
The Effect of Copper Salts on Bioactive Compounds and Ultrastructure of Wheat Plants

 , , and
, , and
Abstract
:1. Introduction
2. Results
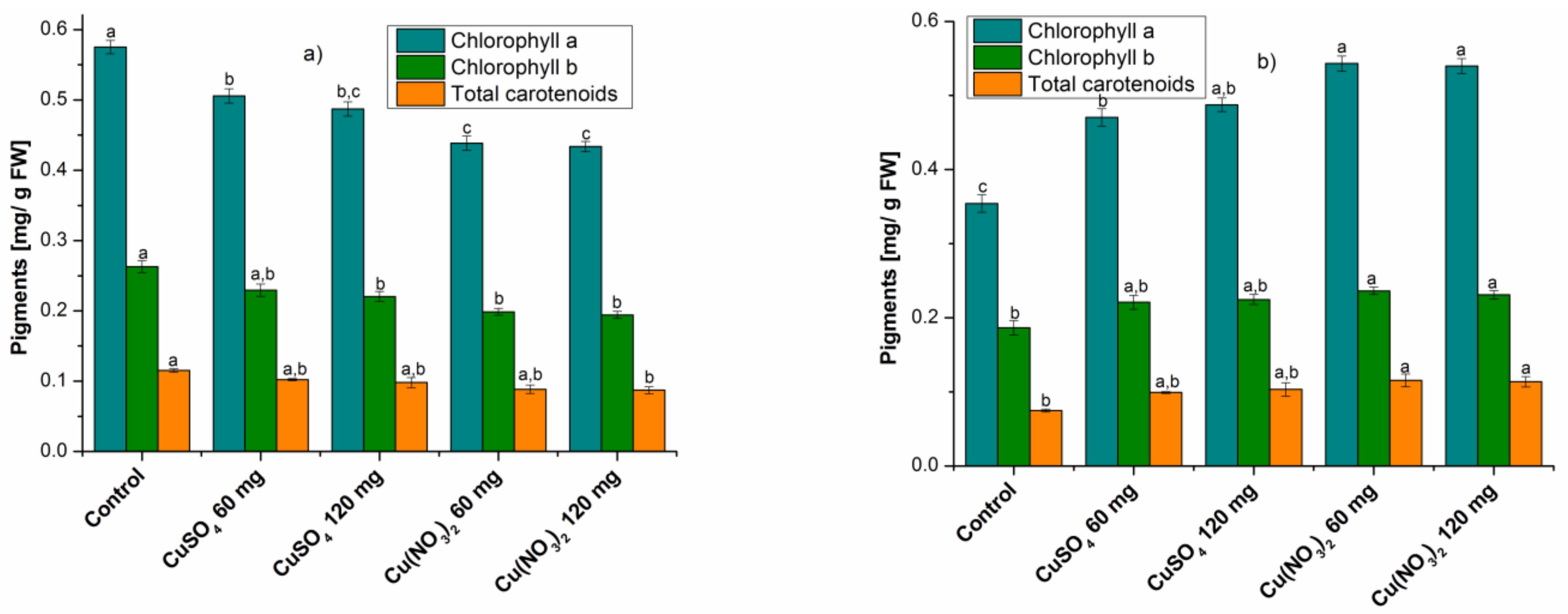
2.1. Analysis of Studied Bioactive Compounds
2.1.1. Determination of Assimilating Pigments
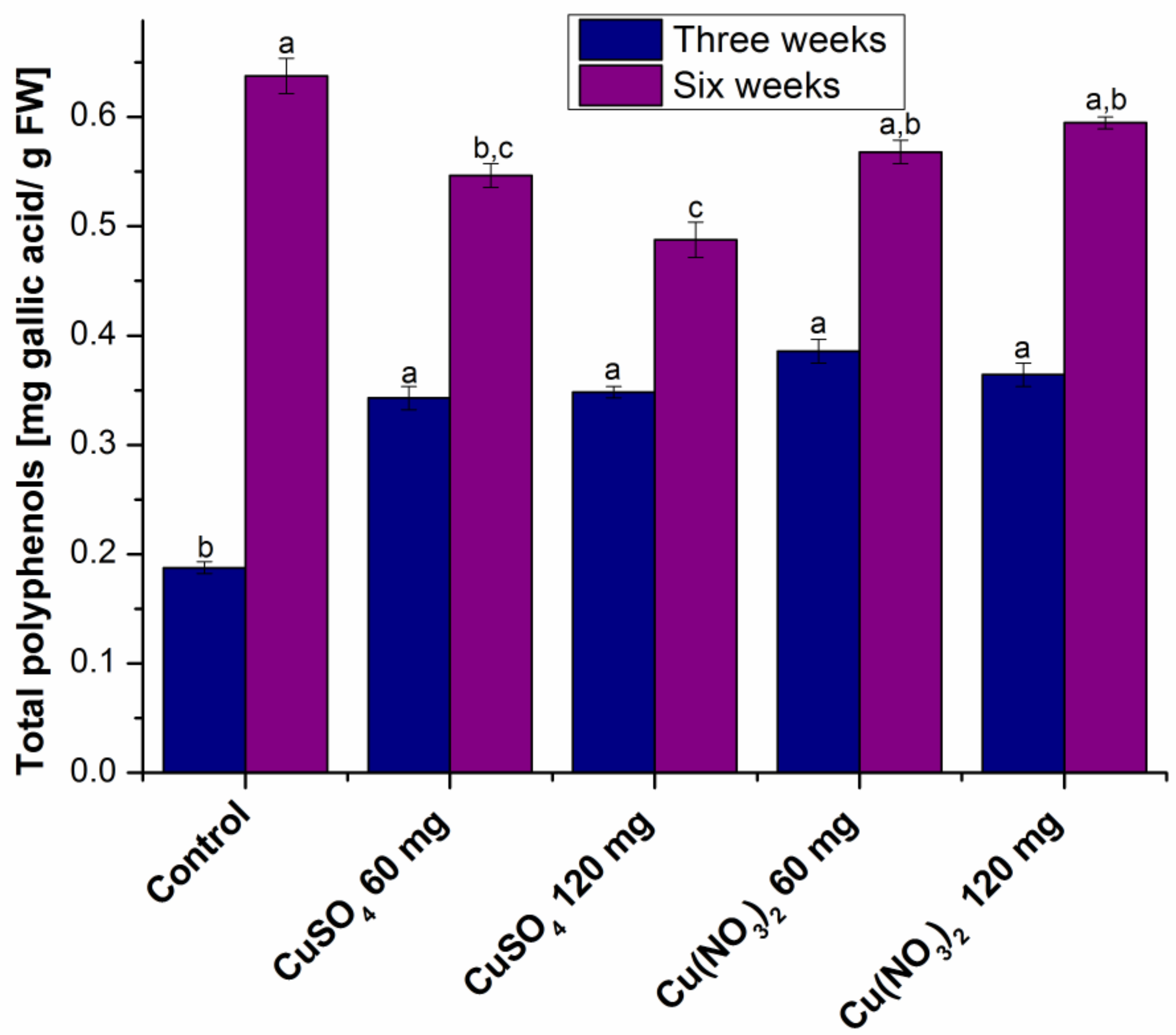
2.1.2. Determination of Total Phenolic Content
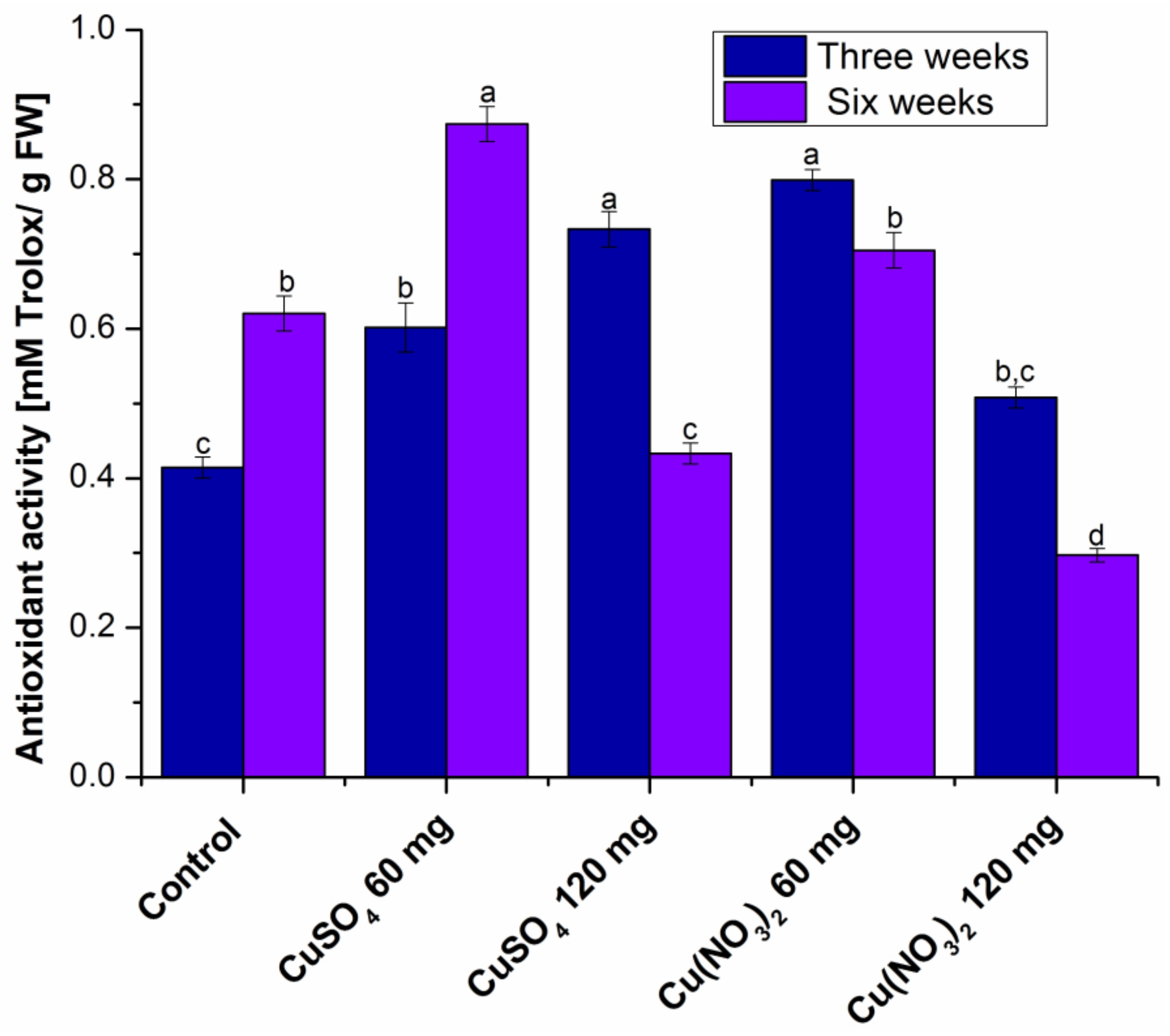
2.1.3. Determination of Total Antioxidant Capacity
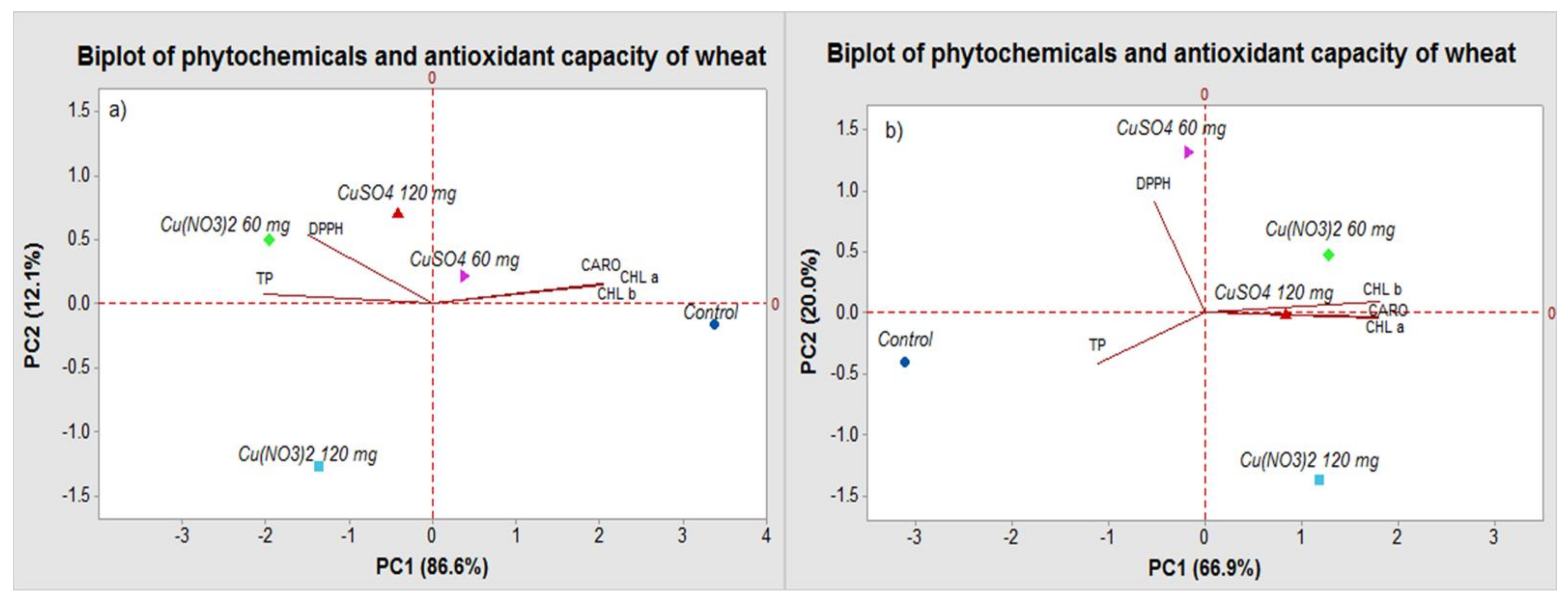
2.2. PCA Analysis
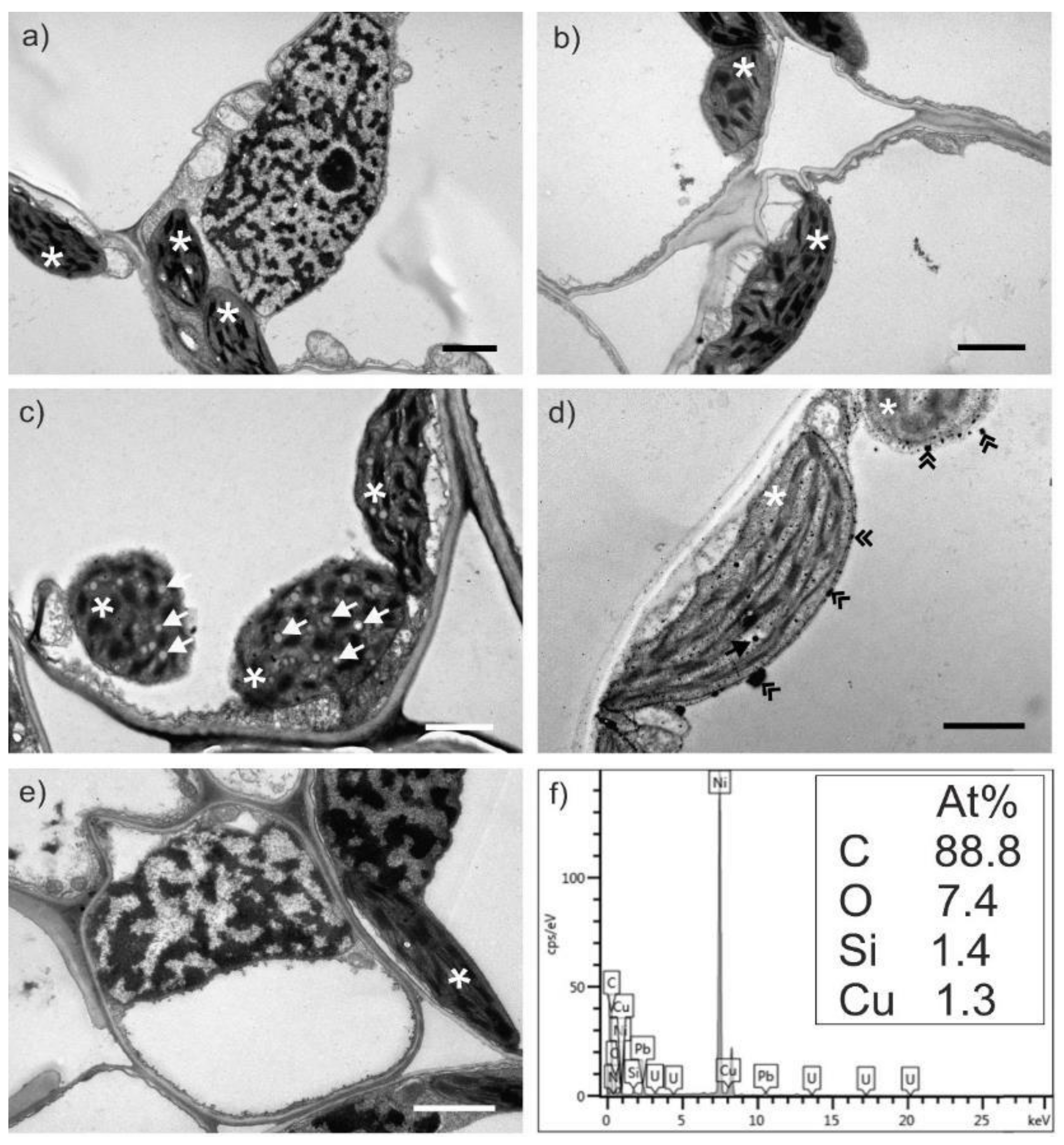
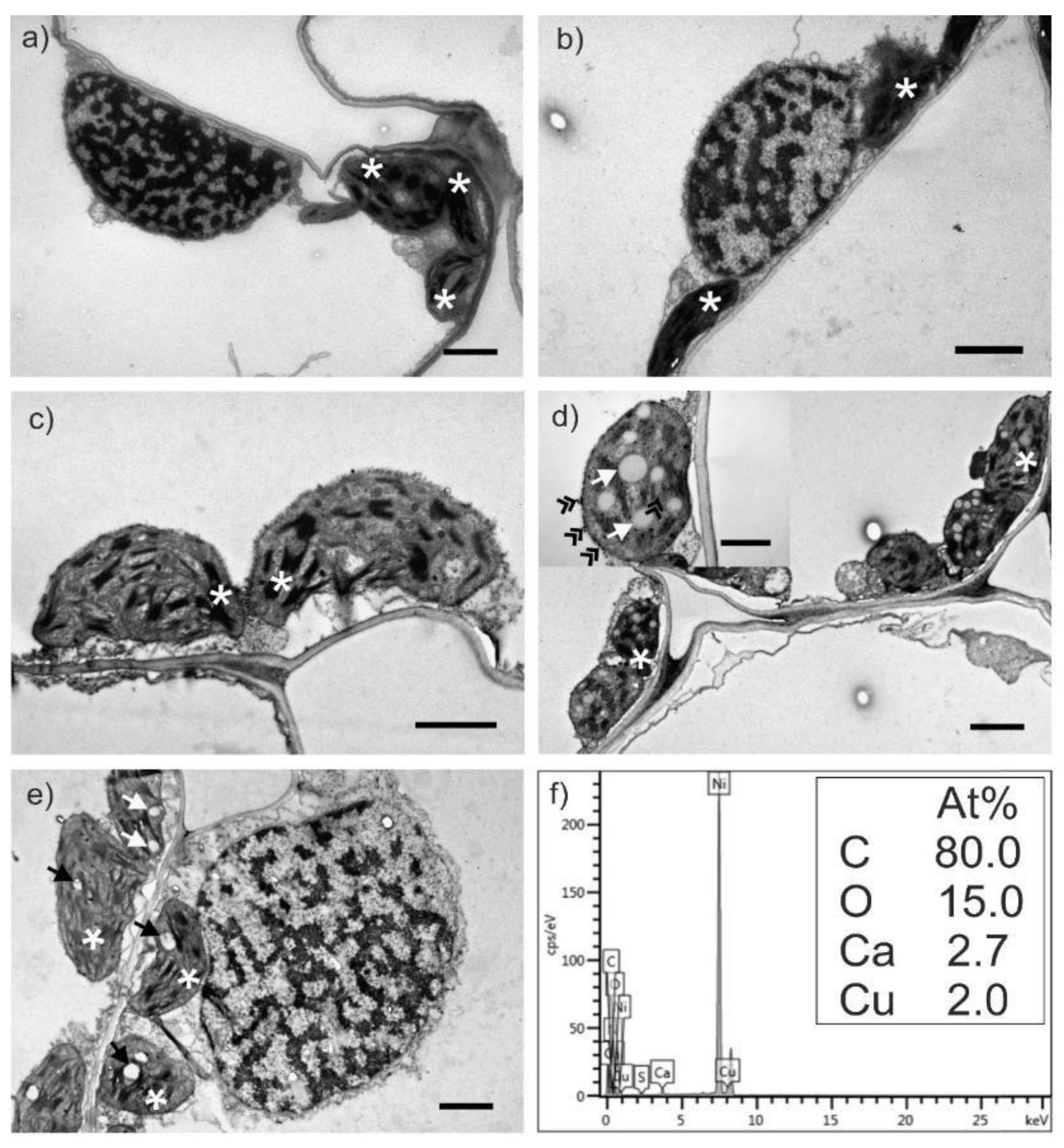
2.3. Transmission Electron Microscopy Analysis of Wheat Leaves
2.4. Effect of Cu on Distribution of Other Elements in Wheat
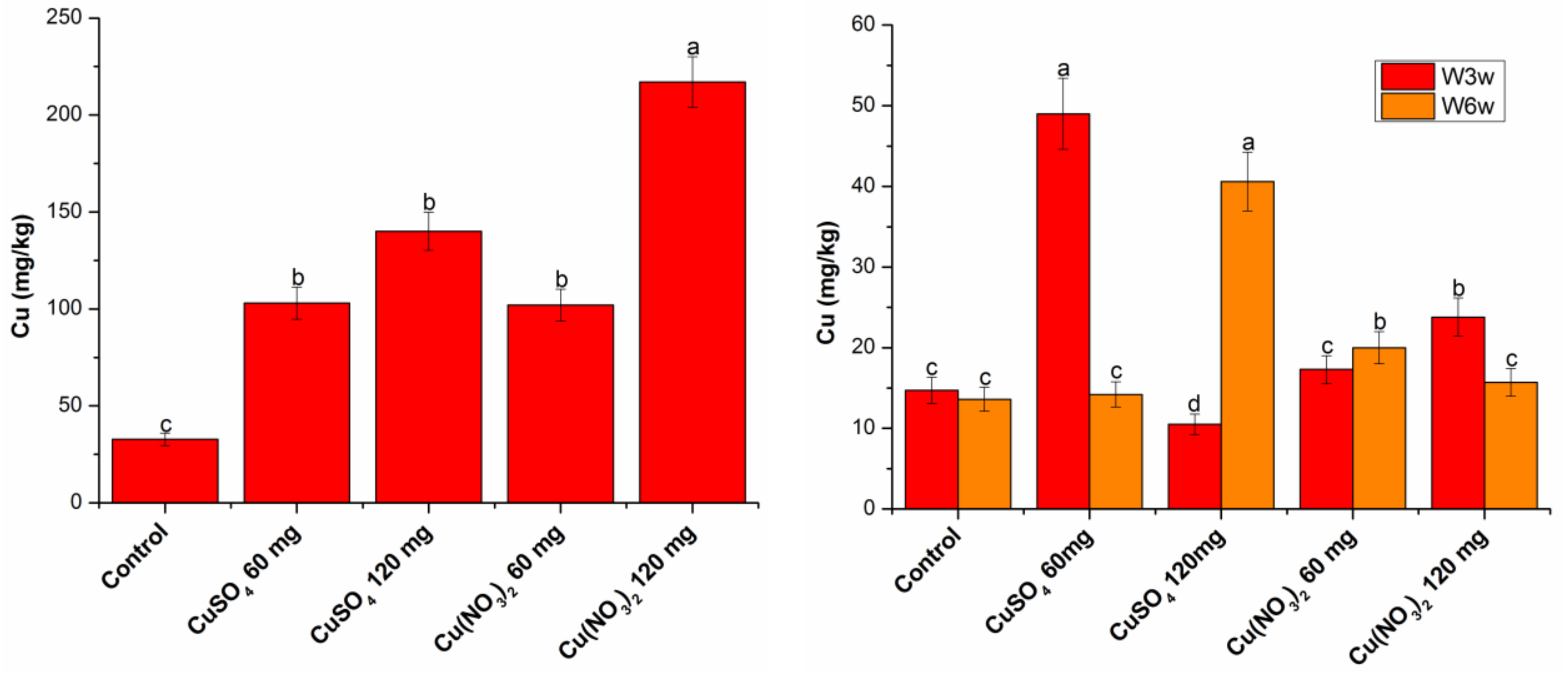
2.5. Cu Content in Wheat
2.6. Bioaccumulation Factor and Translocation Factor
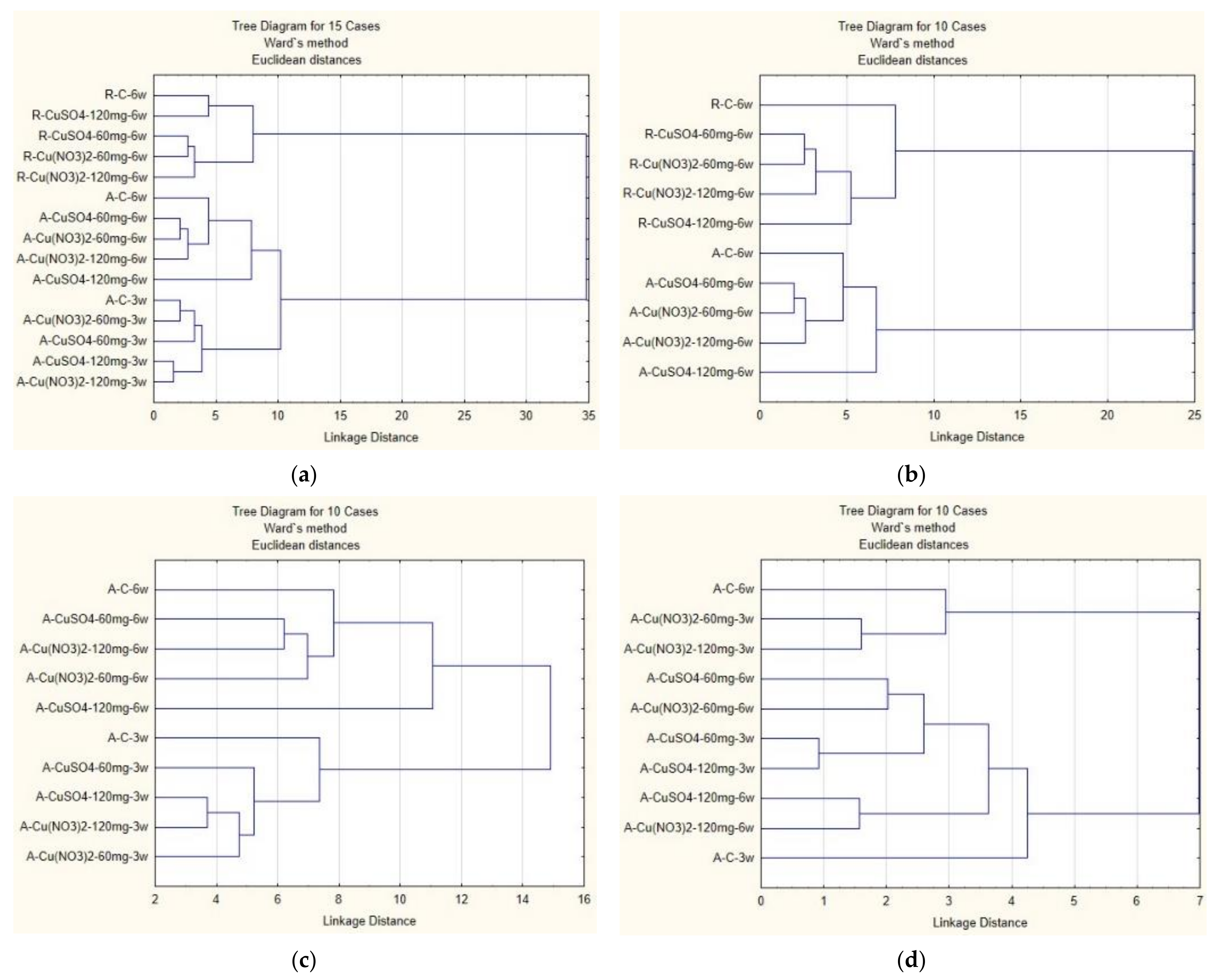
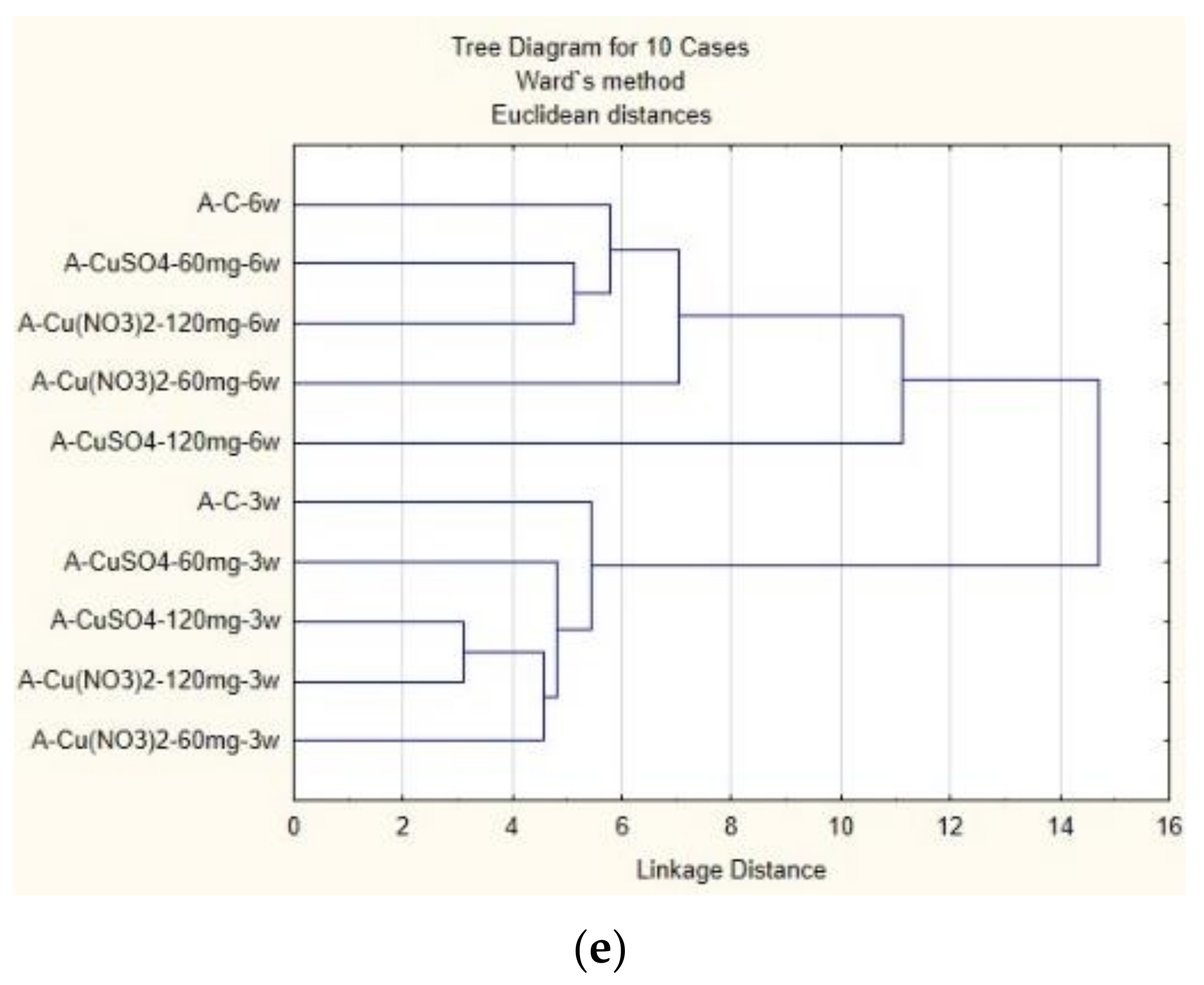
2.7. Cluster Analysis
2.8. Correlation between Bioactive Compounds and Elemental Distribution under the Influence of Sulfate and Nitrate
3. Materials and Methods
3.1. Chemicals and Materials
3.2. Plant Growth Conditions
3.3. Extraction and Analysis of Assimilating Pigments
3.4. Obtaining and Characterizing of Alcoholic Extracts
3.4.1. Determination of Total Polyphenol Content
3.4.2. Determination of the Antioxidant Capacity
3.5. Transmission Electron Microscopy Analysis of the Wheat Leaves
3.6. Elemental Content of the Wheat Biomass and Soil Substrate
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud. 2006, 15, 523–530. [Google Scholar]
- Festa, R.A.; Thiele, D.J. Copper: An essential metal in biology. Curr. Biol. 2011, 21, 877–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, R.A.; Pizarro, M.; Köhler, H.; Sáez, C.A.; Zúñiga, G.E. Copper stress induces antioxidant responses and accumulation of sugars and phytochelatins in Antarctic Colobanthus quitensis (Kunth) Bartl. Biol. Res. 2018, 51, 48. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.H.; Zhang, Y.L.; Wang, X.M.; Cui, J.X.; Xia, X.J.; Shi, K.; Yu, J.Q. Effects of nitrogen form on growth, CO2 assimilation, chlorophyll fluorescence, and photosynthetic electron allocation in cucumber and rice plants. J. Zhejiang Univ. Sci. B. 2011, 12, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Sabir, M.; Hanafi, M.M.; Malik, M.T.; Aziz, T.; Zia-ur-Rehman, M.; Ahmad, H.R.; Hakeem, K.R.; Shahid, M. Differential effect of nitrogen forms on physiological parameters and micronutrient concentration in maize (Zea mays L.). Aust. J. Crop Sci. 2013, 7, 1836–1842. [Google Scholar]
- Sun, Y.D.; Lou, W.R.; Liu, H.C. Effects of different nitrogen forms on the nutritional quality and physiological characteristics of Chinese chive seedlings. Plant Soil Environ. 2014, 60, 216–220. [Google Scholar]
- Chen, L.; Liu, Q.Q.; Gai, J.Y.; Zhu, Y.L.; Yang, L.F.; Wang, C. Effects of nitrogen forms on the growth and polyamine contents in developing seeds of vegetable soybean. J. Plant Nutr. 2011, 2, 504–521. [Google Scholar] [CrossRef]
- Mehrizi, M.H.; Shariatmadari, H.; Khoshgoftarmanesh, A.H.; Dehghani, F. Copper effects on growth, lipid peroxidation, and total phenolic content of rosemary leaves under salinity stress. J. Agric. Sci. Technol. 2012, 14, 205–212. [Google Scholar]
- Ibrahim, M.H.; Jaafar, H.Z.; Rahmat, A.; Rahman, Z.A. Effects of nitrogen fertilization on synthesis of primary and secondary metabolites in three varieties of kacip fatimah (Labisia Pumila Blume). Int. J. Mol. Sci. 2011, 12, 5238–5254. [Google Scholar] [CrossRef]
- Munene, R.; Changamu, E.; Korir, N.; Gweyi, J.O. Effects of different nitrogen forms on growth, phenolics, flavonoids and antioxidant activity in amaranth species. Trop. Plant Res. 2017, 4, 81–89. [Google Scholar] [CrossRef]
- Merlin, T.P.A.; Lima, G.P.P.; Leonel, S.; Vianello, F. Peroxidase activity and total phenol content in citrus cuttings treated with different copper sources. S. Afr. J. Bot. 2012, 83, 159–164. [Google Scholar] [CrossRef]
- Mwamba, T.M.; Ali, S.; Ali, B.; Lwalaba, J.L.; Liu, H.; Farooq, M.A.; Shou, J.; Zhou, W. Interactive effects of cadmium and copper on metal accumulation, oxidative stress, and mineral composition in Brassica napus. Int. J. Environ. Sci. Technol. 2016, 13, 2163–2174. [Google Scholar] [CrossRef]
- Li, L.; Long, M.; Islam, F.; Farooq, M.A.; Wang, J.; Mwamba, T.M.; Shou, J.; Zhou, W. Synergistic effects of chromium and copper on photosynthetic inhibition, subcellular distribution, and related gene expression in Brassica napus cultivars. Environ. Sci. Pollut. Res. 2019, 26, 11827–11845. [Google Scholar] [CrossRef]
- Mwamba, T.M.; Li, L.; Gill, R.A.; Islam, F.; Nawaz, A.; Ali, B.; Farooq, M.A.; Lwalaba, J.L.; Zhou, W. Differential subcellular distribution and chemical forms of cadmium and copper in Brassica napus. Ecotoxicol. Environ. Saf. 2016, 134, 239–249. [Google Scholar] [CrossRef]
- Sonmez, S.; Kaplan, M.; Sonmez, N.K.; Kaya, H.; Uz, I. High level of copper application to soil and leaves reduce the growth and yield of tomato plants. Sci. Agric. 2006, 63, 213–218. [Google Scholar] [CrossRef]
- Lafmejani, Z.N.; Jafari, A.A.; Moradi, P.; Moghadam, A.L. Impact of foliar application of copper sulphate and copper nanoparticles on some morpho-physiological traits and essential oil composition of peppermint (Mentha piperita L.). Herba Pol. 2018, 64, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Krylova, E.G. Seedling Development of Bidens radiate in Solutions of Nickel and Copper Sulfates. Inland Water Biol. 2022, 15, 347–350. [Google Scholar] [CrossRef]
- Krylova, E.G.; Lapirov, A.G.; Berdnik, K.A. Resistance of the initial stages of ontogenesis of Bidens cernua (Asteraceae) to the action of nickel and copper acetates. Vestn. Sev. (Arkt.) Fed. Univ. 2015, 4, 66. [Google Scholar]
- Krylova, E.G.; Vasileva, N.V.; Ivanova, E.S. Resistance of seedlings of native and alien species of genus Bidens (Asteraceae) from different geographical populations to the action of heavy metals. Biosyst. Divers. 2018, 26, 303–308. [Google Scholar] [CrossRef]
- Jańczak-Pieniąźek, M.; Migut, D.; Piechowiak, T.; Balawejder, M. Assessment of the Impact of the Application of a Quercetin—Copper Complex on the Course of Physiological and Biochemical Processes in Wheat Plants (Triticum aestivum L.) Growing under Saline Conditions. Cells 2022, 11, 1141. [Google Scholar] [CrossRef]
- Dey, S.; Mazumder, P.B.; Paul, S.B. Effect of copper on growth and chlorophyll content in tea plants (Camellia sinensis (L.) O. Kuntze). Int. J. Res. Appl. Nat. Soc. Sci. 2014, 2, 223–230. [Google Scholar]
- Giannakoula, A.; Therios, I.; Chatzissavvidis, C. Effect of lead and copper on photosynthetic apparatus in citrus (Citrus aurantium L.) plants. The role of antioxidants in oxidative damage as a response to heavy metal stress. Plants 2021, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- Yruela, I. Copper in plants. Braz. J. Plant Physiol. 2005, 17, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Jadid, N.; Maziyah, R.; Nurcahyani, D.D.; Mubarokah, N.R. Growth and physiological responses of some capsicum frutescens varieties to copper stress. AIP Conf. Proc. 2017, 1854, 020018. [Google Scholar]
- Gratão, P.L.; Alves, L.R.; Lima, L.W. Heavy metal toxicity and plant productivity: Role of metal scavengers. In Plant-Metal Interactions; Srivastava, S., Srivastava, A., Suprasanna, P., Eds.; Springer: Cham, Switzerland, 2019; pp. 49–60. [Google Scholar]
- Piccolella, S.; Crescente, G.; Pacifico, F.; Pacifico, S. Wild aromatic plants bioactivity: A function of their (poly)phenol seasonality? A case study from Mediterranean area. Phytochem. Rev. 2018, 17, 785–799. [Google Scholar] [CrossRef]
- Yin, Y.A.; Qi, J.C.; Li, W.H.; Cao, L.P.; Wang, Z.B. Formation and developmental characteristics of A- and B-type starch granules in wheat endosperm. J. Integr. Agric. 2012, 11, 73–81. [Google Scholar] [CrossRef]
- Buttrose, M.S. Submicroscopic development and structure of starch granules in cereal endosperms 1. J. Ultrastruct. Res. 1960, 4, 231–257. [Google Scholar] [CrossRef] [Green Version]
- Shtangeeva, I.; Venäläinen, S.; Hartikainen, H.; Surgik, M.; Timofeev, S. Silver-induced changes in the nutrient and trace element uptake by wheat and amount of the rhizosphere proteins. J. Plant Nutr. 2011, 34, 1497–1506. [Google Scholar] [CrossRef]
- Stangeeva, I.; Ayrault, S.; Jain, J. Thorium uptake by wheat at different stages of plant growth. J. Environ. Radioact. 2005, 81, 283–293. [Google Scholar] [CrossRef]
- Shtangeeva, I.; Ayrault, S. Effects of Eu and Ca on yield and mineral nutrition of wheat (Triticum aestivum) seedlings. Environ. Exp. Bot. 2007, 59, 49–58. [Google Scholar] [CrossRef]
- Shtangeeva, I. Europium and cerium accumulation in wheat and rye seedlings. Wat. Air Soil Pollut. 2014, 225, 1964–1973. [Google Scholar] [CrossRef]
- Shtangeeva, I.; Niemelä, M.; Perämäki, P.; Timofeev, S. Response of wheat and pea seedlings on increase of bromine concentration in the growth medium. Environ. Sci. Pollut. Res. 2015, 22, 19060–19068. [Google Scholar] [CrossRef]
- Shtangeeva, I.; Niemela, M.; Peramaki, P.; Ryumin, A.; Timofeev, S.; Chukov, S.; Kasatkina, G. Phytoextration of bromine from contaminated soil. J. Geochem. Explor. 2017, 174, 21–28. [Google Scholar] [CrossRef]
- Shtangeeva, I.; Niemelä, M.; Perämäki, P. Effects of soil amendments on antimony uptake by wheat. J. Soils Sediments 2013, 14, 679–686. [Google Scholar] [CrossRef]
- Shtangeeva, I.; Steinnes, E.; Lierhagen, S. Macronutrients and trace elements in rye and wheat: Similarities and differences in uptake and relationships between elements. Environ. Exp. Bot. 2011, 70, 259–265. [Google Scholar] [CrossRef]
- Shtangeeva, I.; Ayrault, S.; Jain, J. Scandium bioaccumulation and its effect on uptake of macro- and trace elements during initial phases of plant growth. Soil Sci. Plant Nutr. 2004, 50, 877–883. [Google Scholar] [CrossRef]
- Soran, M.L.; Lung, I.; Opri, O.; Culicov, O.; Ciorî, A.; Stegarescu, A.; Zinicovscaia, I.; Yushin, N.; Vergel, K.; Kacso, I.; et al. The Effect of TiO2 Nanoparticles on the Composition and Ultrastructure of Wheat. Nanomaterials 2021, 11, 3413. [Google Scholar] [CrossRef]
- Shtangeeva, I.; Steinnes, E.; Lierhagen, S. Uptake of different forms of antimony by wheat and rye seedlings. Environ. Sci. Pollut. Res. 2011, 19, 502–509. [Google Scholar] [CrossRef]
- Zuverza-Mena, N.; Medina-Velo, I.A.; Barrios, A.C.; Tan, W.J.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Copper nanoparticles/compounds impact agronomic and physiological parameters in cilantro (Coriandrum sativum). Environ. Sci. Processes Impacts 2015, 17, 1783–1793. [Google Scholar] [CrossRef]
- Brooks, R.R.; Lee, J.; Reeves, R.D.; Jaffre, T. Detection of nickeliferous rocks by analysis of herbarium specimens of indicator plants. J. Geochem. Explor. 1977, 7, 49–57. [Google Scholar] [CrossRef]
- Baker, A.J.M. Accumulators and excluders strategies in the response of plants to heavy metals. J. Plant Nutr. 1981, 3, 643–654. [Google Scholar] [CrossRef]
- McGrath, S.P.; Zhao, F.J. Phytoextraction of metals and metalloids from contaminated soils. Curr. Opin. Biotechnol. 2003, 14, 277–282. [Google Scholar] [CrossRef]
- Yoon, J.; Cao, X.D.; Zhou, Q.X.; Ma, L.Q. Accumulation of Pb, Cu and Zn in native plants growing on a contaminated Florida site. Sci. Total Environ. 2006, 368, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Sánchez, M.J.; García-Lorenzo, M.L.; Pérez-Sirvent, C.; Bech, J. Trace element accumulation in plants from an acidic area affected by mining activities. J. Geochem. Explor. 2012, 123, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Bech, J.; Duran, P.; Roca, N.; Poma, W.; Sánchez, I.; Barceló, J.; Boluda, R.; Roca Pérez, L.; Poschenrieder, C. Shoot accumulation of several trace elements in native plant species from contaminated soils in the Peruvian Andes. J. Geochem. Explor. 2012, 113, 106–111. [Google Scholar] [CrossRef]
- Lung, I.; Opris, O.; Soran, M.L.; Culicov, O.; Ciorîta, A.; Stegarescu, A.; Zinicovscaia, I.; Yushin, N.; Vergel, K.; Kacso, I.; et al. The impact assessment of CuO nanoparticles on the composition and ultrastructure of Triticum aestivum L. Int. J. Environ. Res. Public Health 2021, 18, 6739. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Current Protocols in Food Analytical Chemistry (Units: F4.3.1–F4.3.8); John Wiley & Sons Inc.: New York, NY, USA, 2001. [Google Scholar]
- Ivanova, V.; Stefova, M.; Chinnici, F. Determination of the polyphenol contents in Macedonian grapes and wines by standardized spectrophotometric methods. J. Serb. Chem. Soc. 2010, 75, 45–59. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Pavlov, S.S.; Dmitriev, A.Y.; Frontasyeva, M.V. Automation system for neutron activation analysis at the reactor IBR-2. Frank Laboratory of neutron physics. Joint Institute for Nuclear Research, Dubna, Russia. J. Radioanal. Nucl. Chem. 2016, 309, 27–38. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exerimental Data for Wheat | Literature Data for Wheat | References | ||||||
|---|---|---|---|---|---|---|---|---|
| Soil 3w | Soil 6w | Root | A 3w | A 6w | Root Control | A Control | ||
| Na | 3020–7700 | 2820–4600 | 1040–1650 | 380–410 | 390–470 | 2900–7710 | 250–10,000 | [29,33,34] |
| Mg * | 5.1–13 | 9.6–18.7 | 2.6–3.3 | 2.0–2.9 | 0.49–2.6 | 0.07–2.7 | 0.079–2.2 | [29,33,35,36] |
| Al * | 7.9–24.4 | 15.4–23 | 0.26–0.71 | 0.070–0.084 | 0.020–0.071 | 0.118–0.314 | 0.0059–0.067 | [29,36,37] |
| Cl * | 0.59–1.4 | 0.72–1.5 | 1.3–4.3 | 9.3–12.6 | 2.5–23 | 3.4–5.4 | 5.5–27 | [33,35,37] |
| K * | 5.9–9.2 | 6–7.5 | 49–60 | 101–108 | 126–148 | 3–351 | 8.6–657 | [32,33] |
| Ca * | 33–50 | 46–61 | 4.7–6.7 | 3.2–4.9 | 0.74–6.5 | 0.26–5.6 | 0.05–5.1 | [29,33,35,36] |
| Sc | 2.6–4.4 | 2.5–4.2 | 0.058–0.19 | 0.014–0.026 | 0.011–0.039 | 0.015–0.18 | 0.01–0.15 | [34,37] |
| V | 14–32.9 | 23.1–38.9 | 0.63–1.5 | ND | ND | 0.071 | 0.04 | [37] |
| Cr | 14.3–31 | 14.2–24.4 | 1.5–8.2 | ND | 1.2–4.9 | 0.68–12.3 | 0.41–6.6 | [29,37] |
| Mn | 350–630 | 520–580 | 25.4–48 | 24.2–35 | 3.2–21.4 | 6.9–63 | 2–44 | [29,33] |
| Fe * | 8–13.8 | 7–12.9 | 0.3–0.68 | 0.12–0.17 | 0.12–0.17 | 0.030–0.84 | 0.15–1.42 | [34,35,36,37] |
| Co | 3.5–5 | 3.1–5 | 0.35–0.56 | 0.039–0.054 | 0.048–0.096 | 0.002–0.88 | 0.013–0.08 | [32,34] |
| Cu | 37.6–182 | 60.4–262 | 32.8–217 | 13.6–40.6 | 10.5–49 | 0.08–12 | 0.14–104 | [32,33,37] |
| Zn | 66–101 | 69–80 | 431–538 | 80–92 | 50–67 | 25.9–329 | 35.7–200 | [33,34,35] |
| As | 4.5–5.8 | 3.9–5.9 | 3.6–5 | 0.67–0.84 | 0.66–0.87 | 0.042–0.58 | 0.01–0.43 | [37,38] |
| Br | 8.1–11 | 10.3–14.1 | 5.42–8.5 | 9.9–11.1 | 4.88–6.8 | 12.4–33 | 7.5–11 | [34,38] |
| Rb | 26–42 | 21–34 | 27–34 | 43–46 | 44–48 | 5.2–61 | 6.8–88 | [34,38] |
| Sr | 96–154 | 90–123 | 18.8–23.7 | 11.9–13.4 | 9.6–18 | 0.24 | 0.1–37.4 | [39] |
| Mo | 1–1.4 | 0.81–3.8 | 0.58–0.88 | 1.7–2 | 1.6–2.5 | 0.31 | 0.47 | [37] |
| Sb | 2–2.8 | 1.8–3.2 | 0.19–0.3 | 0.037–0.076 | 0.018–0.048 | 0.02–1.94 | 0.02–0.98 | [32,37] |
| Cs | 1.2–2.1 | 1.1–1.5 | 0.027–0.12 | 0.0076–0.019 | 0.017–0.05 | 0.014–0.77 | 0.01–0.26 | [37,38] |
| Ba | 240–410 | 230–330 | 24–39 | 25–33 | 28–53 | 12.9–33 | 9.8–158 | [36,37] |
| La | 8.7–14.4 | 7.2–11.9 | 0.18–0.42 | ND | ND | 0.3–0.36 | 0.07 | [34,38] |
| Sm | 1.6–2.5 | 1.3–2.1 | 0.028–0.074 | 0.0036–0.019 | 0.0021–0.016 | 0.04–0.1 | 0.008–0.01 | [34,38] |
| Ta | 0.27–0.44 | 0.22–0.38 | 0.0087–0.98 | 0.013–0.021 | ND | <0.2 | <0.2 | [38] |
| Au | 0.0016–0.032 | 0.0019–0.0068 | 0.005–0.044 | 0.013–0.057 | 0.044–0.066 | 0.008–0.02 | 0.005–0.01 | [34,39] |
| Th | 2.5–4 | 2.2–3.4 | 0.066–0.14 | 0.010–0.017 | 0.0146–0.16 | <0.02–0.09 | <0.007–<0.02 | [34,38,39] |
| U | 0.81–1.32 | 0.74–1.14 | 0.17–0.28 | ND | ND | na | <0.03 | [38] |
| CuSO4 vs. Cu(NO3)2 | CuSO4 | Cu(NO3)2 | |||||
|---|---|---|---|---|---|---|---|
| R−R | A 3w−3w | A 6w−6w | R−A 6w | A 3w−6w | R−A 6w | A 3w−6w | |
| Na | 0.96 | 0.96 | 0.00 | −0.78 | −0.28 | 0.81 | −1.00 |
| Mg | −0.52 | 1.00 | 0.27 | 0.32 | −0.53 | −0.47 | 0.74 |
| Al | 0.56 | −0.99 | 0.56 | −0.25 | −0.31 | −0.25 | 0.92 |
| Cl | 0.97 | 0.57 | 0.99 | −0.95 | −0.69 | −0.75 | 0.03 |
| K | 0.97 | −1.00 | 0.92 | −0.54 | −0.56 | −0.94 | 0.13 |
| Ca | 0.60 | −0.19 | 0.85 | −0.33 | −0.99 | −0.97 | −0.45 |
| Sc | 0.74 | −0.54 | 0.78 | 0.03 | −0.81 | 0.99 | −0.66 |
| V | 0.69 | ND | ND | ND | ND | ND | ND |
| Cr | 0.63 | −0.01 | ND | ND | ND | ND | ND |
| Mn | 0.96 | 0.49 | −0.96 | 0.58 | −0.46 | −0.57 | −0.76 |
| Fe | 0.60 | −0.11 | 0.97 | 0.39 | −0.81 | 1.00 | −0.70 |
| Co | 1.00 | −0.73 | −0.16 | −0.99 | 1.00 | 0.36 | 0.76 |
| Cu | 0.95 | −0.34 | −0.18 | 0.78 | −0.57 | 0.18 | 0.09 |
| Zn | 0.20 | 0.92 | 0.72 | −0.29 | 0.73 | −0.65 | 0.92 |
| As | 0.80 | −1.00 | 0.65 | −0.90 | 0.39 | −0.48 | 0.51 |
| Br | 0.94 | 0.98 | 0.16 | −0.87 | 0.93 | −0.33 | 0.66 |
| Rb | 0.87 | −0.94 | 0.94 | −0.19 | −0.76 | −0.87 | 0.76 |
| Sr | 0.48 | 0.23 | −0.17 | −0.14 | −0.99 | −0.19 | −0.97 |
| Mo | −0.58 | −0.61 | 0.87 | 0.24 | 0.99 | 0.19 | −0.24 |
| Ag | 1.00 | 0.19 | 0.75 | 0.94 | 0.99 | 0.53 | −0.36 |
| Cd | 0.73 | 0.83 | −0.55 | −0.86 | −0.85 | −0.65 | −0.54 |
| Sb | 1.00 | 0.77 | 0.14 | −0.99 | 0.92 | −0.30 | 0.94 |
| Cs | 0.98 | 0.98 | 0.56 | 0.51 | −0.67 | 0.99 | 0.06 |
| Ba | 0.97 | 0.71 | 0.80 | 0.98 | 1.00 | 0.98 | 1.00 |
| La | 0.66 | ND | ND | ND | ND | ND | ND |
| Sm | 0.30 | 1.00 | 0.75 | −0.05 | −0.72 | 0.93 | −0.10 |
| Gd | ND | 0.85 | ND | ND | ND | ND | |
| Ta | −0.36 | ND | 0.56 | −0.33 | ND | −0.96 | ND |
| Au | 0.81 | −0.70 | 1.00 | −0.87 | 0.17 | −0.44 | −0.84 |
| Th | 0.66 | 0.99 | 0.81 | 0.35 | 0.94 | 0.97 | 0.92 |
| U | −0.54 | ND | ND | ND | ND | ND | ND |
| A 6w CuSO4 | A 6w Cu(NO3)2 | A 3w CuSO4 | A 3w Cu(NO3)2 | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chla | Chlb | CARO | TP | DPPH | Chla | Chlb | CARO | TP | DPPH | Chla | Chlb | CARO | TP | DPPH | Chla | Chlb | CARO | TP | DPPH | |
| Cha | 1 | 1 | 1 | 1 | ||||||||||||||||
| Chb | 1.00 | 1 | 1.00 | 1 | 1.00 | 1 | 1.00 | 1 | ||||||||||||
| CARO | 1.00 | 1.00 | 1 | 1.00 | 1.00 | 1 | 1.00 | 1.00 | 1 | 1.00 | 1.00 | 1 | ||||||||
| TP | −0.96 | −0.95 | −0.97 | 1 | −0.93 | −0.96 | −0.94 | 1 | −0.99 | −0.98 | −0.98 | 1 | −0.99 | −0.99 | −0.99 | 1 | ||||
| DPPH | −0.03 | 0.00 | −0.05 | 0.31 | 1 | −0.31 | −0.23 | −0.28 | −0.06 | 1 | −0.98 | −0.98 | −0.98 | 0.92 | 1 | −0.67 | −0.65 | −0.66 | 0.76 | 1 |
| Na | 0.12 | 0.16 | 0.11 | 0.16 | 0.99 | 0.51 | 0.58 | 0.53 | −0.79 | 0.66 | −0.95 | −0.95 | −0.96 | 0.88 | 0.99 | 0.47 | 0.45 | 0.47 | −0.58 | −0.97 |
| Mg | −0.63 | −0.60 | −0.64 | 0.82 | 0.80 | −0.60 | −0.53 | −0.58 | 0.26 | 0.95 | 0.25 | 0.25 | 0.21 | −0.41 | −0.03 | 0.99 | 0.99 | 0.99 | −0.96 | −0.55 |
| Al | −0.28 | −0.25 | −0.30 | 0.54 | 0.97 | 0.02 | −0.06 | −0.01 | 0.35 | −0.96 | −1.00 | −1.00 | −1.00 | 0.99 | 0.96 | −0.46 | −0.49 | −0.47 | 0.34 | −0.35 |
| Cl | −0.72 | −0.70 | −0.74 | 0.89 | 0.71 | 0.28 | 0.35 | 0.30 | −0.61 | 0.83 | −0.99 | −0.99 | −0.99 | 1.00 | 0.94 | −0.98 | −0.98 | −0.98 | 0.95 | 0.50 |
| K | 0.89 | 0.87 | 0.90 | −0.98 | −0.49 | −0.8 | −0.82 | −0.85 | 0.61 | 0.75 | 0.20 | 0.20 | 0.24 | −0.03 | −0.41 | −0.35 | −0.33 | −0.34 | 0.47 | 0.93 |
| Ca | −0.79 | −0.77 | −0.80 | 0.93 | 0.64 | −0.56 | −0.62 | −0.58 | 0.83 | −0.62 | −0.90 | −0.90 | −0.91 | 0.81 | 0.97 | −1.00 | −1.00 | −1.00 | 0.98 | 0.61 |
| Sc | 0.58 | 0.56 | 0.60 | −0.79 | −0.83 | 0.49 | 0.55 | 0.51 | −0.78 | 0.68 | 0.97 | 0.97 | 0.98 | −0.92 | −1.00 | 0.97 | 0.96 | 0.96 | −0.99 | −0.84 |
| Cr | 0.69 | 0.71 | 0.67 | −0.46 | 0.70 | −0.64 | −0.57 | −0.62 | 0.31 | 0.93 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| Mn | −0.88 | −0.86 | −0.89 | 0.98 | 0.51 | −0.91 | −0.94 | −0.92 | 1.00 | −0.11 | −0.06 | −0.06 | −0.09 | −0.11 | 0.28 | −0.38 | −0.35 | −0.37 | 0.49 | 0.94 |
| Fe | 0.82 | 0.80 | 0.83 | −0.95 | −0.59 | 0.59 | 0.65 | 0.61 | −0.85 | 0.59 | 0.99 | 0.99 | 0.99 | −1.00 | −0.95 | 0.98 | 0.98 | 0.98 | −1.00 | −0.79 |
| Co | 0.94 | 0.95 | 0.93 | −0.80 | 0.32 | −0.56 | −0.62 | −0.58 | 0.82 | −0.62 | −0.88 | −0.88 | −0.86 | 0.95 | 0.75 | −0.16 | −0.19 | −0.17 | 0.03 | −0.63 |
| Cu | 0.30 | 0.33 | 0.28 | −0.02 | 0.94 | 0.71 | 0.65 | 0.69 | −0.40 | −0.89 | −0.68 | −0.68 | −0.70 | 0.54 | 0.82 | −0.73 | −0.71 | −0.73 | 0.81 | 1.00 |
| Zn | 0.81 | 0.83 | 0.80 | −0.61 | 0.56 | 0.99 | 1.00 | 1.00 | −0.96 | −0.21 | −1.00 | −1.00 | −1.00 | 0.98 | 0.98 | −0.85 | −0.84 | −0.85 | 0.91 | 0.96 |
| As | 0.04 | 0.07 | 0.02 | 0.24 | 1.00 | −0.24 | −0.31 | −0.26 | 0.58 | −0.85 | 0.94 | 0.94 | 0.95 | −0.86 | −0.99 | 0.94 | 0.93 | 0.94 | −0.98 | −0.87 |
| Br | 0.96 | 0.97 | 0.96 | −0.85 | 0.24 | 1.00 | 0.99 | 1.00 | −0.91 | −0.36 | −0.75 | −0.75 | −0.72 | 0.85 | 0.59 | −0.65 | −0.67 | −0.66 | 0.55 | −0.13 |
| Rb | 0.99 | 1.00 | 0.99 | −0.92 | 0.09 | −0.94 | −0.91 | −0.93 | 0.75 | 0.61 | 0.87 | 0.87 | 0.89 | −0.77 | −0.96 | 0.53 | 0.55 | 0.53 | −0.41 | 0.28 |
| Sr | −0.99 | −1.00 | −0.99 | 0.92 | −0.09 | −0.24 | −0.32 | −0.27 | 0.58 | −0.85 | −1.00 | −1.00 | −1.00 | 0.99 | 0.97 | 0.03 | 0.06 | 0.04 | 0.10 | 0.73 |
| Mo | −0.69 | −0.66 | −0.70 | 0.86 | 0.75 | −0.29 | −0.36 | −0.32 | 0.62 | −0.82 | 0.66 | 0.66 | 0.69 | −0.53 | −0.81 | 0.88 | 0.89 | 0.89 | −0.81 | −0.23 |
| Ag | −0.92 | −0.91 | −0.93 | 0.99 | 0.42 | −0.67 | −0.72 | −0.68 | 0.89 | −0.51 | 0.99 | 0.99 | 0.99 | −0.94 | −1.00 | 0.49 | 0.52 | 0.50 | −0.38 | 0.32 |
| Cd | 0.76 | 0.74 | 0.78 | −0.91 | −0.67 | 0.97 | 0.95 | 0.97 | −0.82 | −0.52 | 0.38 | 0.38 | 0.41 | −0.22 | −0.57 | 0.69 | 0.67 | 0.69 | −0.78 | −1.00 |
| Sb | 0.91 | 0.92 | 0.90 | −0.75 | 0.39 | 0.93 | 0.89 | 0.92 | −0.72 | −0.64 | −0.61 | −0.61 | −0.58 | 0.74 | 0.42 | −0.78 | −0.79 | −0.78 | 0.69 | 0.05 |
| Cs | 0.34 | 0.31 | 0.35 | −0.58 | −0.95 | 0.38 | 0.31 | 0.36 | −0.02 | −1.00 | 0.95 | 0.95 | 0.96 | −0.89 | −1.00 | 0.88 | 0.86 | 0.87 | −0.93 | −0.94 |
| Ba | −0.82 | −0.80 | −0.83 | 0.95 | 0.60 | −1.00 | −1.00 | −1.00 | 0.94 | 0.27 | 0.91 | 0.91 | 0.92 | −0.82 | −0.98 | 1.00 | 1.00 | 1.00 | −1.00 | −0.69 |
| Sm | 0.78 | 0.76 | 0.79 | −0.92 | −0.65 | 0.71 | 0.65 | 0.69 | −0.40 | −0.89 | 0.98 | 0.98 | 0.98 | −1.00 | −0.92 | 0.74 | 0.73 | 0.74 | −0.82 | −0.99 |
| Gd | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | 0.99 | 0.99 | 0.99 | −1.00 | −0.94 | 0.87 | 0.86 | 0.87 | −0.93 | −0.94 |
| Ta | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | 0.85 | 0.85 | 0.84 | −0.93 | −0.72 | 0.22 | 0.19 | 0.21 | −0.35 | −0.87 |
| Au | −0.92 | −0.91 | −0.93 | 0.99 | 0.42 | 0.24 | 0.16 | 0.21 | 0.13 | −1.00 | −0.15 | −0.15 | −0.11 | 0.32 | −0.07 | −0.29 | −0.26 | −0.28 | 0.41 | 0.91 |
| Th | −0.95 | −0.96 | −0.95 | 0.83 | −0.28 | −1.00 | −1.00 | −1.00 | 0.94 | 0.28 | 1.00 | 1.00 | 1.00 | −0.99 | −0.97 | 0.89 | 0.87 | 0.88 | −0.94 | −0.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Culicov, O.; Stegarescu, A.; Soran, M.-L.; Lung, I.; Opriș, O.; Ciorîță, A.; Nekhoroshkov, P. The Effect of Copper Salts on Bioactive Compounds and Ultrastructure of Wheat Plants. Molecules 2022, 27, 4835. https://doi.org/10.3390/molecules27154835
Culicov O, Stegarescu A, Soran M-L, Lung I, Opriș O, Ciorîță A, Nekhoroshkov P. The Effect of Copper Salts on Bioactive Compounds and Ultrastructure of Wheat Plants. Molecules. 2022; 27(15):4835. https://doi.org/10.3390/molecules27154835
Chicago/Turabian StyleCulicov, Otilia, Adina Stegarescu, Maria-Loredana Soran, Ildiko Lung, Ocsana Opriș, Alexandra Ciorîță, and Pavel Nekhoroshkov. 2022. "The Effect of Copper Salts on Bioactive Compounds and Ultrastructure of Wheat Plants" Molecules 27, no. 15: 4835. https://doi.org/10.3390/molecules27154835
APA StyleCulicov, O., Stegarescu, A., Soran, M. -L., Lung, I., Opriș, O., Ciorîță, A., & Nekhoroshkov, P. (2022). The Effect of Copper Salts on Bioactive Compounds and Ultrastructure of Wheat Plants. Molecules, 27(15), 4835. https://doi.org/10.3390/molecules27154835