
The Attenuating Effect of Beta-Carotene on Streptozotocin Induced Diabetic Vascular Dementia Symptoms in Rats


Abstract
:1. Introduction
2. Results
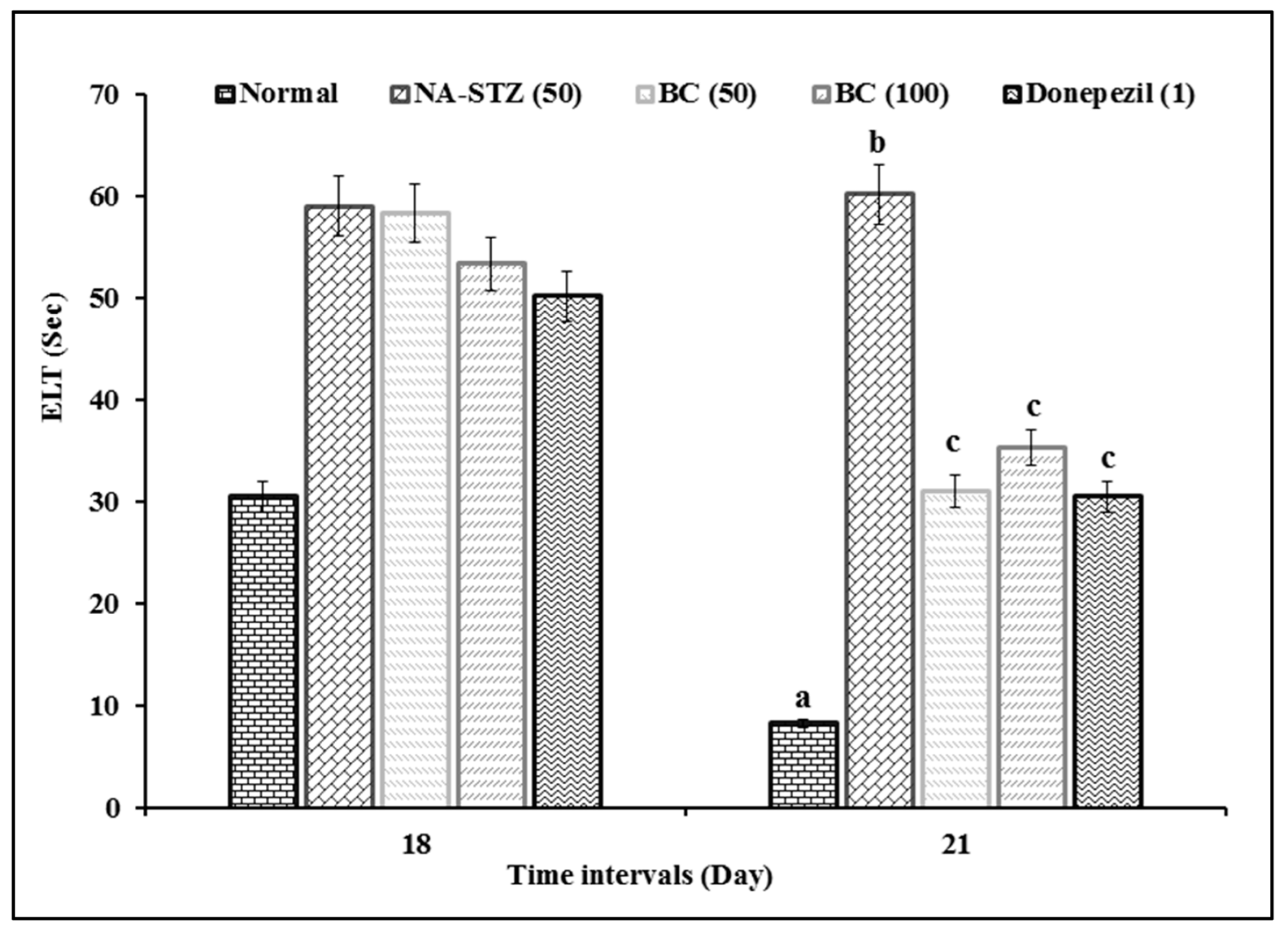
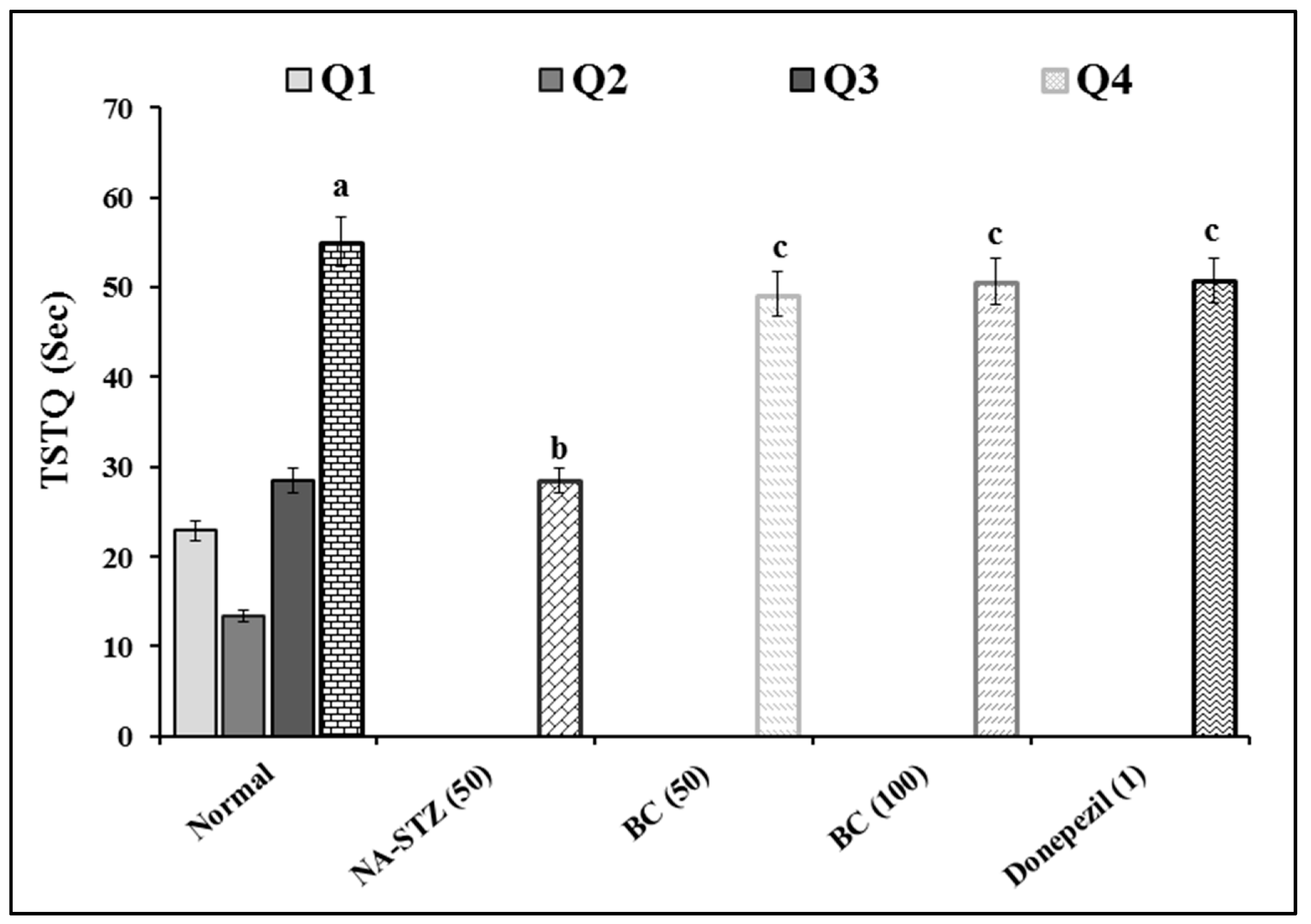
2.1. Effect of Beta Carotene on Escape Latency Time and Time Spent in Target Quadrant in the Morris Water Maze Test
2.2. Effect of Beta Carotene on Tissue Biomarkers
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Induction of Diabetic Vascular Dementia
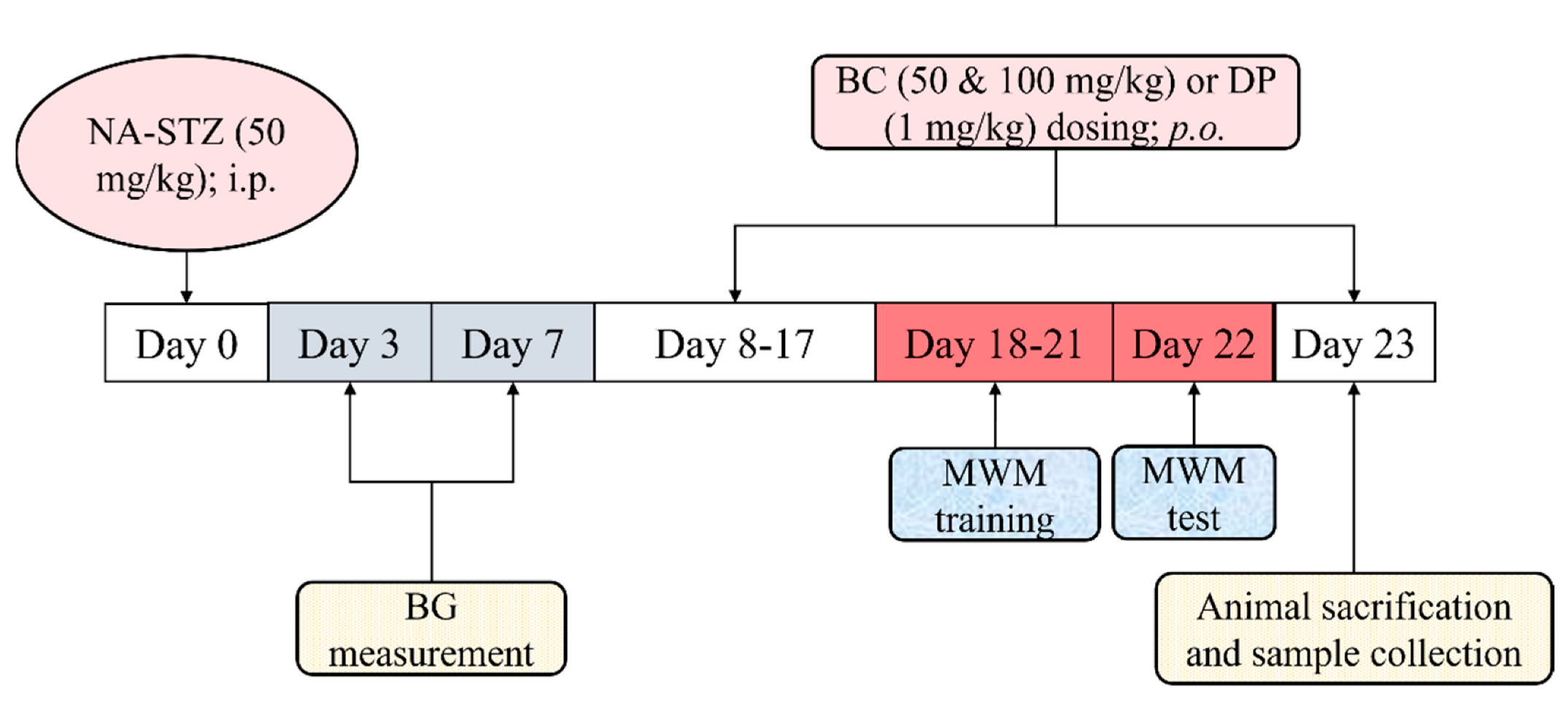
4.3. Experimental Protocol
4.4. Collection of Biological Samples
4.5. Assessment of Spatial Learning and Memory by Morris Water Maze
4.6. Estimation of Acetylcholinesterase (AChE) Activity
4.7. Estimation of Reduced Glutathione (GSH)
4.8. Estimation of Thiobarbituric Acid Reactive Substances (TBARS)
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Calabrese, V.; Giordano, J.; Signorile, A.; Laura Ontario, M.; Castorina, S.; De Pasquale, C.; Eckert, G.; Calabrese, E.J. Major Pathogenic Mechanisms in Vascular Dementia: Roles of Cellular Stress Response and Hormesis in Neuroprotection. J. Neurosci. Res. 2016, 94, 1588–1603. [Google Scholar] [CrossRef] [PubMed]
- Vandepitte, S.; Van Wilder, L.; Putman, K.; Van Den Noortgate, N.; Verhaeghe, S.; Trybou, J.; Annemans, L. Factors Associated with Costs of Care in Community-Dwelling Persons with Dementia from a Third Party Payer and Societal Perspective: A Cross-Sectional Study. BMC Geriatr. 2020, 20, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaria, R.N. The Pathology and Pathophysiology of Vascular Dementia. Neuropharmacology 2018, 134, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Iadecola, C. The Pathobiology of Vascular Dementia. Neuron 2013, 80, 844–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.T.; Thomas, A. Vascular Dementia. Lancet 2015, 386, 1698–1706. [Google Scholar] [CrossRef] [Green Version]
- Venkat, P.; Chopp, M.; Chen, J. Models and Mechanisms of Vascular Dementia. Exp. Neurol. 2015, 272, 97–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.P.; Corriveau, R.A.; Wilcock, D.M. Vascular Contributions to Cognitive Impairment and Dementia (VCID). Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2016, 1862, 857–859. [Google Scholar] [CrossRef]
- Strobel, M.; Tinz, J.; Biesalski, H.-K. The Importance of β-Carotene as a Source of Vitamin A with Special Regard to Pregnant and Breastfeeding Women. Eur. J. Nutr. 2007, 46, 1–20. [Google Scholar] [CrossRef]
- Gong, X.; Marisiddaiah, R.; Rubin, L.P. Inhibition of Pulmonary β-Carotene 15, 15’-Oxygenase Expression by Glucocorticoid Involves PPARα. PLoS ONE 2017, 12, e0181466. [Google Scholar] [CrossRef] [Green Version]
- Kawata, A.; Murakami, Y.; Suzuki, S.; Fujisawa, S. Anti-Inflammatory Activity of β-Carotene, Lycopene and Tri-n-Butylborane, a Scavenger of Reactive Oxygen Species. In Vivo 2018, 32, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.O.; Kim, M.-H.; Kim, H. β-Carotene Inhibits Activation of NF-ΚB, Activator Protein-1, and STAT3 and Regulates Abnormal Expression of Some Adipokines in 3T3-L1 Adipocytes. J. Cancer Prev. 2018, 23, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, M.; Reddy, P.H. Stroke, Vascular Dementia, and Alzheimer’s Disease: Molecular Links. J. Alzheimer’s Dis. 2016, 54, 427–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Liu, M.; Liu, J.; Zhang, T.; Zhang, L.; Li, H.; Luo, Z. ShenmaYizhi Decoction Improves the Mitochondrial Structure in the Brain and Ameliorates Cognitive Impairment in VCI Rats via the AMPK/UCP2 Signaling Pathway. Neuropsychiatr. Dis. Treat. 2021, 17, 1937–1951. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Cao, Y.; Ma, L.; Pei, H.; Rausch, W.D.; Li, H. Dysfunction of Cerebrovascular Endothelial Cells: Prelude to Vascular Dementia. Front. Aging Neurosci. 2018, 10, 376. [Google Scholar] [CrossRef] [Green Version]
- Hussain, B.; Fang, C.; Chang, J. Blood–Brain Barrier Breakdown: An Emerging Biomarker of Cognitive Impairment in Normal Aging and Dementia. Front. Neurosci. 2021, 15, 688090. [Google Scholar] [CrossRef]
- Venkat, P.; Chopp, M.; Zacharek, A.; Cui, C.; Zhang, L.; Li, Q.; Lu, M.; Zhang, T.; Liu, A.; Chen, J. White Matter Damage and Glymphatic Dysfunction in a Model of Vascular Dementia in Rats with No Prior Vascular Pathologies. Neurobiol. Aging 2017, 50, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Grande, G.; Qiu, C.; Fratiglioni, L. Prevention of Dementia in an Ageing World: Evidence and Biological Rationale. Ageing Res. Rev. 2020, 64, 101045. [Google Scholar] [CrossRef]
- Ballinger, E.C.; Ananth, M.; Talmage, D.A.; Role, L.W. Basal Forebrain Cholinergic Circuits and Signaling in Cognition and Cognitive Decline. Neuron 2016, 91, 1199–1218. [Google Scholar] [CrossRef] [Green Version]
- Bohnen, N.I.; Grothe, M.J.; Ray, N.J.; Müller, M.L.T.M.; Teipel, S.J. Recent Advances in Cholinergic Imaging and Cognitive Decline—Revisiting the Cholinergic Hypothesis of Dementia. Curr. Geri. Rep. 2018, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Swardfager, W.; Black, S.E. Pathophysiology of Vascular Cognitive Impairment (I): Theoretical Background. In Stroke Revisited: Vascular Cognitive Impairment; Stroke Revisited; Lee, S.-H., Lim, J.-S., Eds.; Springer: Singapore, 2020; pp. 71–86. ISBN 978-981-10-1432-1. [Google Scholar]
- Jellinger, K.A. Pathology and Pathogenesis of Vascular Cognitive Impairment—A Critical Update. Front. Aging Neurosci. 2013, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kang, H.S.; Kim, H.J.; Moon, Y.; Ryu, H.J.; Kim, M.Y.; Han, S.-H. The Effect of Ischemic Cholinergic Damage on Cognition in Patients With Subcortical Vascular Cognitive Impairment. J. Geriatr. Psychiatry Neurol. 2012, 25, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhu, Z.; Teipel, S.J.; Yang, J.; Xing, Y.; Tang, Y.; Jia, J. White Matter Damage in the Cholinergic System Contributes to Cognitive Impairment in Subcortical Vascular Cognitive Impairment, No Dementia. Front. Aging Neurosci. 2017, 9, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohnen, N.I.; Bogan, C.W.; Müller, M.L.T.M. Frontal and Periventricular Brain White Matter Lesions and Cortical Deafferentation of Cholinergic and Other Neuromodulatory Axonal Projections. Eur. Neurol. J. 2009, 1, 33–50. [Google Scholar] [PubMed]
- Marzoughi, S.; Banerjee, A.; Jutzeler, C.R.; Prado, M.A.M.; Rosner, J.; Cragg, J.J.; Cashman, N. Tardive Neurotoxicity of Anticholinergic Drugs: A Review. J. Neurochem. 2021, 158, 1334–1344. [Google Scholar] [CrossRef] [PubMed]
- Benisty, S. Current Concepts in Vascular Dementia. Gériatrie Psychol. Neuropsychiatr. Viellissement 2013, 11, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Ishrat, T.; Khan, M.B.; Hoda, M.N.; Yousuf, S.; Ahmad, M.; Ansari, M.A.; Ahmad, A.S.; Islam, F. Coenzyme Q10 Modulates Cognitive Impairment against Intracerebroventricular Injection of Streptozotocin in Rats. Behav. Brain Res. 2006, 171, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Karunanayake, E.H.; Baker, J.R.J.; Christian, R.A.; Hearse, D.J.; Mellows, G. Autoradiographic Study of the Distribution and Cellular Uptake of (14C)-Streptozotocin in the Rat. Diabetologia 1976, 12, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.Y.; Song, J.H.; Shin, Y.K.; Han, E.S.; Lee, C.S. Protective Effect of Boldine on Oxidative Mitochondrial Damage in Streptozotocin-Induced Diabetic Rats. Pharmacol. Res. 2000, 42, 361–371. [Google Scholar] [CrossRef]
- Elango, B.; Dornadula, S.; Paulmurugan, R.; Ramkumar, K.M. Pterostilbene Ameliorates Streptozotocin-Induced Diabetes through Enhancing Antioxidant Signaling Pathways Mediated by Nrf2. Chem. Res. Toxicol. 2016, 29, 47–57. [Google Scholar] [CrossRef]
- Nahdi, A.M.T.A.; John, A.; Raza, H. Elucidation of Molecular Mechanisms of Streptozotocin-Induced Oxidative Stress, Apoptosis, and Mitochondrial Dysfunction in Rin-5F Pancreatic β-Cells. Oxidative Med. Cell. Longev. 2017, 2017, 7054272. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, D. B Vitamins and the Brain: Mechanisms, Dose and Efficacy—A Review. Nutrients 2016, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirza, R.; Sharma, B. Beneficial Effects of Pioglitazone, a Selective Peroxisome Proliferator-activated Receptor-γ Agonist in Prenatal Valproic Acid-induced Behavioral and Biochemical Autistic like Features in Wistar Rats. Int. J. Dev. Neurosci. 2019, 76, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Muriach, M.; Flores-Bellver, M.; Romero, F.J.; Barcia, J.M. Diabetes and the Brain: Oxidative Stress, Inflammation, and Autophagy. Oxidative Med. Cell. Longev. 2014, 2014, 102158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houlton, J.; Barwick, D.; Clarkson, A.N. Frontal Cortex Stroke-Induced Impairment in Spatial Working Memory on the Trial-Unique Nonmatching-to-Location Task in Mice. Neurobiol. Learn. Mem. 2021, 177, 107355. [Google Scholar] [CrossRef]
- Tayler, H.; Miners, J.S.; Güzel, Ö.; MacLachlan, R.; Love, S. Mediators of Cerebral Hypoperfusion and Blood-brain Barrier Leakiness in Alzheimer’s Disease, Vascular Dementia and Mixed Dementia. Brain Pathol. 2021, 31, e12935. [Google Scholar] [CrossRef]
- Zhou, T.; Liu, H.-J.; Xu, P. Effect of beta-carotene on learning, memory and expression of caspase-3 and phosphorylated tau in hippocampus of rats with obstructive sleep apnea syndrome. J. Clin. Neurol. 2019, 6, 50–53. [Google Scholar]
- Hira, S.; Saleem, U.; Anwar, F.; Sohail, M.F.; Raza, Z.; Ahmad, B. β-Carotene: A Natural Compound Improves Cognitive Impairment and Oxidative Stress in a Mouse Model of Streptozotocin-Induced Alzheimer’s Disease. Biomolecules 2019, 9, 441. [Google Scholar] [CrossRef] [Green Version]
- Darvesh, S.; Hopkins, D.A.; Geula, C. Neurobiology of Butyrylcholinesterase. Nat. Rev. Neurosci. 2003, 4, 131–138. [Google Scholar] [CrossRef]
- Ha, Z.Y.; Mathew, S.; Yeong, K.Y. Butyrylcholinesterase: A Multifaceted Pharmacological Target and Tool. CPPS 2020, 21, 99–109. [Google Scholar] [CrossRef]
- Singh, P.; Gupta, S.; Sharma, B. Antagonism of Endothelin (ETA and ETB) Receptors During Renovascular Hypertension-Induced Vascular Dementia Improves Cognition. CNR 2016, 13, 219–229. [Google Scholar] [CrossRef]
- Sodhi, R.K.; Singh, N. All-Trans Retinoic Acid Rescues Memory Deficits and Neuropathological Changes in Mouse Model of Streptozotocin-Induced Dementia of Alzheimer’s Type. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 40, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (US). Panel on Dietary Antioxidants and Related Compound. In β-Carotene and Other Carotenoids; National Academies Press (US): Washington, DC, USA, 2000. [Google Scholar]
- Heymann, T.; Heinz, P.; Glomb, M.A. Lycopene Inhibits the Isomerization of β-Carotene during Quenching of Singlet Oxygen and Free Radicals. J. Agric. Food Chem. 2015, 63, 3279–3287. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Kim, Y.J.; Lim, Y.; Oh, B.; Kim, J.Y.; Bouwman, J.; Kwon, O. Combination of Diet Quality Score, Plasma Carotenoids, and Lipid Peroxidation to Monitor Oxidative Stress. Oxidative Med. Cell. Longev. 2018, 2018, 8601028. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Li, L.; Gao, Y.; Xie, Z.; Zhang, Y.; Pan, Z.; Tu, Y.; Wang, H.; Han, Q.; Hu, X.; et al. β-Carotene Provides Neuro Protection after Experimental Traumatic Brain Injury via the Nrf2-ARE Pathway. J. Integr. Neurosci. 2019, 18, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, P.; Unni, S.; Krishnappa, G.; Padmanabhan, B. The Keap1-Nrf2 Pathway: Promising Therapeutic Target to Counteract ROS-Mediated Damage in Cancers and Neurodegenerative Diseases. Biophys. Rev. 2017, 9, 41–56. [Google Scholar] [CrossRef]
- Ghasemi, A.; Khalifi, S.; Jedi, S. Streptozotocin-Nicotinamide-Induced Rat Model of Type 2 Diabetes (Review). Acta Physiol. Hung. 2014, 101, 408–420. [Google Scholar] [CrossRef]
- Masiello, P.; Broca, C.; Gross, R.; Roye, M.; Manteghetti, M.; Hillaire-Buys, D.; Novelli, M.; Ribes, G. Experimental NIDDM: Development of a New Model in Adult Rats Administered Streptozotocin and Nicotinamide. Diabetes 1998, 47, 224–229. [Google Scholar] [CrossRef]
- Birgani, G.A.; Ahangarpour, A.; Khorsandi, L.; Moghaddam, H.F. Anti-Diabetic Effect of Betulinic Acid on Streptozotocin-Nicotinamide Induced Diabetic Male Mouse Model. Braz. J. Pharm. Sci. 2018, 54, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bagle, S.; Muke, S.; Saha, S.; Jayakodi, S. Evaluation of Novel and Superior Formulation CaroTexTM Developed by Biofusion Technology. IJRPS 2019, 10, 1868–1873. [Google Scholar] [CrossRef]
- Spijker, S. Dissection of Rodent Brain Regions. In Neuroproteomics; Li, K.W., Ed.; Neuromethods; Humana Press: Totowa, NJ, USA, 2011; Volume 57, pp. 13–26. ISBN 978-1-61779-110-9. [Google Scholar]
- Bhatia, P.; Singh, N. Tadalafil Ameliorates Memory Deficits, Oxidative Stress, Endothelial Dysfunction and Neuropathological Changes in Rat Model of Hyperhomocysteinemia Induced Vascular Dementia. Int. J. Neurosci. 2020, 1–13. [Google Scholar] [CrossRef]
- Morris, R. Developments of a Water-Maze Procedure for Studying Spatial Learning in the Rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Singh, M.; Prakash, A. Possible Role of Endothelin Receptor against Hyperhomocysteinemia and β-Amyloid Induced AD Type of Vascular Dementia in Rats. Brain Res. Bull. 2017, 133, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved Method for the Determination of Blood Glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal Lipid Peroxidation. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1978; Volume 52, pp. 302–310. ISBN 978-0-12-181952-1. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Groups | ACHE (μmol of Acetylthiocholine Iodide/min/mg of Protein) | GSH (μmol/L/mg of Protein) | TBARS (μM/mg of Protein) |
|---|---|---|---|
| Normal | 159.73 ± 29.05 | 35.68 ± 3.47 | 1.05 ± 0.16 |
| NA-STZ | 311.21 ± 26.58 a | 23.53 ± 2.54 a | 1.67 ± 0.12 a |
| BC (50) | 227.11 ± 18.82 a,b | 31.41 ± 2.45 b | 1.18 ± 0.16 b |
| BC (100) | 234.56 ± 20.97 a,b | 33.51 ± 2.36 b | 1.05 ± 0.22 b |
| DP (1) | 231.29 ± 22.60 a,b | 35.88 ± 1.10 b | 1.01 ± 0.17 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, K.G.; Varatharajan, R.; Muthuraman, A. The Attenuating Effect of Beta-Carotene on Streptozotocin Induced Diabetic Vascular Dementia Symptoms in Rats. Molecules 2022, 27, 4293. https://doi.org/10.3390/molecules27134293
Lim KG, Varatharajan R, Muthuraman A. The Attenuating Effect of Beta-Carotene on Streptozotocin Induced Diabetic Vascular Dementia Symptoms in Rats. Molecules. 2022; 27(13):4293. https://doi.org/10.3390/molecules27134293
Chicago/Turabian StyleLim, Khian Giap, Rajavel Varatharajan, and Arunachalam Muthuraman. 2022. "The Attenuating Effect of Beta-Carotene on Streptozotocin Induced Diabetic Vascular Dementia Symptoms in Rats" Molecules 27, no. 13: 4293. https://doi.org/10.3390/molecules27134293
APA StyleLim, K. G., Varatharajan, R., & Muthuraman, A. (2022). The Attenuating Effect of Beta-Carotene on Streptozotocin Induced Diabetic Vascular Dementia Symptoms in Rats. Molecules, 27(13), 4293. https://doi.org/10.3390/molecules27134293