryoprotective agents. Cryobiology 2020, 96, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.H.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.-X.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef] [PubMed]
- Luchetti, F.; Canonico, B.; Betti, M.; Arcangeletti, M.; Pilolli, F.; Piroddi, M.; Canesi, L.; Papa, S.; Galli, F. Melatonin signaling and cell protection function. FASEB J. 2010, 24, 3603–3624. [Google Scholar] [CrossRef]
- Valerian, E.K.; Gaowei, M.; Feng, Q.; Jose, P.F.A.; Sebastian, D.; Claudette, S.C.; Haider, H.D.; Bing, L.; Vladimir, A.T.; Vladimir, B.R. Oxidized Arachidonic and Adrenic Pes Navigate Cells to Ferroptosis. Nat. Chem. Biol. 2017, 13, 81–90. [Google Scholar]
- Moo-Young, M. Comprehensive Biotechnology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Matsumura, K.; Hayashi, F.; Nagashima, T.; Rajan, R.; Hyon, S.-H. Molecular mechanisms of cell cryopreservation with polyampholytes studied by solid-state NMR. Commun. Mater. 2021, 2, 15. [Google Scholar] [CrossRef]
- Rajan, R.; Hayashi, F.; Nagashima, T.; Matsumura, K. Toward a Molecular Understanding of the Mechanism of Cryopreservation by Polyampholytes: Cell Membrane Interactions and Hydrophobicity. Biomacromolecules 2016, 17, 1882–1893. [Google Scholar] [CrossRef]
- Zhao, J.; Johnson, M.; Fisher, R.B.; Burke, N.A.D.; Stöver, H.D. Synthetic Polyampholytes as Macromolecular Cryoprotective Agents. Langmuir 2019, 35, 1807–1817. [Google Scholar] [CrossRef]
- Nagao, M.; Sengupta, J.; Diaz-Dussan, D.; Adam, M.K.; Wu, M.; Acker, J.P.; Ben, R.; Ishihara, K.; Zeng, H.; Miura, Y.; et al. Synthesis of Highly Biocompatible and Temperature-Responsive Physical Gels for Cryopreservation and 3D Cell Culture. ACS Appl. Bio Mater. 2018, 1, 356–366. [Google Scholar] [CrossRef]
- Tucker, M.; Wright, G.; Morton, P.; Shanguo, L.; Massey, J.; Kort, H. Fertilization and early embryology: Preliminary experience with human oocyte cryopreservation using 1,2-propanediol and sucrose. Hum. Reprod. 1996, 11, 1513–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbri, R.; Porcu, E.; Marsella, T.; Primavera, M.R.; Seracchioli, R.; Ciotti, P.M.; Magrini, O.; Venturoli, S.; Flamigni, C. Oocyte cryopreservation. Hum. Reprod. 1998, 13 (Suppl. 4), 98–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celik, Y.; Drori, R.; Pertaya-Braun, N.; Altan, A.; Barton, T.; Bar-Dolev, M.; Groisman, A.; Davies, P.L.; Braslavsky, I. Microfluidic experiments reveal that antifreeze proteins bound to ice crystals suffice to prevent their growth. Proc. Natl. Acad. Sci. USA 2013, 110, 1309–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raju, R.; Bryant, S.J.; Wilkinson, B.L.; Bryant, G. The need for novel cryoprotectants and cryopreservation protocols: Insights into the importance of biophysical investigation and cell permeability. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129749. [Google Scholar] [CrossRef] [PubMed]
- Erol, O.D.; Pervin, B.; Seker, M.E.; Aerts-Kaya, F. Effects of storage media, supplements and cryopreservation methods on quality of stem cells. World J. Stem Cells 2021, 13, 1197–1214. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Matosevic, S. Cryopreservation of NK and T Cells Without DMSO for Adoptive Cell-Based Immunotherapy. BioDrugs 2021, 35, 529–545. [Google Scholar] [CrossRef] [PubMed]
- Whaley, D.; Damyar, K.; Witek, R.P.; Mendoza, A.; Alexander, M.; Lakey, J.R. Cryopreservation: An Overview of Principles and Cell-Specific Considerations. Cell Transplant. 2021, 30, 963689721999617. [Google Scholar] [CrossRef]
- Ekpo, M.D.; Xie, J.; Hu, Y.; Liu, X.; Liu, F.; Xiang, J.; Zhao, R.; Wang, B.; Tan, S. Antifreeze Proteins: Novel Applications and Navigation towards Their Clinical Application in Cryobanking. Int. J. Mol. Sci. 2022, 23, 2639. [Google Scholar] [CrossRef]
- Hornberger, K.; Li, R.; Duarte, A.R.C.; Hubel, A. Natural deep eutectic systems for nature-inspired cryopreservation of cells. AIChE J. 2021, 67. [Google Scholar] [CrossRef]
- Taylor, M.J.; Weegman, B.P.; Baicu, S.C.; Giwa, S.E. New Approaches to Cryopreservation of Cells, Tissues, and Organs. Transfus. Med. Hemotherapy 2019, 46, 197–215. [Google Scholar] [CrossRef]
- Wowk, B. How cryoprotectants work. Cryonics 2007, 28, 3–7. [Google Scholar]
- Fahy, G.M.; Wowk, B. Principles of Ice-Free Cryopreservation by Vitrification. Methods Mol. Biol. 2021, 2180, 27–97. [Google Scholar] [PubMed]
- Best, B.P. Cryoprotectant Toxicity: Facts, Issues, and Questions. Rejuvenation Res. 2015, 18, 422–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meryman, H.T. Cryoprotective agents. Cryobiology 1971, 8, 173–183. [Google Scholar] [CrossRef]
- Bachtiger, F.; Congdon, T.R.; Stubbs, C.; Gibson, M.I.; Sosso, G.C. The atomistic details of the ice recrystallisation inhibition activity of PVA. Nat. Commun. 2021, 12, 1323. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Liu, W.; Liu, B.; He, X.; Guo, H.; Xue, S.; Yan, X.; Jaganathan, G.K. Cryopreservation of human T lymphocytes under fast cooling with controlled ice nucleation in cryoprotective solutions of low toxicity. Cryobiology 2021, 103, 92–100. [Google Scholar] [CrossRef]
- Huebinger, J.; Han, H.-M.; Hofnagel, O.; Vetter, I.R.; Bastiaens, P.I.; Grabenbauer, M. Direct Measurement of Water States in Cryopreserved Cells Reveals Tolerance toward Ice Crystallization. Biophys. J. 2016, 110, 840–849. [Google Scholar] [CrossRef] [Green Version]
- Crowe, L.M.; Mouradian, R.; Crowe, J.H.; Jackson, S.A.; Womersley, C. Effects of carbohydrates on membrane stability at low water activities. Biochim. Biophys. Acta Biomembr. 1984, 769, 141–150. [Google Scholar] [CrossRef]
- Spindler, R.; Wolkers, W.; Glasmacher, B. Effect of Me2SO on Membrane Phase Behavior and Protein Denaturation of Human Pulmonary Endothelial Cells Studied by In Situ FTIR Spectroscopy. J. Biomech. Eng. 2009, 131, 074517. [Google Scholar] [CrossRef]
- Oldenhof, H.; Friedel, K.; Akhoondi, M.; Gojowsky, M.; Wolkers, W.; Sieme, H. Membrane phase behavior during cooling of stallion sperm and its correlation with freezability. Mol. Membr. Biol. 2012, 29, 95–106. [Google Scholar] [CrossRef]
- Fahy, G.M.; Lilley, T.H.; Linsdell, H.; Douglas, M.S.; Meryman, H.T. Cryoprotectant toxicity and cryoprotectant toxicity reduction: In search of molecular mechanisms. Cryobiology 1990, 27, 247–268. [Google Scholar] [CrossRef]
- Arakawa, T.; Carpenter, J.F.; Kita, Y.A.; Crowe, J.H. The basis for toxicity of certain cryoprotectants: A hypothesis. Cryobiology 1990, 27, 401–415. [Google Scholar] [CrossRef]
- Dludla, P.V.; Jack, B.; Viraragavan, A.; Pheiffer, C.; Johnson, R.; Louw, J.; Muller, C.J. A dose-dependent effect of dimethyl sulfoxide on lipid content, cell viability and oxidative stress in 3T3-L1 adipocytes. Toxicol. Rep. 2018, 5, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Kopeika, J.; Thornhill, A.; Khalaf, Y. The effect of cryopreservation on the genome of gametes and embryos: Principles of cryobiology and critical appraisal of the evidence. Hum. Reprod. Update 2015, 21, 209–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwatani, M.; Ikegami, K.; Kremenska, Y.; Hattori, N.; Tanaka, S.; Yagi, S.; Shiota, K. Dimethyl Sulfoxide Has an Impact on Epigenetic Profile in Mouse Embryoid Body. Stem Cells 2006, 24, 2549–2556. [Google Scholar] [CrossRef] [PubMed]
- Scirè, A.; Cianfruglia, L.; Minnelli, C.; Bartolini, D.; Torquato, P.; Principato, G.; Galli, F.; Armeni, T. Glutathione compartmentalization and its role in glutathionylation and other regulatory processes of cellular pathways. BioFactors 2019, 45, 152–168. [Google Scholar] [CrossRef]
- Fahy, G.M. Cryoprotectant toxicity neutralization. Cryobiology 2010, 60 (Suppl. 3), S45–S53. [Google Scholar] [CrossRef]
- Fahy, G.M.; Wowk, B.; Wu, J.; Phan, J.; Rasch, C.; Chang, A.; Zendejas, E. Cryopreservation of organs by vitrification: Perspectives and recent advances. Cryobiology 2004, 48, 157–178. [Google Scholar] [CrossRef]
- Griffith, M.; Lumb, C.; Wiseman, S.B.; Wisniewski, M.; Johnson, R.W.; Marangoni, A.G. Antifreeze Proteins Modify the Freezing Process In Planta. Plant Physiol. 2005, 138, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Kuiper, M.J.; Morton, C.; E Abraham, S.; Gray-Weale, A. The biological function of an insect antifreeze protein simulated by molecular dynamics. eLife 2015, 4, e05142. [Google Scholar] [CrossRef]
- Gilfanova, R.; Callegari, A.; Childs, A.; Yang, G.; Luarca, M.; Gutierrez, A.G.; Medina, K.I.; Mai, J.; Hui, A.; Kline, M.; et al. A bioinspired and chemically defined alternative to dimethyl sulfoxide for the cryopreservation of human hematopoietic stem cells. Bone Marrow Transplant. 2021, 56, 2644–2650. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Dooley, D.C. Assessment of Cell Viability and Apoptosis in Human Umbilical Cord Blood Following Storage. J. Hematotherapy 2003, 12, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Storey, K.B.; Storey, J.M. Molecular Biology of Freezing Tolerance. Compr. Physiol. 2013, 3, 1283–1308. [Google Scholar] [PubMed]
- Elliott, G.D.; Liu, X.-H.; Cusick, J.L.; Menze, M.; Vincent, J.; Witt, T.; Hand, S.; Toner, M. Trehalose uptake through P2X7 purinergic channels provides dehydration protection. Cryobiology 2006, 52, 114–127. [Google Scholar] [CrossRef] [Green Version]
- Eroglu, A. Cryopreservation of mammalian oocytes by using sugars: Intra- and extracellular raffinose with small amounts of dimethylsulfoxide yields high cryosurvival, fertilization, and development rates. Cryobiology 2010, 60 (Suppl. 3), S54–S59. [Google Scholar] [CrossRef] [Green Version]
- Petrenko, Y.A.; Rogulska, O.; Mutsenko, V.V.; Petrenko, A.Y. A sugar pretreatment as a new approach to the Me2SO- and xeno-free cryopreservation of human mesenchymal stromal cells. Cryo Lett. 2014, 35, 239–246. [Google Scholar]
- Sola-Penna, M.; Ferreira-Pereira, A.; Lemos, A.D.P.; Meyer-Ferwandes, J.R. Carbohydrate Protection of Enzyme Structure and Function against Guanidinium Chloride Treatment Depends on the Nature of Carbohydrate and Enzyme. JBIC J. Biol. Inorg. Chem. 1997, 248, 24–29. [Google Scholar] [CrossRef]
- Crowe, J.H.; Carpenter, J.F.; Crowe, L.M. The Role of Vitrification in Anhydrobiosis. Annu. Rev. Physiol. 1998, 60, 73–103. [Google Scholar] [CrossRef]
- Jesus, A.R.; Meneses, L.; Duarte, A.R.C.; Paiva, A. Natural deep eutectic systems, an emerging class of cryoprotectant agents. Cryobiology 2021, 101, 95–104. [Google Scholar] [CrossRef]
- Maneffa, A.J.; Harrison, A.B.; Radford, S.J.; Whitehouse, A.S.; Clark, J.H.; Matharu, A.S. Deep Eutectic Solvents Based on Natural Ascorbic Acid Analogues and Choline Chloride. ChemistryOpen 2020, 9, 559–567. [Google Scholar] [CrossRef]
- Qiao, Y.; Cai, H.-L.; Yang, X.; Zang, Y.-Y.; Chen, Z.-G. Effects of natural deep eutectic solvents on lactic acid bacteria viability during cryopreservation. Appl. Microbiol. Biotechnol. 2018, 102, 5695–5705. [Google Scholar] [CrossRef] [PubMed]
- Castro, V.I.; Craveiro, R.; Silva, J.M.; Reis, R.L.; Paiva, A.; Duarte, A.R.C. Natural deep eutectic systems as alternative nontoxic cryoprotective agents. Cryobiology 2018, 83, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Pollock, K.; Yu, G.; Moller-Trane, R.; Koran, M.; Dosa, P.; McKenna, D.H.; Hubel, A. Combinations of Osmolytes, Including Monosaccharides, Disaccharides, and Sugar Alcohols Act in Concert During Cryopreservation to Improve Mesenchymal Stromal Cell Survival. Tissue Eng. Part C Methods 2016, 22, 999–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pi, C.-H.; Yu, G.; Petersen, A.; Hubel, A. Characterizing the “sweet spot” for the preservation of a T-cell line using osmolytes. Sci. Rep. 2018, 8, 16223. [Google Scholar] [CrossRef] [PubMed]
- Knani, L.; Bartolini, D.; Kechiche, S.; Tortoioli, C.; Murdolo, G.; Moretti, M.; Messaoudi, I.; Reiter, R.J.; Galli, F. Melatonin prevents cadmium-induced bone damage: First evidence on an improved osteogenic/adipogenic differentiation balance of mesenchymal stem cells as underlying mechanism. J. Pineal Res. 2019, 67, e12597. [Google Scholar] [CrossRef] [PubMed]
- Siu, A.W.; Maldonado, M.; Hidalgo, M.S.; Tan, D.-X.; Reiter, R.J. Protective effects of melatonin in experimental free radical-related ocular diseases. J. Pineal Res. 2006, 40, 101–109. [Google Scholar] [CrossRef]
- Luchetti, F.; Canonico, B.; Bartolini, D.; Arcangeletti, M.; Ciffolilli, S.; Murdolo, G.; Piroddi, M.; Papa, S.; Reiter, R.J.; Galli, F. Melatonin regulates mesenchymal stem cell differentiation: A review. J. Pineal Res. 2014, 56, 382–397. [Google Scholar] [CrossRef] [Green Version]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef]
- Alvarez-Diduk, R.; Galano, A.; Tan, D.X.; Reiter, R.J. N-Acetylserotonin and 6-Hydroxymelatonin against Oxidative Stress: Implications for the Overall Protection Exerted by Melatonin. J. Phys. Chem. B 2015, 119, 8535–8543. [Google Scholar] [CrossRef] [Green Version]
- Luchetti, F.; Betti, M.; Canonico, B.; Arcangeletti, M.; Ferri, P.; Galli, F.; Papa, S. ERK MAPK activation mediates the antiapoptotic signaling of melatonin in UVB-stressed U937 cells. Free Radic. Biol. Med. 2009, 46, 339–351. [Google Scholar] [CrossRef]
- Han, L.; Wang, H.; Li, L.; Li, X.; Ge, J.; Reiter, R.J.; Wang, Q. Melatonin protects against maternal obesity-associated oxidative stress and meiotic defects in oocytes via the SIRT3-SOD2-dependent pathway. J. Pineal Res. 2017, 63, e12431. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J. Pineal. Res. 2013, 54, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.S.; Han, X.; Sivakumaran, P.; Lim, S.Y.; Morrison, W.A. Melatonin Protects Human Adipose-Derived Stem Cells from Oxidative Stress and Cell Death. Arch. Plast. Surg. 2016, 43, 237–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, A.; García, J.A.; Escames, G.; Venegas, C.; Ortiz, F.; López, L.C.; Acuña-Castroviejo, D. Melatonin protects the mitochondria from oxidative damage reducing oxygen consumption, membrane potential, and superoxide anion production. J. Pineal Res. 2009, 46, 188–198. [Google Scholar] [CrossRef]
- Solanas, E.; Sostres, C.; Serrablo, A.; García-Gil, A.; Aranguren, F.; Jimenez, P.; Serrano, M.T. Incubation with Dimethyl Sulfoxide Prior to Cryopreservation Improves Functionality of Thawed Human Primary Hepatocytes. Biopreservation Biobanking 2012, 10, 446–453. [Google Scholar] [CrossRef]
- Fitzpatrick, E.; Mitry, R.R.; Dhawan, A. Human hepatocyte transplantation: State of the art. J. Intern. Med. 2009, 266, 339–357. [Google Scholar] [CrossRef]
- Solanas, E.; Aranguren, F.J.; Hughes, R.D.; Sostres, C.; Serrablo, A.; García-Gil, A.; García, J.J.; Jiménez, P.; Serrano, M.T. Effect of Dimethyl Sulfoxide and Melatonin on the Isolation of Human Primary Hepatocytes. Cells Tissues Organs 2015, 200, 316–325. [Google Scholar] [CrossRef]
- Youm, H.W.; Lee, J.; Jee, B.C.; Suh, C.S.; Kim, S.H. Optimal vitrification protocol for mouse ovarian tissue cryopreservation: Effect of cryoprotective agents and in vitro culture on vitrified-warmed ovarian tissue survival. Hum. Reprod. 2014, 29, 720–730. [Google Scholar] [CrossRef] [Green Version]
- Rocha, C.D.; Soares, M.M.; Antonino, D.D.C.; Júnior, J.M.; Mohallem, R.F.F.; Rodrigues, A.P.R.; Figueiredo, J.R.; Beletti, M.E.; Jacomini, J.O.; Alves, B.G.; et al. Positive effect of resveratrol against preantral follicles degeneration after ovarian tissue vitrification. Theriogenology 2018, 114, 244–251. [Google Scholar] [CrossRef]
- Liu, X.C.; Sun, T.C.; Li, H.Y.; Si, L.N.; Wei, M.; Chen, Z.H.; Cheng, L.Y.; Yang, S.H. Antioxidative Effect of Melatonin on Cryopreserved Ovarian Tissue in Mice. Cryobiology 2020, 96, 99–105. [Google Scholar] [CrossRef]
- Sun, T.C.; Liu, X.C.; Yang, S.H.; Song, L.L.; Zhou, S.J.; Deng, S.L.; Tian, L.; Cheng, L.Y. Melatonin Inhibits Oxidative Stress and Apoptosis in Cryopreserved Ovarian Tissues via Nrf2/HO-1 Signaling Pathway. Front. Mol. Biosci. 2020, 7, 163. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.; Wang, X.; Xu, Q.; Wu, Y.; Si, X.; Jiang, D. Effect of 3-nitropropionic acid inducing oxidative stress and apoptosis of granulosa cells in geese. Biosci. Rep. 2018, 38, BSR20180274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amidi, F.; Pazhohan, A.; Nashtaei, M.S.; Khodarahmian, M.; Nekoonam, S. The role of antioxidants in sperm freezing: A review. Cell Tissue Bank. 2016, 17, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Lin, Q.; Liu, R.; Xiao, W.; Liu, W. Protective Effects of Ascorbate and Catalase on Human Spermatozoa during Cryopreservation. J. Androl. 2010, 31, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Bahmyari, R.; Zare, M.; Sharma, R.; Agarwal, A.; Halvaei, I. The efficacy of antioxidants in sperm parameters and production of reactive oxygen species levels during the freeze-thaw process: A systematic review and meta-analysis. Andrologia 2020, 52, e13514. [Google Scholar] [CrossRef]
- Najafi, A.; Adutwum, E.; Yari, A.; Salehi, E.; Mikaeili, S.; Dashtestani, F.; Abolhassani, F.; Rashki, L.; Shiasi, S.; Asadi, E. Melatonin affects membrane integrity, intracellular reactive oxygen species, caspase3 activity and AKT phosphorylation in frozen thawed human sperm. Cell Tissue Res. 2018, 372, 149–159. [Google Scholar] [CrossRef]
- López, L.C.; Acuña-Castroviejo, D.; Pino, A.D.; Tejada, M.; Escames, G. Mitochondrial disorders therapy: The utility of melatonin. Open Biol. J. 2010, 3, 53–56. [Google Scholar] [CrossRef]
- Garcia, J.J.; López-Pingarrón, L.; Almeida-Souza, P.; Tres, A.; Escudero, P.; García-Gil, F.A.; Tan, D.-X.; Reiter, R.J.; Ramírez, J.M.; Bernal-Pérez, M. Protective effects of melatonin in reducing oxidative stress and in preserving the fluidity of biological membranes: A review. J. Pineal Res. 2014, 56, 225–237. [Google Scholar] [CrossRef]
- Alevra, A.I.; Exadactylos, A.; Mente, E.; Papadopoulos, S. The Protective Role of Melatonin in Sperm Cryopreservation of Farm Animals and Human: Lessons for Male Fish Cryopreservation. Animals 2022, 12, 791. [Google Scholar] [CrossRef]
- Divar, M.R.; Azari, M.; Mogheiseh, A.; Ghahramani, S. Supplementation of melatonin to cooling and freezing extenders improves canine spermatozoa quality measures. BMC Vet. Res. 2022, 18, 86. [Google Scholar] [CrossRef]
- Chen, L.-Y.; Willis, W.D.; Eddy, E.M. Targeting the Gdnf Gene in peritubular myoid cells disrupts undifferentiated spermatogonial cell development. Proc. Natl. Acad. Sci. USA 2016, 113, 1829–1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savvulidi, F.; Ptacek, M.; Vargova, K.S.; Stadnik, L. Manipulation of spermatogonial stem cells in livestock species. J. Anim. Sci. Biotechnol. 2019, 10, 46. [Google Scholar] [CrossRef] [PubMed]
- ZZhou, H.; Zeng, Z.; Koentgen, F.; Khan, M.; Mombaerts, P. The testicular soma of Tsc22d3 knockout mice supports spermatogenesis and germline transmission from spermatogonial stem cell lines upon transplantation. Genesis 2019, 57, e23295. [Google Scholar]
- Takashima, S.; Shinohara, T. Culture and transplantation of spermatogonial stem cells. Stem Cell Res. 2018, 29, 46–55. [Google Scholar] [CrossRef]
- Kim, K.-J.; Lee, Y.-A.; Kim, B.-J.; Kim, Y.-H.; Kim, B.-G.; Kang, H.-G.; Jung, S.-E.; Choi, S.-H.; Schmidt, J.A.; Ryu, B.-Y. Cryopreservation of putative pre-pubertal bovine spermatogonial stem cells by slow freezing. Cryobiology 2015, 70, 175–183. [Google Scholar] [CrossRef]
- Feng, T.-Y.; Li, Q.; Ren, F.; Xi, H.-M.; Lv, D.-L.; Li, Y.; Hu, J.-H. Melatonin Protects Goat Spermatogonial Stem Cells against Oxidative Damage during Cryopreservation by Improving Antioxidant Capacity and Inhibiting Mitochondrial Apoptosis Pathway. Oxidative Med. Cell. Longev. 2020, 2020, 5954635. [Google Scholar] [CrossRef]
- Navid, S.; Rastegar, T.; Baazm, M.; Alizadeh, R.; Talebi, A.; Gholami, K.; Khosravi-Farsani, S.; Koruji, M.; Abbasi, M. In vitro effects of melatonin on colonization of neonate mouse spermatogonial stem cells. Syst. Biol. Reprod. Med. 2017, 63, 370–381. [Google Scholar] [CrossRef] [Green Version]
- McBride, J.A.; Lipshultz, L.I. Male Fertility Preservation. Curr. Urol. Rep. 2018, 19, 49. [Google Scholar] [CrossRef]
- de Vries, R.J.; Banik, P.D.; Nagpal, S.; Weng, L.; Ozer, S.; van Gulik, T.M.; Toner, M.; Tessier, S.N.; Uygun, K. Bulk Droplet Vitrification: An Approach to Improve Large-Scale Hepatocyte Cryopreservation Outcome. Langmuir 2019, 35, 7354–7363. [Google Scholar] [CrossRef]
- Cypser, J.R.; Chick, W.S.; Fahy, G.M.; Schumacher, G.; Johnson, T.E. Genetic suppression of cryoprotectant toxicity. Cryobiology 2019, 86, 95–102. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CPA | Source and Examples | Classification | Reference |
|---|---|---|---|
| Sulfoxides | Synthesized from dimethyl sulfide (the prototypal CPA of this category is dimethyl sulfoxide) | Penetrating | [17] |
| Diols | Propylene glycol (or 1,2-propanediol) is the prototypical agent of this category of CPA that is produced via hydration of propylene oxide; other diols are methylene glycol and ethylene glycol | Penetrating | [17] |
| Synthetic and semisynthetic polymeric agents | Synthetic resins obtained from hydrolysis of polyvinyl acetate (such as polyvinyl alcohols), and copolymers and semisynthetic polymers of different origin, including polyampholytes (such as carboxylated poly-ʟ-lysine and others) | Nonpenetrating | [18,19,20,21] |
| Saccharides | Fungi, plants, invertebrate animals (examples are: sucrose and trehalose) | Nonpenetrating | [22,23] |
| Proteins | Animals (antifreeze proteins) | Nonpenetrating | [24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcantonini, G.; Bartolini, D.; Zatini, L.; Costa, S.; Passerini, M.; Rende, M.; Luca, G.; Basta, G.; Murdolo, G.; Calafiore, R.; et al. Natural Cryoprotective and Cytoprotective Agents in Cryopreservation: A Focus on Melatonin. Molecules 2022, 27, 3254. https://doi.org/10.3390/molecules27103254
Marcantonini G, Bartolini D, Zatini L, Costa S, Passerini M, Rende M, Luca G, Basta G, Murdolo G, Calafiore R, et al. Natural Cryoprotective and Cytoprotective Agents in Cryopreservation: A Focus on Melatonin. Molecules. 2022; 27(10):3254. https://doi.org/10.3390/molecules27103254
Chicago/Turabian StyleMarcantonini, Giada, Desirée Bartolini, Linda Zatini, Stefania Costa, Massimiliano Passerini, Mario Rende, Giovanni Luca, Giuseppe Basta, Giuseppe Murdolo, Riccardo Calafiore, and et al. 2022. "Natural Cryoprotective and Cytoprotective Agents in Cryopreservation: A Focus on Melatonin" Molecules 27, no. 10: 3254. https://doi.org/10.3390/molecules27103254
APA StyleMarcantonini, G., Bartolini, D., Zatini, L., Costa, S., Passerini, M., Rende, M., Luca, G., Basta, G., Murdolo, G., Calafiore, R., & Galli, F. (2022). Natural Cryoprotective and Cytoprotective Agents in Cryopreservation: A Focus on Melatonin. Molecules, 27(10), 3254. https://doi.org/10.3390/molecules27103254