


Thiourea-Capped Nanoapatites Amplify Osmotic Stress Tolerance in Zea mays L. by Conserving Photosynthetic Pigments, Osmolytes Biosynthesis and Antioxidant Biosystems

 , , ,
, , ,  , ,
, , 
Abstract
:1. Introduction
2. Results
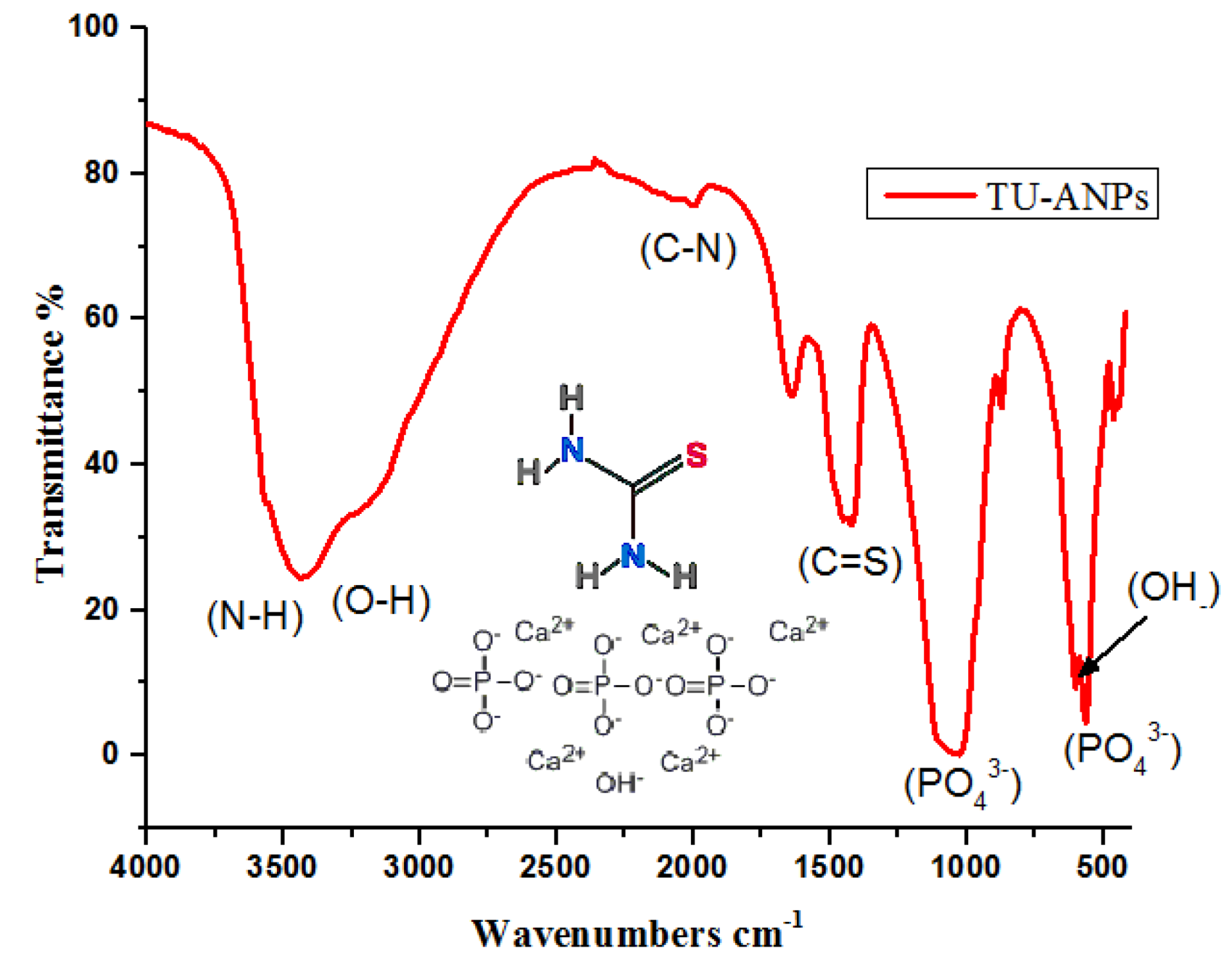
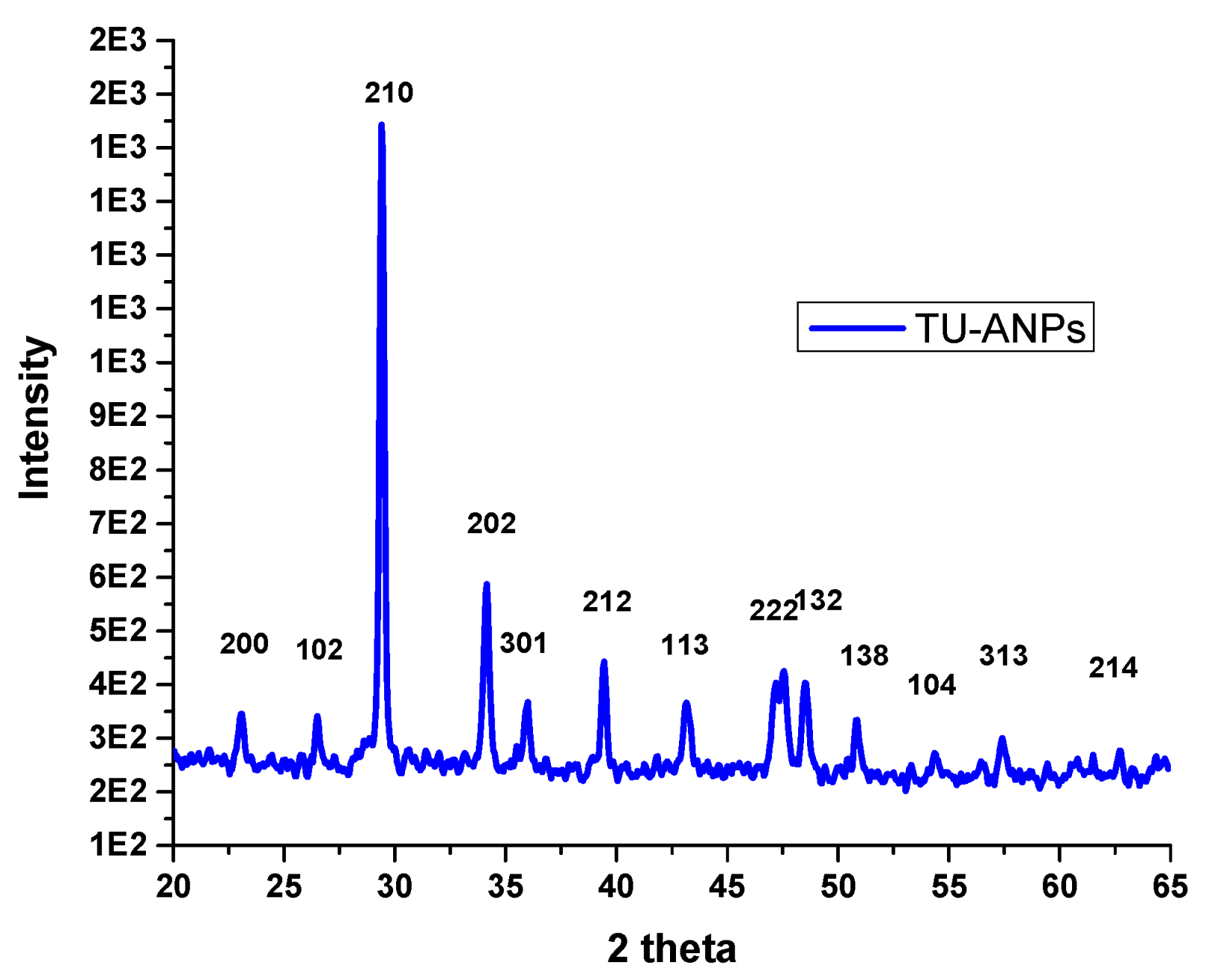
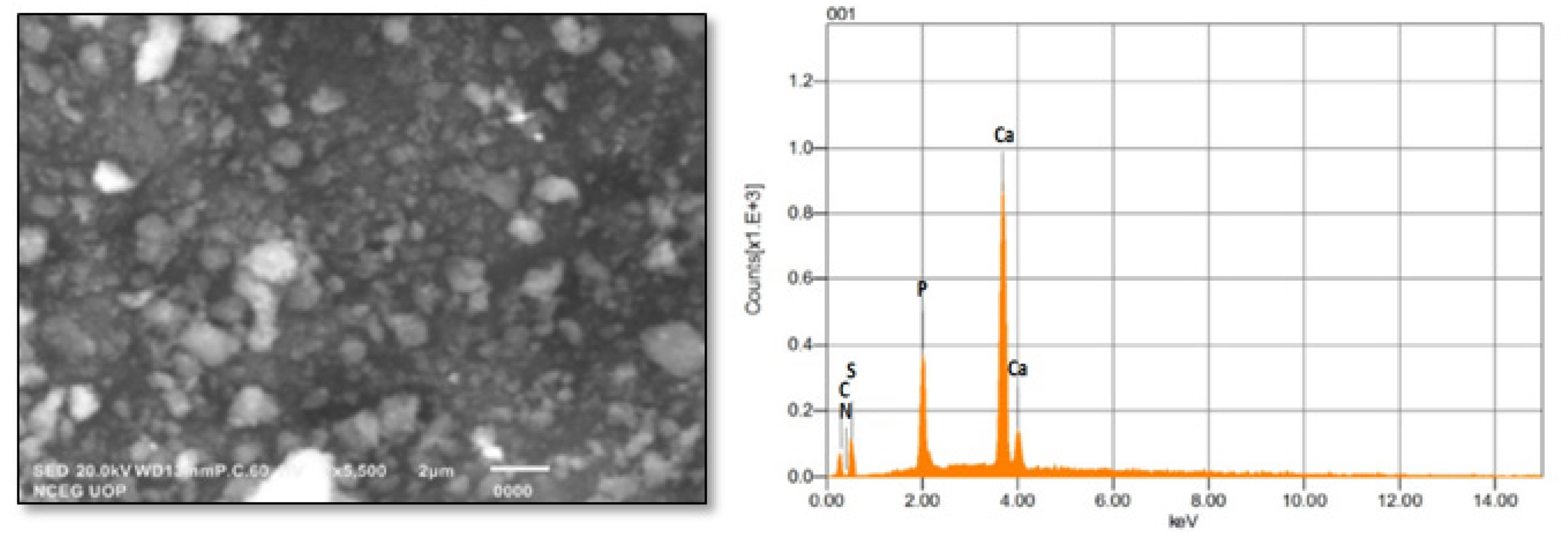
2.1. Characterization of TU–ANPs
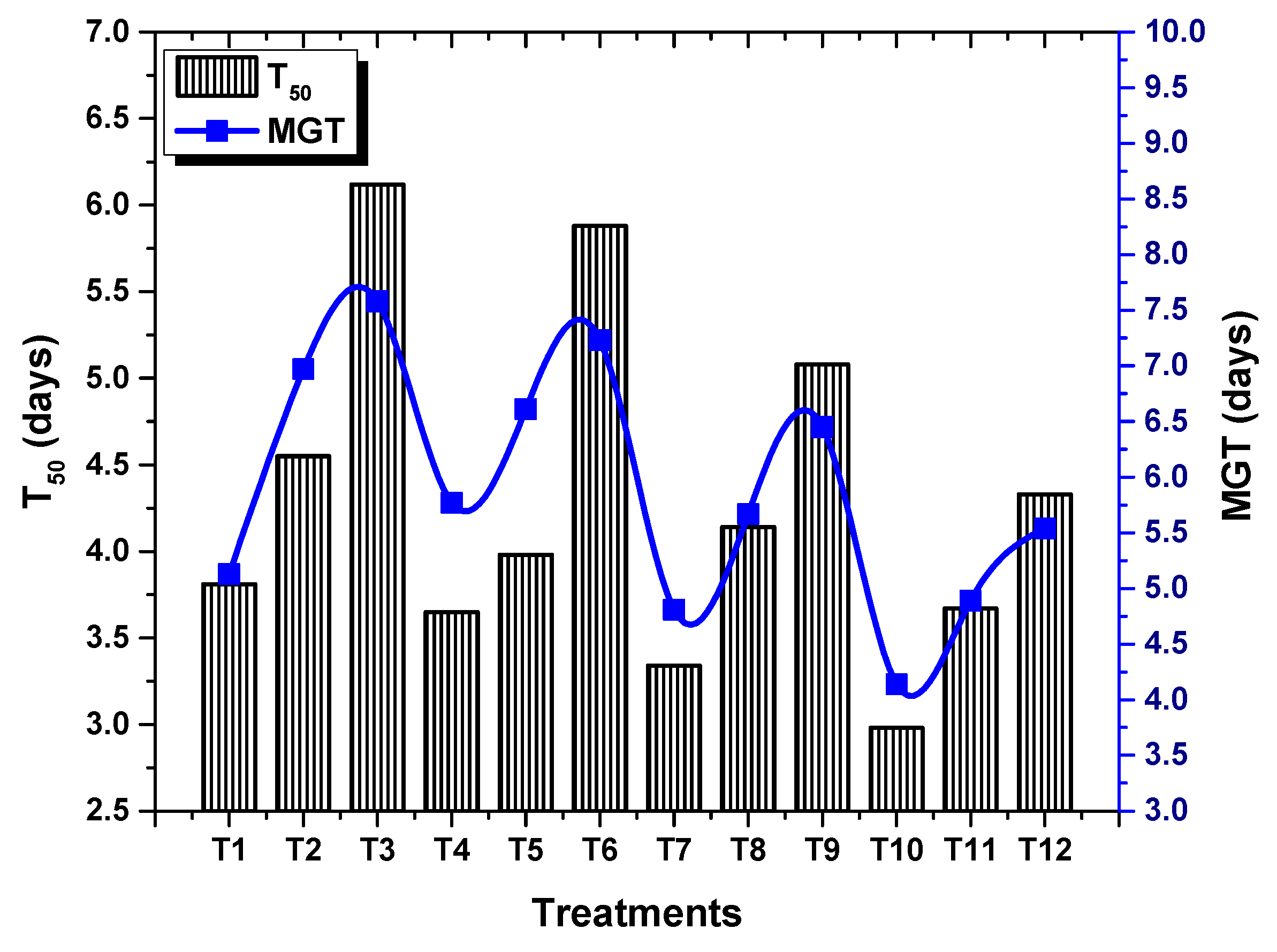
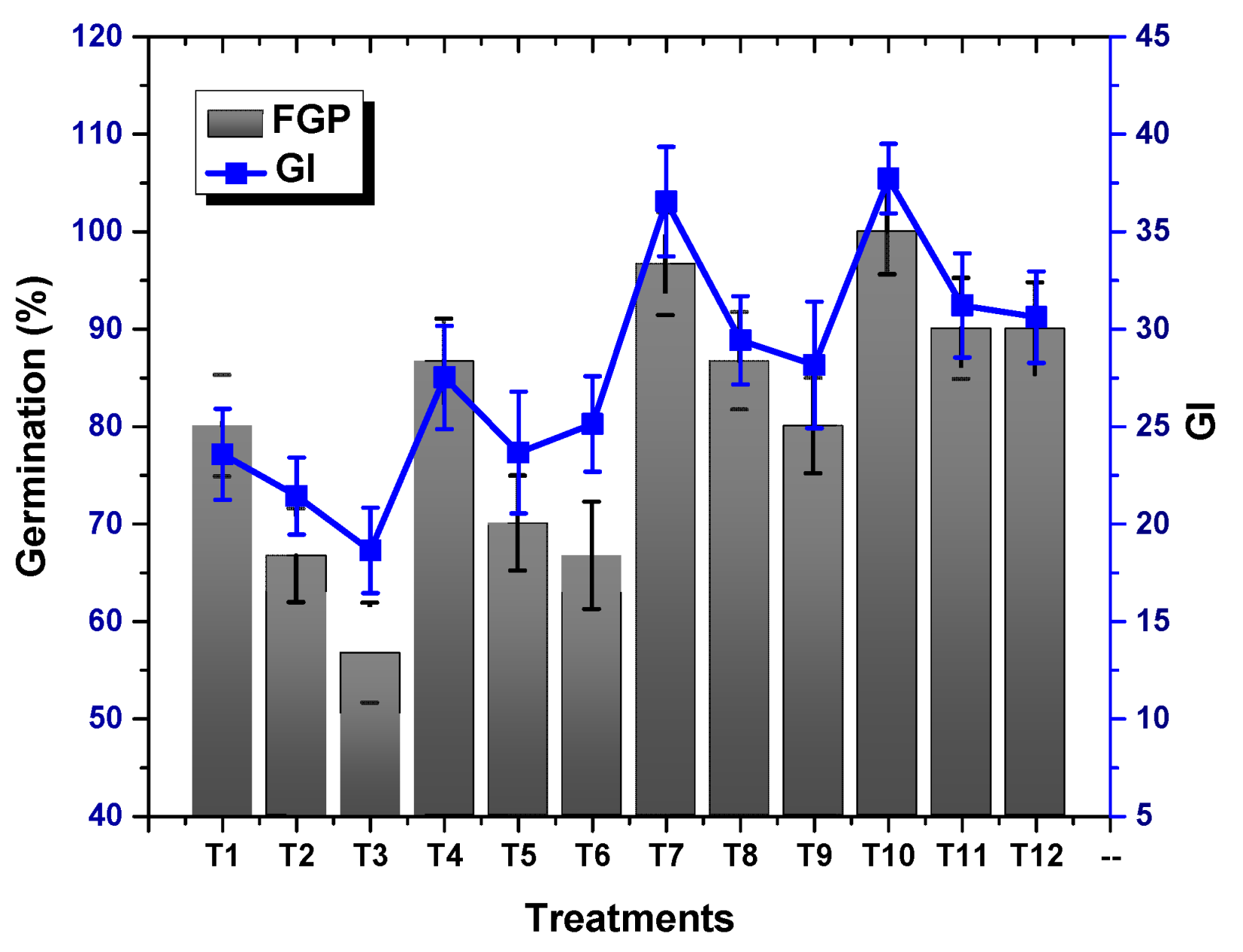
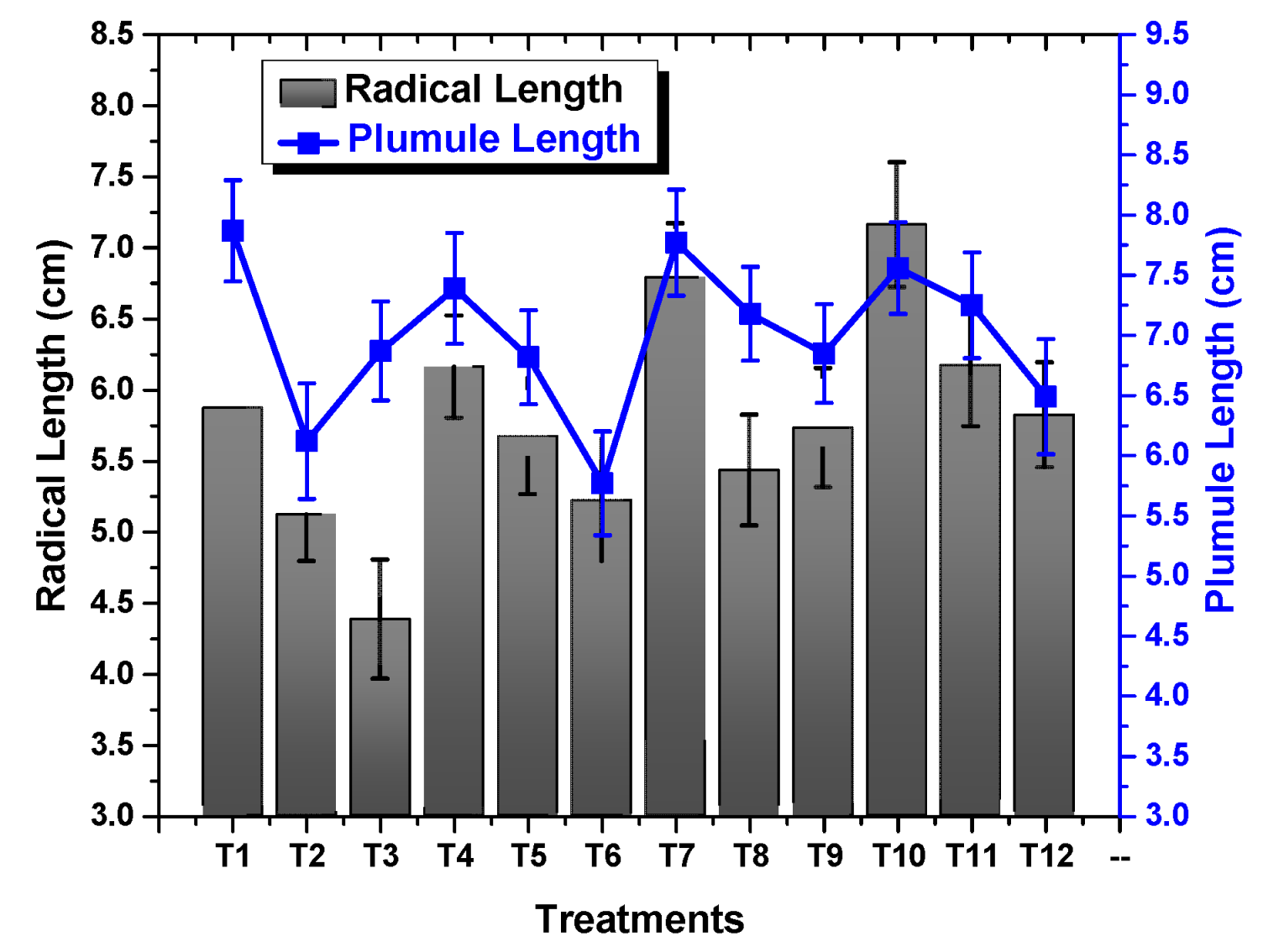
2.2. Effect of TU–ANPs on Germination Indices and Vegetative Performance
2.3. Effect of TU–ANPs on Photosynthetic Pigments and Osmolytes
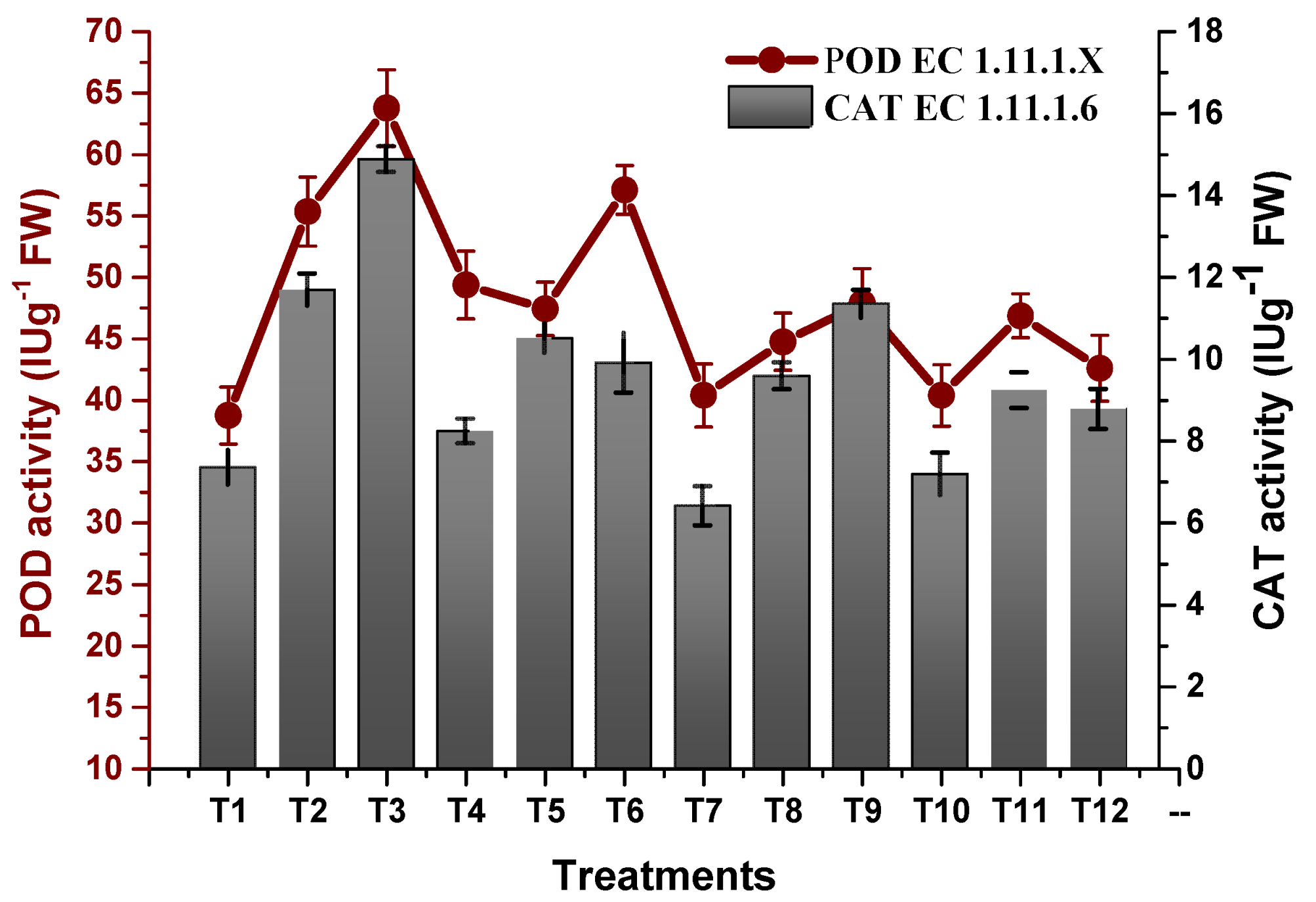
2.4. Effect of TU–ANPs on Antioxidation Biosystem
3. Discussion
4. Material and Methods
4.1. Synthesis and Characterization of TU–ANPs
4.2. Plant Material and Growth Conditions
4.3. Physiological and Biochemical Analysis
4.4. Activity of Antioxidant Enzymes (POD, APX, CAT and SOD)
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roychowdhury, D.; Mondal, S.; Banerjee, S.K. The effect of biofertilizers and the effect of vermicompost on the cultivation and productivity of maize-a review. Adv. Crop Sci. Technol. 2017, 5, 1–4. [Google Scholar] [CrossRef]
- Ali, B.; Hafeez, A.; Javed, M.A.; Ahmad, S.; Afridi, M.S.; Sumaira Nadeem, M.; Khan, A.U.R.; Malik, A.; Ullah, A.; Alwahibi, M.S.; et al. Bacterial-mediated salt tolerance in maize: Insights into plant growth promotion, antioxidant defense system, oxidative stress, and surfactant production. Front. Plant Sci. 2022, 13, 978291. [Google Scholar] [CrossRef]
- Ali, B.; Hafeez, A.; Ahmad, S.; Javed, M.A.; Sumaira Afridi, M.S.; Dawoud, T.M.; Almaary, K.S.; Muresan, C.C.; Marc, R.A.; Alkhalifah, D.H.M.; et al. Bacillus thuringiensis PM25 ameliorates oxidative damage of salinity stress in maize via regulating growth, leaf pigments, antioxidant defense system, and stress responsive gene expression. Front. Plant Sci. 2022, 13, 921668. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Wang, X.; Saleem, M.H.; Azeem, M.A.; Afridi, M.S.; Nadeem, M.; Ghazal, M.; Batool, T.; Qayyum, A.; Alatawi, A.; et al. Bacillus mycoides PM35 Reinforces Photosynthetic Efficiency, Antioxidant Defense, Expression of Stress-Responsive Genes, and Ameliorates the Effects of Salinity Stress in Maize. Life 2022, 12, 219. [Google Scholar] [CrossRef]
- Ali, B.; Wang, X.; Saleem, M.H.; Hafeez, A.; Afridi, M.S.; Khan, S.; Ullah, I.; Amaral Júnior, A.T.; Alatawi, A.; Ali, S. PGPR-Mediated Salt Tolerance in Maize by Modulating Plant Physiology, Antioxidant Defense, Compatible Solutes Accumulation and Bio-Surfactant Producing Genes. Plants 2022, 11, 345. [Google Scholar] [CrossRef]
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The new perspective in precision agriculture. Biotechnol. Rep. 2017, 125, 11–23. [Google Scholar] [CrossRef]
- GOP. Economic Survey. Government of Pakistan, Finance Division, Economic Adviser’s Wing, Islamabad; GOP: Washington, DC, USA, 2005. [Google Scholar]
- WB. Pakistan Strategic Country Environmental Assessment; World Bank: Washington, DC, USA, 2006; Volume II. [Google Scholar]
- Sher, A.; Wang, X.; Sattar, A.; Ijaz, M.; Ul-Allah, S.; Nasrullah, M.; Bibi, Y.; Manaf, A.; Fiaz, S.; Qayyum, A. Exogenous Application of Thiourea for Improving the Productivity and Nutritional Quality of Bread Wheat (Triticum aestivum L.). Agronomy 2021, 11, 1432. [Google Scholar] [CrossRef]
- Hassanein, R.A.; Amin AA, E.; Rashad ES, M.; Ali, H. Effect of thiourea and salicylic acid on antioxidant defense of wheat plants under drought stress. Int. J. ChemTech Res. 2015, 7, 346–354. [Google Scholar]
- Mehmood, S.; Khatoon, Z.; Amna; Ahmad, I.; Muneer, M.A.; Kamran, M.A.; Ali, J.; Ali, B.; Chaudhary, H.J.; Munis, M.F.H. Bacillus sp. PM31 harboring various plant growth-promoting activities regulates Fusarium dry rot and wilt tolerance in potato. Arch. Agron. Soil Sci. 2021, 2021, 1–15. [Google Scholar] [CrossRef]
- Nawaz, H.; Ali, A.; Saleem, M.H.; Ameer, A.; Hafeez, A.; Alharbi, K.; Khan, A.; Jamil, M.; Farid, G. Comparative effectiveness of EDTA and citric acid assisted phytoremediation of Ni contaminated soil by using canola (Brassica napus). Braz. J. Biol. 2022, 82, e261785. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Sönmez, O. Promotive effect of exogenously applied thiourea on key physiological parametersand oxidative defense mechanism in salt-stressed Zea mays L. plants. Turk. J. Bot. 2015, 39, 786–795. [Google Scholar] [CrossRef]
- Dola, D.B.; Mannan, M.A.; Sarker, U.; Mamun, M.A.A.; Islam, T.; Ercisli, S.; Saleem, M.H.; Ali, B.; Pop, O.L.; Marc, R.A. Nano-iron oxide accelerates growth, yield, and quality of Glycine max seed in water deficits. Front. Plant Sci. 2022, 13, 992535. [Google Scholar] [CrossRef]
- Farooq, T.H.; Rafay, M.; Basit, H.; Shakoor, A.; Shabbir, R.; Riaz, M.U.; Ali, B.; Kumar, U.; Qureshi, K.A.; Jaremko, M. Morpho-physiological growth performance and phytoremediation capabilities of selected xerophyte grass species toward Cr and Pb stress. Front. Plant Sci. 2022, 13, 997120. [Google Scholar] [CrossRef]
- Marchiol, L.; Filippi, A.; Adamiano, A.; Degli Esposti, L.; Iafisco, M.; Mattiello, A.; Braidot, E. Influence of hydroxyapatite nanoparticles on germination and plant metabolism of tomato (Solanum lycopersicum L.): Preliminary evidence. Agronomy 2019, 9, 161. [Google Scholar] [CrossRef]
- Fatima, F.; Hashim, A.; Anees, S. Efficacy of nanoparticles as nanofertilizer production: A review. Environ. Sci. Pollut. Res. 2021, 28, 1292–1303. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 231–239. [Google Scholar] [CrossRef]
- Saeed, S.; Ullah, A.; Ullah, S.; Noor, J.; Ali, B.; Khan, M.N.; Hashem, M.; Mostafa, Y.S.; Alamri, S. Validating the Impact of Water Potential and Temperature on Seed Germination of Wheat (Triticum aestivum L.) via Hydrothermal Time Model. Life 2022, 12, 983. [Google Scholar] [CrossRef]
- Haider, M.W.; Nafees, M.; Ahmad, I.; Ali, B.; Maryam Iqbal, R.; Vodnar, D.C.; Marc, R.A.; Kamran, M.; Saleem, M.H.; Al-Ghamdi, A.A.; et al. Postharvest dormancy-related changes of endogenous hormones in relation to different dormancy-breaking methods of potato (Solanum tuberosum L.) tubers. Front. Plant Sci. 2022, 13, 945256. [Google Scholar] [CrossRef]
- Hussain, S.Q.; Rasheed, M.; Saleem, M.H.; Ahmed, Z.I.; Hafeez, A.; Jilani, G.; Alamri, S.; Hashem, M.; Ali, S. Salt tolerance in maize with melatonin priming to achieve sustainability in yield on salt affected soils. Pak. J. Bot. 2023, 55, 1. [Google Scholar] [CrossRef]
- Ali, B.; Azeem, M.A.; Qayyum, A.; Mustafa, G.; Ahmad, M.A.; Javed, M.T.; Chaudhary, H.J. Bio-Fabricated Silver Nanoparticles: A Sustainable Approach for Augmentation of Plant Growth and Pathogen Control. In Sustainable Agriculture Reviews; Springer: Cham, Switzerland, 2021; Volume 53, pp. 345–371. [Google Scholar]
- Afridi, M.S.; Javed, M.A.; Ali, S.; De Medeiros, F.H.V.; Ali, B.; Salam, A.; Sumaira Marc, R.A.; Alkhalifah, D.H.M.; Selim, S.; Santoyo, G. New opportunities in plant microbiome engineering for increasing agricultural sustainability under stressful conditions. Front. Plant Sci. 2022, 13, 899464. [Google Scholar] [CrossRef]
- Rohman, M.M.; Ahmed, I.; Molla, M.R.; Hossain, M.A.; Amiruzzaman, M. Evaluation of salt tolerant mungbean (Vigna radiata L.) Genotypes on growth through bio-molecular approaches. Bangladesh J. Agric. Res. 2019, 44, 469–492. [Google Scholar] [CrossRef]
- El-Ramady, H.; Abdalla, N.; Alshaal, T.; El-Henawy, A.; Elmahrouk, M.; Bayoumi, Y.; Domokos-Szabolcsy, É. Plant Nano-nutrition: Perspectives and Challenges. In Nanotechnology, Food Security and Water Treatment; Springer International Publishing: Cham, Switzerland, 2018; pp. 129–161. [Google Scholar]
- Amin, A.A.; El-Kader, A.A.A.; Abouziena, H.F.; El-Awadi, M.; Gharib, F.A. Effects of benzoic acid and thiourea on growth and productivity of wheat (Triticum aestivum L.) plants. Int. J. Sci. Res. 2016, 72, 1032–1037. [Google Scholar]
- Srivastava, A.K.; Sablok, G.; Hackenberg, M.; Deshpande, U.; Suprasanna, P. Thiourea priming enhances salt tolerance through co-ordinated regulation of microRNAs and hormones in Brassica juncea. Sci. Rep. 2017, 7, 45490. [Google Scholar] [CrossRef] [PubMed]
- Torabian, S.; Zahedi, M.; Khoshgoftar, A.H. Effects of foliar spray of nanoparticles of FeSO4 on the growth and ion content of sunflower under saline condition. J. Plant Nutr. 2017, 40, 615–623. [Google Scholar] [CrossRef]
- Wahab, A.; Abdi, G.; Saleem, M.H.; Ali, B.; Ullah, S.; Shah, W.; Mumtaz, S.; Yasin, G.; Muresan, C.C.; Marc, R.A. Plants’ Physio-Biochemical and Phyto-Hormonal Responses to Alleviate the Adverse Effects of Drought Stress: A Comprehensive Review. Plants 2022, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Saleem, K.; Asghar, M.A.; Saleem, M.H.; Raza, A.; Kocsy, G.; Iqbal, N.; Ali, B.; Albeshr, M.F.; Bhat, E.A. Chrysotile-Asbestos-Induced Damage in Panicum virgatum and Phleum pretense Species and Its Alleviation by Organic-Soil Amendment. Sustainability 2022, 14, 10824. [Google Scholar] [CrossRef]
- Ma, J.; Saleem, M.H.; Yasin, G.; Mumtaz, S.; Qureshi, F.F.; Ali, B.; Ercisli, S.; Alhag, S.K.; Ahmed, A.E.; Vodnar, D.C.; et al. Individual and combinatorial effects of SNP and NaHS on morpho-physio-biochemical attributes and phytoextraction of chromium through Cr-stressed spinach (Spinacia oleracea L.). Front. Plant Sci. 2022, 13, 973740. [Google Scholar] [CrossRef]
- Ma, J.; Saleem, M.H.; Ali, B.; Rasheed, R.; Ashraf, M.A.; Aziz, H.; Ercisli, S.; Riaz, S.; Elsharkawy, M.M.; Hussain, I.; et al. Impact of foliar application of syringic acid on tomato (Solanum lycopersicum L.) under heavy metal stress-insights into nutrient uptake, redox homeostasis, oxidative stress, and antioxidant defense. Front. Plant Sci. 2022, 13, 950120. [Google Scholar] [CrossRef]
- Nasrallah, A.K.; Kheder, A.A.; Kord, M.A.; Fouad, A.S.; El-Mogy, M.M.; Atia, M.A.M. Mitigation of Salinity Stress Effects on Broad Bean Productivity Using Calcium Phosphate Nanoparticles Application. Horticulturae 2022, 8, 75–84. [Google Scholar] [CrossRef]
- Biswas, P.; Wu, C.Y. Critical review: Nanoparticles and the environment. J. Air Waste Manag. Assoc. 2005, 55, 708–746. [Google Scholar] [CrossRef]
- Rehman, M.Z.; Rizwan, M.; Sabir, M.; Shahjahan, A.S.; Ahmed, H.R. Comparative effects of different soil conditioners on wheat growth and yield grown in saline-sodic soils. Sains Malays. 2016, 45, 339–346. [Google Scholar]
- Zainab, N.; Khan, A.A.; Azeem, M.A.; Ali, B.; Wang, T.; Shi, F.; Alghanem, S.M.; Hussain Munis, M.F.; Hashem, M.; Alamri, S.; et al. PGPR-Mediated Plant Growth Attributes and Metal Extraction Ability of Sesbania sesban L. in Industrially Contaminated Soils. Agronomy 2021, 11, 1820. [Google Scholar] [CrossRef]
- Claussen, W. Proline as a measure of stress in tomato plants. Plant Sci. 2005, 168, 241–248. [Google Scholar] [CrossRef]
- Jain, M.; Mathur, G.; Koul, S.; Sarin, N.B. Ameliorative effect of praline on salt stress induced lipid peroxidation in cell lines of groundnut (Arachis hypogea L.). Plant Cell Rep. 2001, 20, 463–468. [Google Scholar] [CrossRef]
- Jeevanandam, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: History, sources, toxicity and regulations. Belistein J. Nanotechnol. 2018, 9, 1050–1074. [Google Scholar] [CrossRef]
- Paz, A.; Guadarrama, D.; Lopez, M.; Gonzalez, J.E.; Brizuela, N.; Aragon, J. A comparative study of Hydroxyapaptite nanoparticles synthesized by different routes. Quim. Nova 2012, 35, 1724–1727. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids-Extraction, Isolation and purification. In Current Protocols in Food and Analytical Chemistry (CPFA); John Wiley: New York, NY, USA, 2001; Volume 9. [Google Scholar]
- Zhang, Y.; He, Z.H.; Zhou, G.Y.; Wang, D.S. Genotype and environment effects on major quality characters of wintersown Chinese wheats. J. Chin. Cer. Oils Assoc. 1999, 4, 1–5. [Google Scholar]
- Zhang, X.Z. Crop Physiology Research Methods; China Agriculture Press: Beijing, China, 1992; pp. 148–149. [Google Scholar]
- Verma, S.; Dubey, R.S. Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci. 2003, 164, 645–655. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Leaf Length (cm) | Leaf Width (cm) | Leaf Fresh wt. (gm) | Leaf Dry wt. (gm) | Leaf Area (cm2) | Shoot Length (cm) | Shoot Fresh wt. (gm) | Shoot Dry wt. (gm) | Root Length (cm) | Root Fresh wt. (gm) | Root Dry wt. (gm) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | 38 b | 2.3 cd | 1.9 | 0.22 | 42.10 ab | 8.5 a | 27.13 bc | 0.75 | 6.2 bc | 7.5 abc | 0.70 |
| T2 | 35 cd | 1.9 d | 1.4 | 0.20 | 41.89 bc | 8.5 a | 23.44 de | 0.73 | 6.0 cd | 7.2 bcd | 0.69 |
| T3 | 31 d | 1.8 d | 1.3 | 0.20 | 40.54 bc | 6.1 bc | 20.29 e | 0.70 | 5.5 d | 7.0 bcd | 0.68 |
| T4 | 41 b | 3.2 abc | 2.0 | 0.25 | 41.54 bc | 5.5 bc | 28.07 ab | 0.74 | 6.8 cb | 7.7 ab | 0.71 |
| T5 | 40 bc | 2.7 bc | 1.8 | 0.21 | 40.64 bc | 5.0 c | 26.92 bc | 0.70 | 6.0 c | 7.1 bc | 0.70 |
| T6 | 39 bc | 2.3 cd | 1.4 | 0.20 | 40.55 bc | 4.9 c | 21.47 de | 0.66 | 5.8 cd | 7.0 bc | 0.69 |
| T7 | 40 bc | 3.5 ab | 2.2 | 0.26 | 43.42 a | 7.6 ab | 28.86 a | 0.79 | 7.0 a | 7.9 ab | 0.74 |
| T8 | 39 bc | 2.6 bc | 1.9 | 0.25 | 42.78 ab | 8.7 a | 27.20 ab | 0.76 | 6.8 ab | 7.6 abc | 0.69 |
| T9 | 31 d | 1.9 d | 1.5 | 0.22 | 40.87 bc | 7.1 ab | 25.11 cd | 0.60 | 6.1 cd | 6.9 cd | 0.60 |
| T10 | 51 a | 3.8 a | 2.5 | 0.25 | 43.98 a | 6.4 bc | 29.79 a | 0.80 | 7.1 a | 8.0 a | 0.73 |
| T11 | 50 a | 2.8 bc | 2.0 | 0.19 | 40.00 bc | 7.0 ab | 25.37 cd | 0.60 | 6.2 bc | 7.6 ab | 0.69 |
| T12 | 49 a | 1.6 d | 1.2 | 0.17 | 39.98 bc | 6.1 bc | 23.25 d | 0.59 | 6.0 cd | 6.5 d | 0.59 |
| LSD | 5.68 | 1.12 | NS | NS | 2.25 | 2.32 | 2.61 | NS | 0.65 | 0.80 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faryal, S.; Ullah, R.; Khan, M.N.; Ali, B.; Hafeez, A.; Jaremko, M.; Qureshi, K.A. Thiourea-Capped Nanoapatites Amplify Osmotic Stress Tolerance in Zea mays L. by Conserving Photosynthetic Pigments, Osmolytes Biosynthesis and Antioxidant Biosystems. Molecules 2022, 27, 5744. https://doi.org/10.3390/molecules27185744
Faryal S, Ullah R, Khan MN, Ali B, Hafeez A, Jaremko M, Qureshi KA. Thiourea-Capped Nanoapatites Amplify Osmotic Stress Tolerance in Zea mays L. by Conserving Photosynthetic Pigments, Osmolytes Biosynthesis and Antioxidant Biosystems. Molecules. 2022; 27(18):5744. https://doi.org/10.3390/molecules27185744
Chicago/Turabian StyleFaryal, Sana, Rehman Ullah, Muhammad Nauman Khan, Baber Ali, Aqsa Hafeez, Mariusz Jaremko, and Kamal Ahmad Qureshi. 2022. "Thiourea-Capped Nanoapatites Amplify Osmotic Stress Tolerance in Zea mays L. by Conserving Photosynthetic Pigments, Osmolytes Biosynthesis and Antioxidant Biosystems" Molecules 27, no. 18: 5744. https://doi.org/10.3390/molecules27185744
APA StyleFaryal, S., Ullah, R., Khan, M. N., Ali, B., Hafeez, A., Jaremko, M., & Qureshi, K. A. (2022). Thiourea-Capped Nanoapatites Amplify Osmotic Stress Tolerance in Zea mays L. by Conserving Photosynthetic Pigments, Osmolytes Biosynthesis and Antioxidant Biosystems. Molecules, 27(18), 5744. https://doi.org/10.3390/molecules27185744