
Recent Updates on the Functional Impact of Kahweol and Cafestol on Cancer

 , , and
, , and
Abstract
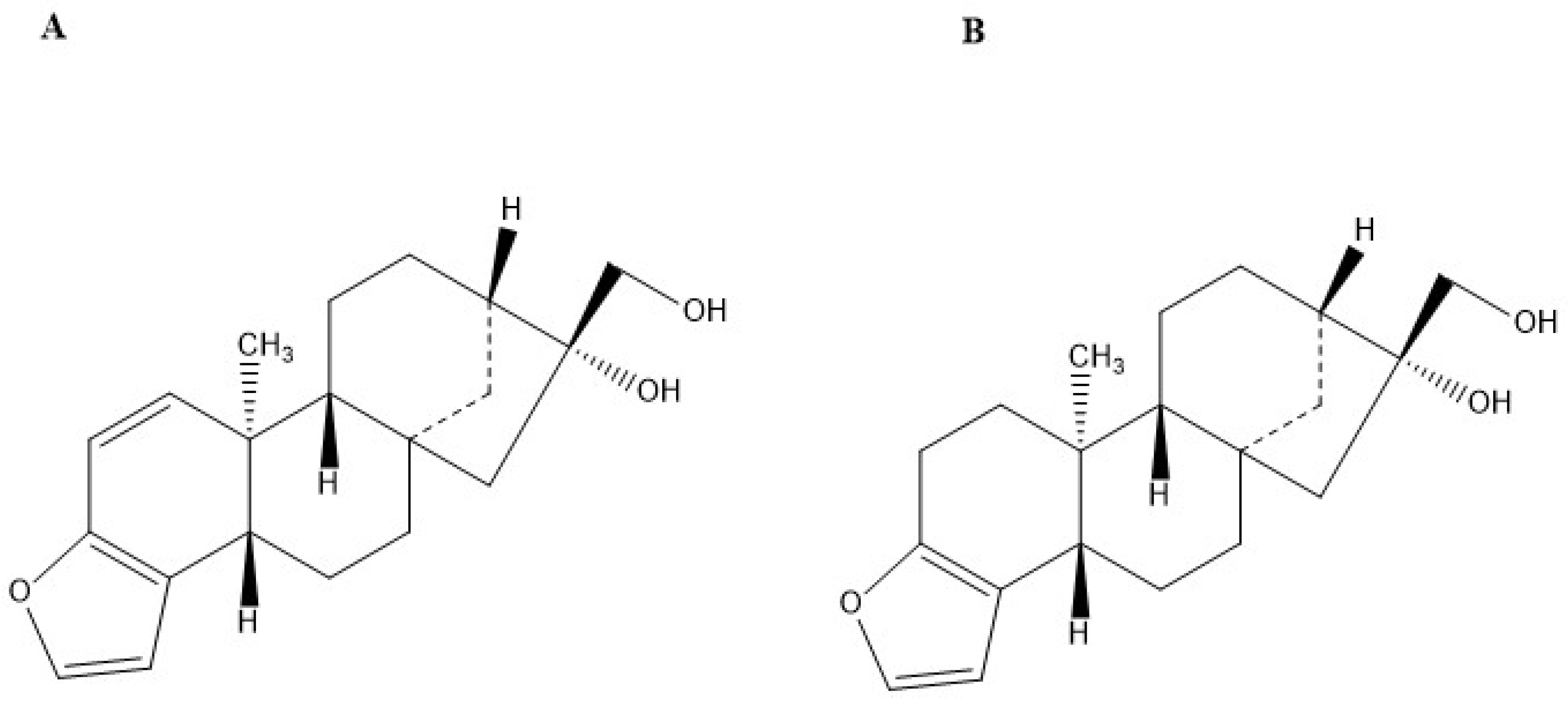
:1. Introduction
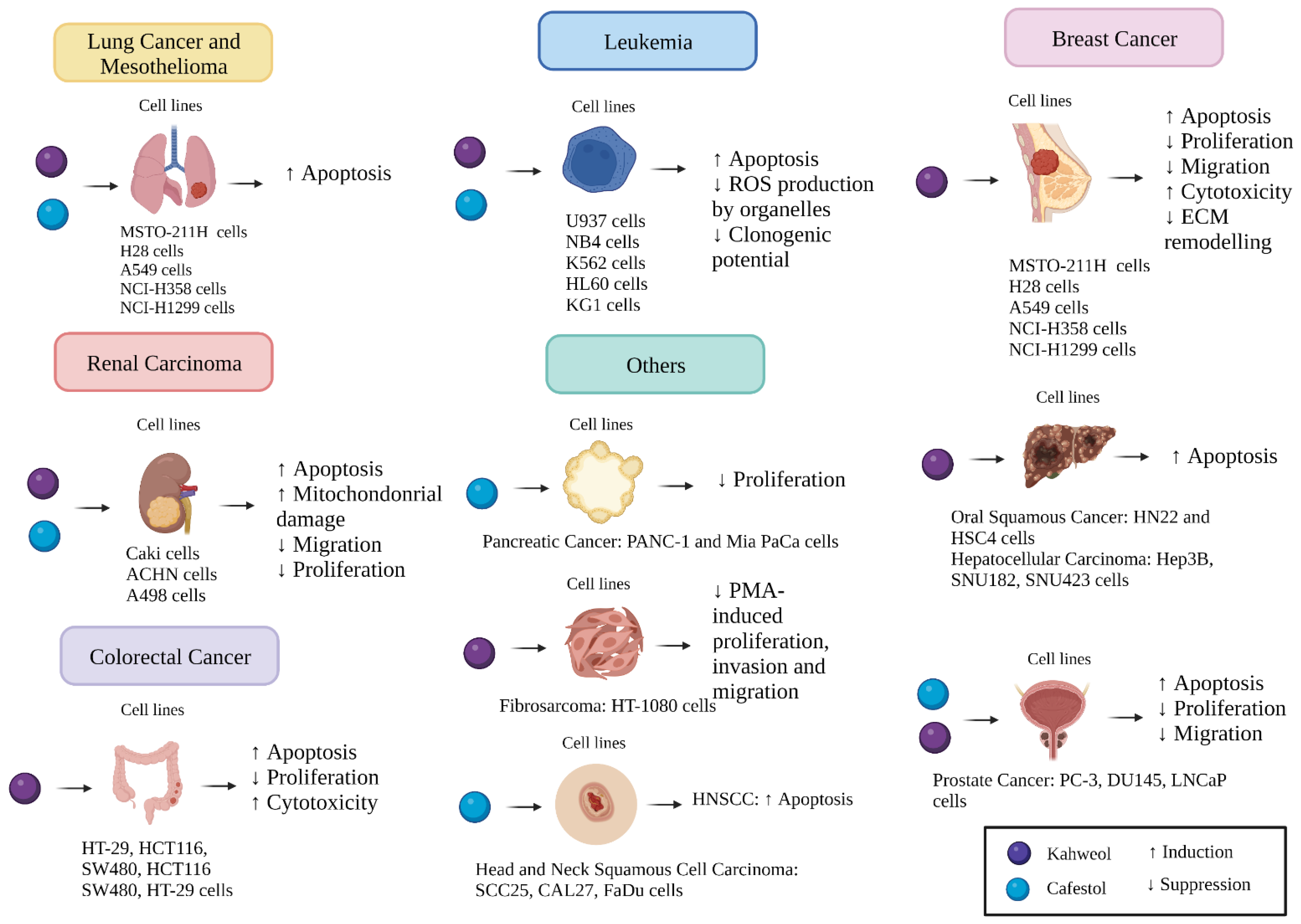
2. Kahweol and Cafestol Effects on Several Cancer Cell Lines
2.1. Lung Cancer
2.2. Oral Squamous Cancer
2.3. Prostate Cancer
2.4. Breast Cancer
2.5. Colorectal Cancer
2.6. Renal Carcinoma
2.7. Leukaemia
2.8. Fibrosarcoma
2.9. Hepatocellular Carcinoma
2.10. Head and Neck Squamous Cell Carcinoma
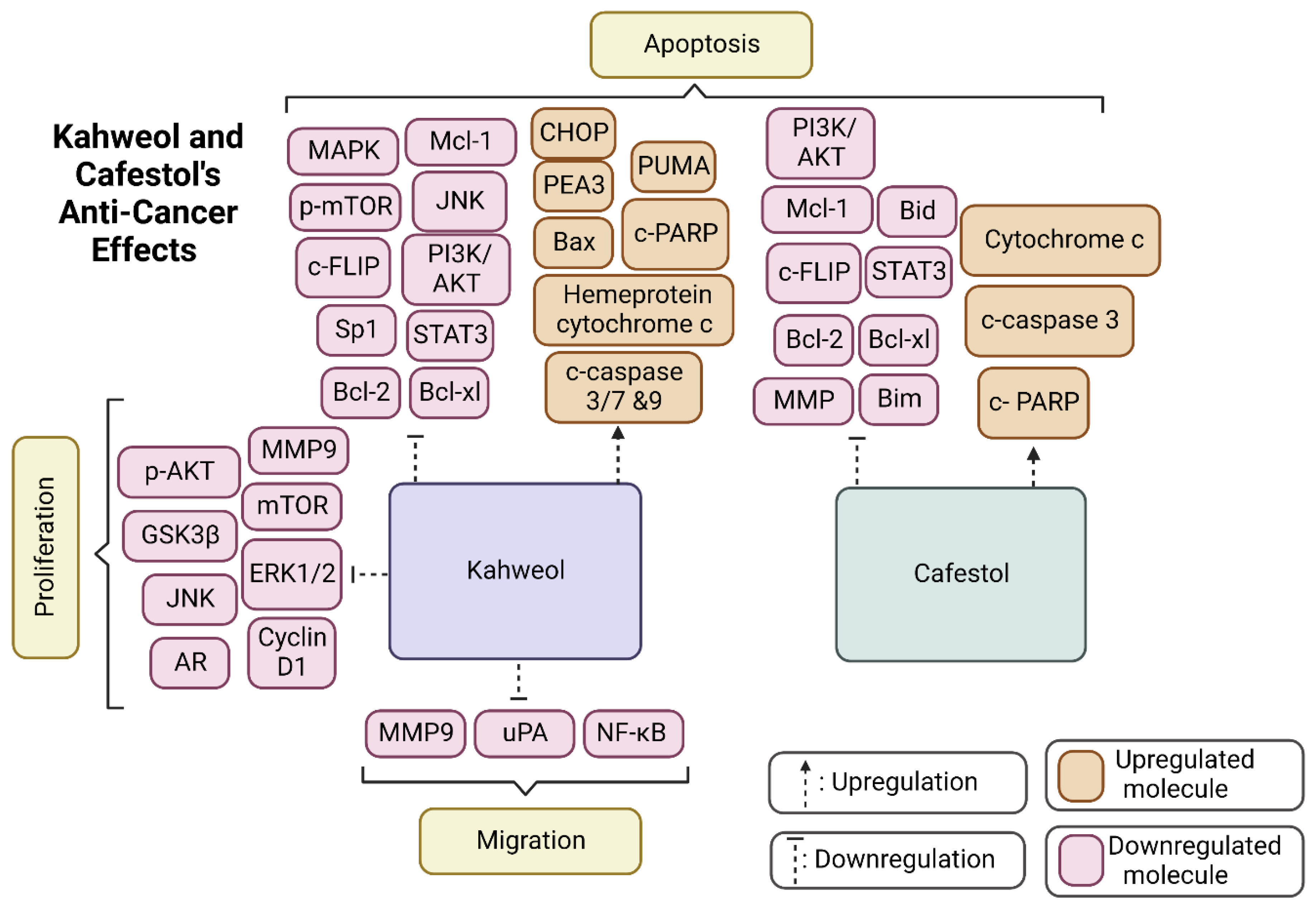
3. Conclusions and Future Perspectives
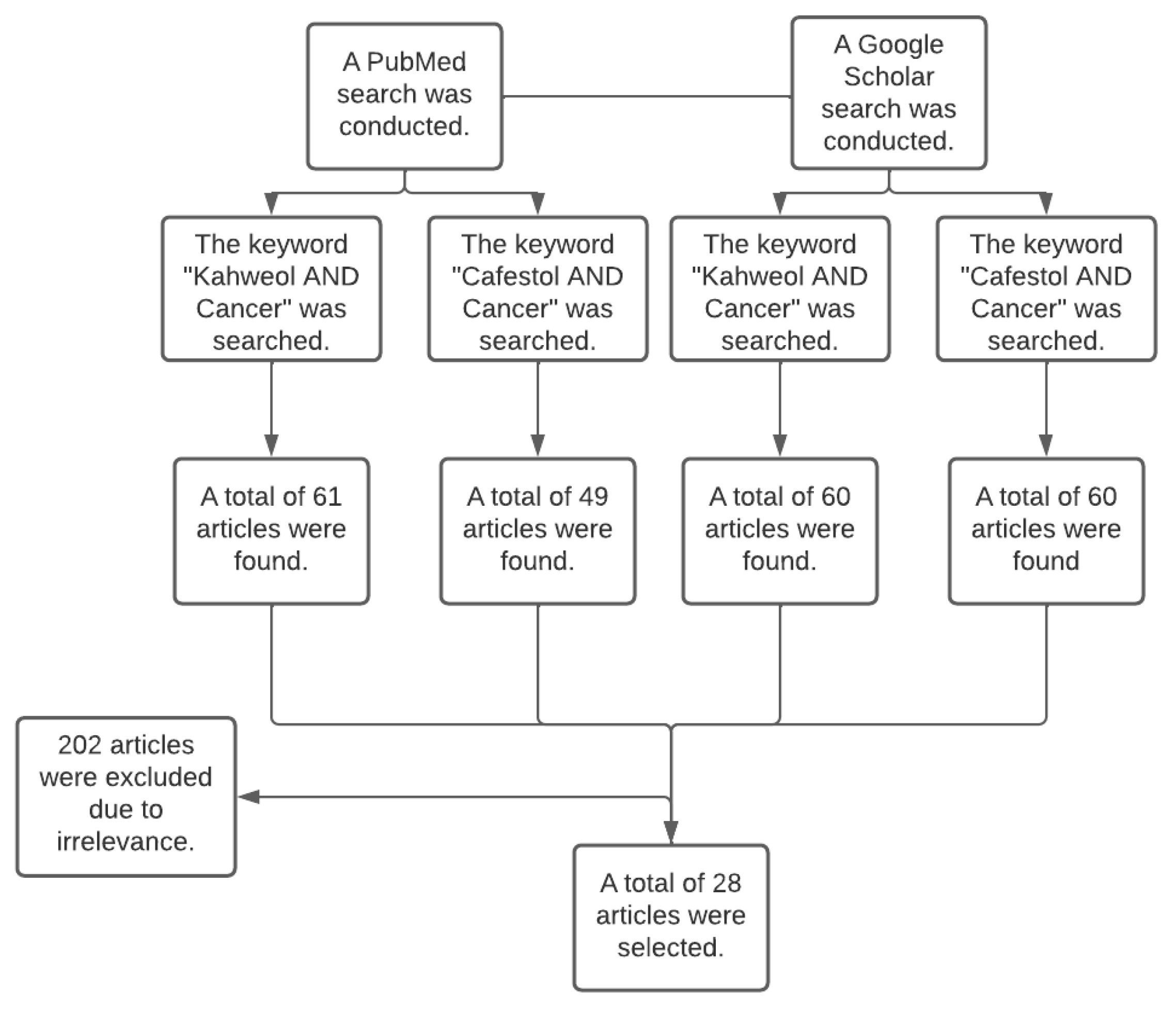
4. Methods
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AP-2 | Activator Protein 2 |
| AR | Androgen Receptor |
| ATF3 | Cyclic AMP-dependent transcription factor |
| BALB/c-nude mice | Bagg and Albino/c-nude mice |
| Bax | BCL2-Associated X Protein |
| Bcl-2 | B-cell leukemia/lymphoma 2 protein. |
| Bcl-xL | B-cell lymphoma-extra large |
| Bid | BH3 interacting-domain death agonist |
| Bim | Bcl-2-like protein 11 |
| BTF3 | Basic transcription factor 3 |
| CCL2-CCR2 | Chemokine (C-C motif) ligand 2-CC chemokine receptor 2 |
| CCL5-CCR5 | Chemokine (C-C motif) ligand 5-CC chemokine receptor 5 |
| CCR6 | CC chemokine receptor 6 |
| CD11b | Cluster of differentiation 11b |
| CD15 | Cluster of differentiation 15 |
| cFLIP | Cellular FLICE (FADD-like IL-1β-converting enzyme)-inhibitory protein |
| CHOP | C/EBP Homologous Protein |
| DEVDase | Caspase-3-like proteases |
| ECM | Extra-cellular Matrix |
| ER | Endoplasmic Reticulum |
| ERK signaling | Extracellular signal-regulated kinase |
| FASN | Fatty Acid Synthase |
| FDA | Food and Drug administration |
| GSK-3β | Glycogen synthase kinase-3 beta |
| HER2 | Human epidermal growth factor receptor 2 |
| HMVECs | Human microvascular endothelial cell-1 |
| HSP40/70/90 | Human Heat shock protein 40/70/90 |
| ICAM1 | Intercellular adhesion molecule 1 |
| IL-8 | Interleukin-8 |
| JNK | Jun N-terminal kinase |
| LDL | Low-density Lipoprotein |
| MAPK | Mitogen-activated protein kinase |
| mcl1 | Myeloid cell leukemia-1 |
| MCP1 | Monocyte chemoattractant protein-1 |
| MMP-9 | Matrix metallopeptidase 9 |
| mTOR | Mammalian target of rapamycin |
| NK-kB | nNuclear factor kappa light chain enhancer of activated B cells |
| p-4EBP1 | Phosphorylated eukaryotic translation initiation factor |
| p-AKT | Phosphorylated Protein kinase B |
| PARP | Poly-ADP ribose polymerase |
| PD-L1 | Programmed death-ligand 1 |
| PEA3 | Polyoma enhancer activator 3 |
| PI3K/Akt pathway | Phosphatidylinositol 3-kinase/Protein kinase B |
| PMA | Phorbol 12-myristate 13-acetate |
| p-mTOR | Phosphorylated mammalian target of rapamycin |
| p-p70S6K | Phosphorylated Ribosomal protein S6 kinase beta-1 |
| p-Src | Phosphorilated sarcoma |
| p-STAT3 | Phosphorylated signal transducer and activator of transcription 3 |
| PUMA | p53-upregulated modulator of apoptosis |
| ROS | Reactive oxygen species |
| SCID mice | Severe combined immunodeficiency mice |
| Sp1 | Specificity protein 1 |
| SREBP-1c | Sterol regulatory element-binding protein-1 |
| STAT3 | Signal transducer and activator of transcription 3 |
| Thr286 | Threonine 286 |
| TRAIL-Mediated apoptosis | Tumor necrosis factor-related apoptosis-inducing ligand |
| uPA | Urokinase-type plasminogen activator (uPA) |
| XIAP | X-linked inhibitor of apoptosis protein |
References
- Hong, C.T.; Chan, L.; Bai, C.H. The Effect of Caffeine on the Risk and Progression of Parkinson’s Disease: A Meta-Analysis. Nutrients 2020, 12, 1860. [Google Scholar] [CrossRef]
- Gokcen, B.B.; Sanlier, N. Coffee consumption and disease correlations. Crit. Rev. Food Sci. Nutr. 2019, 59, 336–348. [Google Scholar] [CrossRef]
- Feld, J.J.; Lavoie, E.G.; Fausther, M.; Dranoff, J.A. I drink for my liver, Doc: Emerging evidence that coffee prevents cirrhosis. F1000Research 2015, 4, 95. [Google Scholar] [CrossRef]
- Cano-Marquina, A.; Tarin, J.J.; Cano, A. The impact of coffee on health. Maturitas 2013, 75, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Camandola, S.; Plick, N.; Mattson, M.P. Impact of Coffee and Cacao Purine Metabolites on Neuroplasticity and Neurodegenerative Disease. Neurochem. Res. 2019, 44, 214–227. [Google Scholar] [CrossRef] [PubMed]
- El-Huneidi, W.; Anjum, S.; Bajbouj, K.; Abu-Gharbieh, E.; Taneera, J. The Coffee Diterpene, Kahweol, Ameliorates Pancreatic beta-Cell Function in Streptozotocin (STZ)-Treated Rat INS-1 Cells through NF-kB and p-AKT/Bcl-2 Pathways. Molecules 2021, 26, 5167. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.A.; Chae, J.I.; Shim, J.H. Natural diterpenes from coffee, cafestol and kahweol induce apoptosis through regulation of specificity protein 1 expression in human malignant pleural mesothelioma. J. Biomed. Sci. 2012, 19, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.; Wang, C.; Xu, J.; Wang, S. Cafestol and Kahweol: A Review on Their Bioactivities and Pharmacological Properties. Int. J. Mol. Sci. 2019, 20, 4238. [Google Scholar] [CrossRef] [Green Version]
- Dias, R.C.; Faria, A.F.D.; Mercadante, A.Z.; Bragagnolo, N.; Benassi, M.D.T. Comparison of extraction methods for kahweol and cafestol analysis in roasted coffee. J. Braz. Chem. Soc. 2013, 24, 492–499. [Google Scholar] [CrossRef] [Green Version]
- Novaes, F.J.M.; Junior, I.I.; Sutili, F.K.; Marriott, P.J.; Bizzo, H.R.; de Aquino Neto, F.R.; de Souza, R.O.M.A.; Rezende, C.M. Lipase-catalysed esters synthesis of cafestol and kahweol. Food Chem. 2018, 259, 226–233. [Google Scholar] [CrossRef] [PubMed]
- De Roos, B.; Meyboom, S.; Kosmeijer-Schuil, T.G.; Katan, M.B. Absorption and urinary excretion of the coffee diterpenes cafestol and kahweol in healthy ileostomy volunteers. J. Intern. Med. 1998, 244, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Cruchten, S.T.; de Waart, D.R.; Kunne, C.; Hooiveld, G.J.; Boekschoten, M.V.; Katan, M.B.; Elferink, R.P.; Witkamp, R.F. Absorption, distribution, and biliary excretion of cafestol, a potent cholesterol-elevating compound in unfiltered coffees, in mice. Drug Metab. Dispos. 2010, 38, 635–640. [Google Scholar] [CrossRef] [Green Version]
- De Roos, B.; Van Tol, A.; Urgert, R.; Scheek, L.M.; Van Gent, T.; Buytenhek, R.; Princen, H.M.; Katan, M.B. Consumption of French-press coffee raises cholesteryl ester transfer protein activity levels before LDL cholesterol in normolipidaemic subjects. J. Intern. Med. 2000, 248, 211–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halvorsen, B.; Ranheim, T.; Nenseter, M.S.; Huggett, A.C.; Drevon, C.A. Effect of a coffee lipid (cafestol) on cholesterol metabolism in human skin fibroblasts. J. Lipid Res. 1998, 39, 901–912. [Google Scholar] [CrossRef]
- Post, S.M.; de Wit, E.C.; Princen, H.M. Cafestol, the cholesterol-raising factor in boiled coffee, suppresses bile acid synthesis by downregulation of cholesterol 7 alpha-hydroxylase and sterol 27-hydroxylase in rat hepatocytes. Arter. Thromb. Vasc. Biol. 1997, 17, 3064–3070. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.R.; Sung, L.C.; Chen, C.C.; Hong, H.J.; Liu, J.C.; Chen, J.J. Cafestol Activates Nuclear Factor Erythroid-2 Related Factor 2 and Inhibits Urotensin II-Induced Cardiomyocyte Hypertrophy. Am. J. Chin. Med. 2019, 47, 337–350. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.R.; Sung, L.C.; Chen, C.C.; Chen, P.Y.; Cheng, T.H.; Chao, H.H.; Liu, J.C.; Chen, J.J. Corrigendum to “Cafestol Inhibits Cyclic-Strain-Induced Interleukin-8, Intercellular Adhesion Molecule-1, and Monocyte Chemoattractant Protein-1 Production in Vascular Endothelial Cells”. Oxid. Med. Cell. Longev. 2019, 2019, 1536397. [Google Scholar] [CrossRef] [PubMed]
- Fumimoto, R.; Sakai, E.; Yamaguchi, Y.; Sakamoto, H.; Fukuma, Y.; Nishishita, K.; Okamoto, K.; Tsukuba, T. The coffee diterpene kahweol prevents osteoclastogenesis via impairment of NFATc1 expression and blocking of Erk phosphorylation. J. Pharmacol. Sci. 2012, 118, 479–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuma, Y.; Sakai, E.; Nishishita, K.; Okamoto, K.; Tsukuba, T. Cafestol has a weaker inhibitory effect on osteoclastogenesis than kahweol and promotes osteoblast differentiation. Biofactors 2015, 41, 222–231. [Google Scholar] [CrossRef]
- Choi, M.J.; Park, E.J.; Oh, J.H.; Min, K.J.; Yang, E.S.; Kim, Y.H.; Lee, T.J.; Kim, S.H.; Choi, Y.H.; Park, J.W.; et al. Cafestol, a coffee-specific diterpene, induces apoptosis in renal carcinoma Caki cells through down-regulation of anti-apoptotic proteins and Akt phosphorylation. Chem. Biol. Interact. 2011, 190, 102–108. [Google Scholar] [CrossRef]
- Moeenfard, M.; Cortez, A.; Machado, V.; Costa, R.; Luis, C.; Coelho, P.; Soares, R.; Alves, A.; Borges, N.; Santos, A. Anti-Angiogenic Properties of Cafestol and Kahweol Palmitate Diterpene Esters. J. Cell. Biochem. 2016, 117, 2748–2756. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.G.; Hwang, Y.P.; Jeong, H.G. Kahweol blocks STAT3 phosphorylation and induces apoptosis in human lung adenocarcinoma A549 cells. Toxicol. Lett. 2009, 187, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Bang, W.; Cho, J.H.; Lee, R.H.; Kim, S.H.; Kim, M.S.; Park, S.M.; Shin, J.C.; Chung, H.J.; Oh, K.B.; et al. Kahweol induces apoptosis by suppressing BTF3 expression through the ERK signaling pathway in non-small cell lung cancer cells. Int. J. Oncol. 2016, 49, 2294–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chae, J.I.; Jeon, Y.J.; Shim, J.H. Anti-proliferative properties of kahweol in oral squamous cancer through the regulation specificity protein 1. Phytother. Res. 2014, 28, 1879–1886. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, H.; Izumi, K.; Natsagdorj, A.; Naito, R.; Makino, T.; Kadomoto, S.; Hiratsuka, K.; Shigehara, K.; Kadono, Y.; Narimoto, K.; et al. Coffee diterpenes kahweol acetate and cafestol synergistically inhibit the proliferation and migration of prostate cancer cells. Prostate 2019, 79, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, C.; Quesada, A.R.; Medina, M.A. Insights on the antitumor effects of kahweol on human breast cancer: Decreased survival and increased production of reactive oxygen species and cytotoxicity. Biochem. Biophys. Res. Commun. 2014, 447, 452–458. [Google Scholar] [CrossRef]
- Oh, S.H.; Hwang, Y.P.; Choi, J.H.; Jin, S.W.; Lee, G.H.; Han, E.H.; Chung, Y.H.; Chung, Y.C.; Jeong, H.G. Kahweol inhibits proliferation and induces apoptosis by suppressing fatty acid synthase in HER2-overexpressing cancer cells. Food Chem. Toxicol. 2018, 121, 326–335. [Google Scholar] [CrossRef]
- Park, G.H.; Song, H.M.; Jeong, J.B. Kahweol from Coffee Induces Apoptosis by Upregulating Activating Transcription Factor 3 in Human Colorectal Cancer Cells. Biomol. Ther. 2017, 25, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Park, G.H.; Song, H.M.; Jeong, J.B. The coffee diterpene kahweol suppresses the cell proliferation by inducing cyclin D1 proteasomal degradation via ERK1/2, JNK and GKS3beta-dependent threonine-286 phosphorylation in human colorectal cancer cells. Food Chem. Toxicol. 2016, 95, 142–148. [Google Scholar] [CrossRef]
- Choi, J.H.; Hwang, Y.P.; Jin, S.W.; Lee, G.H.; Kim, H.G.; Han, E.H.; Kim, S.K.; Kang, K.W.; Chung, Y.C.; Jeong, H.G. Suppression of PMA-induced human fibrosarcoma HT-1080 invasion and metastasis by kahweol via inhibiting Akt/JNK1/2/p38 MAPK signal pathway and NF-kappaB dependent transcriptional activities. Food Chem. Toxicol. 2019, 125, 1–9. [Google Scholar] [CrossRef]
- Choi, D.W.; Lim, M.S.; Lee, J.W.; Chun, W.; Lee, S.H.; Nam, Y.H.; Park, J.M.; Choi, D.H.; Kang, C.D.; Lee, S.J.; et al. The Cytotoxicity of Kahweol in HT-29 Human Colorectal Cancer Cells Is Mediated by Apoptosis and Suppression of Heat Shock Protein 70 Expression. Biomol. Ther. 2015, 23, 128–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Um, H.J.; Oh, J.H.; Kim, Y.N.; Choi, Y.H.; Kim, S.H.; Park, J.W.; Kwon, T.K. The coffee diterpene kahweol sensitizes TRAIL-induced apoptosis in renal carcinoma Caki cells through down-regulation of Bcl-2 and c-FLIP. Chem. Biol. Interact. 2010, 186, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Um, H.J.; Park, J.W.; Kwon, T.K. Melatonin sensitizes Caki renal cancer cells to kahweol-induced apoptosis through CHOP-mediated up-regulation of PUMA. J. Pineal. Res. 2011, 50, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Min, K.J.; Um, H.J.; Kim, J.I.; Kwon, T.K. The coffee diterpene kahweol enhances sensitivity to sorafenib in human renal carcinoma Caki cells through down-regulation of Mcl-1 and c-FLIP expression. Oncotarget 2017, 8, 83195–83206. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Izumi, K.; Hiratsuka, K.; Kano, H.; Shimada, T.; Nakano, T.; Kadomoto, S.; Naito, R.; Iwamoto, H.; Yaegashi, H.; et al. Anti-proliferative and anti-migratory properties of coffee diterpenes kahweol acetate and cafestol in human renal cancer cells. Sci. Rep. 2021, 11, 675. [Google Scholar] [CrossRef]
- Woo, S.M.; Min, K.J.; Seo, B.R.; Nam, J.O.; Choi, K.S.; Yoo, Y.H.; Kwon, T.K. Cafestol overcomes ABT-737 resistance in Mcl-1-overexpressed renal carcinoma Caki cells through downregulation of Mcl-1 expression and upregulation of Bim expression. Cell Death Dis. 2014, 5, e1514. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.Y.; Lee, S.H.; Lee, J.H.; Lee, J.H.; Jang, B.K.; Kim, M.K. Kahweol Induces Apoptosis in Hepatocellular Carcinoma Cells by Inhibiting the Src/mTOR/STAT3 Signaling Pathway. Int. J. Mol. Sci. 2021, 22, 10509. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Lee, J.T.; Yang, E.S.; Chang, J.S.; Lee, D.S.; Kim, S.H.; Choi, Y.H.; Park, J.W.; Kwon, T.K. The coffee diterpene kahweol induces apoptosis in human leukemia U937 cells through down-regulation of Akt phosphorylation and activation of JNK. Apoptosis 2009, 14, 1378–1386. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.S.; Spindola, D.G.; Bechara, A.; Garcia, D.M.; Palmeira-Dos-Santos, C.; Peixoto-da-Silva, J.; Erustes, A.G.; Michelin, L.F.G.; Pereira, G.J.S.; Smaili, S.S.; et al. Cafestol, a diterpene molecule found in coffee, induces leukemia cell death. Biomed. Pharmacother. 2017, 92, 1045–1054. [Google Scholar] [CrossRef]
- Saito, T.; Abe, D.; Nogata, Y.J.F.S. Coffee diterpenes potentiate the cytolytic activity of KHYG-1 NK Leukemia cells. Food Sci. Technol. Res. 2015, 21, 281–284. [Google Scholar] [CrossRef]
- Kotowski, U.; Heiduschka, G.; Seemann, R.; Eckl-Dorna, J.; Schmid, R.; Kranebitter, V.; Stanisz, I.; Brunner, M.; Lill, C.; Thurnher, D. Effect of the coffee ingredient cafestol on head and neck squamous cell carcinoma cell lines. Strahlenther. Onkol. 2015, 191, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef] [PubMed]
- Noel, P.; Von Hoff, D.D.; Saluja, A.K.; Velagapudi, M.; Borazanci, E.; Han, H. Triptolide and Its Derivatives as Cancer Therapies. Trends Pharmacol. Sci. 2019, 40, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.P.; Tan, Z.J.; Wu, X.S.; Liu, T.Y.; Jiang, L.; Bao, R.F.; Shu, Y.J.; Li, M.L.; Weng, H.; Ding, Q.; et al. Triptolide induces s phase arrest and apoptosis in gallbladder cancer cells. Molecules 2014, 19, 2612–2628. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Jung, K.S.; Jeong, H.G. Suppressive effects of the kahweol and cafestol on cyclooxygenase-2 expression in macrophages. FEBS Lett. 2004, 569, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.J.; Choi, J.H.; Jeong, H.G. Hepatoprotective and antioxidant effects of the coffee diterpenes kahweol and cafestol on carbon tetrachloride-induced liver damage in mice. Food Chem. Toxicol. 2007, 45, 2118–2125. [Google Scholar] [CrossRef]
- Parker, G.L.; Smith, L.K.; Baxendale, I.R. Development of the industrial synthesis of vitamin A. Tetrahedron 2016, 72, 1645–1652. [Google Scholar] [CrossRef]
- Yoham, A.L.; Casadesus, D. Tretinoin. In StatPearls [Internet]; StatPearls Publishing: Tampa, FL, USA, 2022. [Google Scholar]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Shaw, S.; Islam, M.A.; Ahmed, M.I.; Chandra Shill, M.; Karmakar, U.K.; Yarla, N.S.; Khan, I.N.; et al. Phytol: A review of biomedical activities. Food Chem. Toxicol. 2018, 121, 82–94. [Google Scholar] [CrossRef]
- Schwalfenberg, G.K. Vitamins K1 and K2: The Emerging Group of Vitamins Required for Human Health. J. Nutr. Metab. 2017, 2017, 6254836. [Google Scholar] [CrossRef]
- Daines, A.M.; Payne, R.J.; Humphries, M.E.; Abell, A.D. The synthesis of naturally occurring vitamin K and vitamin K analogues. Curr. Org. Chem. 2003, 7, 1625–1634. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Cell Line | Coffee Derivative | Molecular Target | Functional Impact | Ref. | |
|---|---|---|---|---|---|---|
| Mesothelioma | MSTO-211H cells H28 cells | Kahweol and cafestol | ↑ Bax ↓ Bcl-xl ↑ Cleavage of Bid, Caspase-3, and PARP ↓ Sp1 | ↑ Kahweol-induced apoptosis | [7] | |
| Lung Adenocarcinoma | A549 cells | Kahweol | ↑ Bax ↓ Bcl-2 ↓ Bcl-xl ↑ Cleavage of Caspase-3 and PARP ↓ STAT3 | ↑ DNA fragmentation ↑ Caspase-3-mediated apoptosis ↑ STAT3-mediated apoptosis | [22] | |
| Non-small Cell Lung Cancer | NCI-H358 cells NCI-H1299 cells | Kahweol | ↓ BTF3 ↓ ERK signalling pathway ↑ Cleavage of PARP and Caspase-3 ↑ p27 and p21 ↓ cyclin D1 ↑ Bax ↓ Bcl-2 ↓ Bcl-xl | ↑ Kahweol-induced apoptosis | [23] | |
| Oral Squamous Cancer | HN22 cells HSC4 cells | Kahweol | ↓ Sp1 ↑ p27 and p21 ↓ cyclin D1, Mcl-1, and survivin ↑ Cleavage of Bid, Caspase-3, and PARP ↓ Bcl-xl ↑ Bax | ↑ Kahweol-induced apoptosis | [24] | |
| Prostate Cancer | PC-3 DU145 LNCaP | Kahweol acetate Cafestol | ↑ Caspase-3 cleavage ↑ PARP cleavage ↓ Bcl-2 ↓ Bcl-xL ↓ AR ↓ CCL2-CCR2 ↓ CCL5-CCR5 | ↓ Proliferation ↓ Migration ↑ Apoptosis | [25] | |
| Breast Cancer | MDA-MB231 ZR75-1 MCF-7 | Kahweol | ↑ Caspases-3/7, 9 ↑ Cytochrome C ↑ H2O2 | ↓ Proliferation ↑ Apoptosis ↑ H2O2 cytotoxicity | [26] | |
| MDA-MB231 | Kahweol | ↑ Caspases-3/7, 9 ↑ Cytochrome C ↑ p-AKT ↑ ERK ↓ MMP-9 ↓ uPA | ↑ Apoptosis ↓ Migration ↓ ECM remodelling | [26] | ||
| SKBR3MCF-10A | Kahweol | ↑ PARP cleavage via ↑ caspase 3 ↓ HER2 via ↑ PEA3 and ↓ AP-2 ↓ FASN via ↓ SREBP-1c, ↓ p-Akt ↓ cyclin D1 via ↓ mTOR, ↓ GSK-3β | ↑ Apoptosis ↑ Cytotoxicity ↓ Proliferation | [27] | ||
| Colorectal Cancer | HCT116 SW480 LoVo HT-29 | Kahweol | ↑ PARP cleavage via ↑ ATF3 | ↑ Apoptosis | [28] | |
| HCT116 SW480 | Kahweol | ↓ cyclin D1 via ↑ Thr286 | ↓ Proliferation | [29] | ||
| HT-29 | Kahweol | ↑ Caspase-3 cleavage ↑ PARP cleavage ↓ Bcl-2 ↓ p-AKT ↓ HSP40, HSP70, HSP90 | ↑ Apoptosis ↓ Proliferation | [20,30] | ||
| HT-29 | Kahweol | ↑ Caspase-3 ↑ PARP cleavage ↓ Bcl-2 ↓ p-AKT ↓ HSP-70 | ↑ Apoptosis ↑ Cytotoxicity | [31] | ||
| Renal Carcinoma | Caki Cells | Kahweol | ↓ Bcl-2 ↓ c-FLIP ↑ Cleavage of PARP ↑ DEVDase | ↑ TRAIL-mediated apoptosis | [32] | |
| Caki Cells | Kahweol | ↑ PUMA via ↑ CHOP ↑ DEVDase | ↑ p53-independent apoptosis ↑ ER stress-mediated apoptosis | [33] | ||
| Caki cells ACHN cells A498 cells | Kahweol | ↓ Mcl-1 ↓ c-FLIP | ↑ Caspase-mediated apoptosis | [34] | ||
| Caki cells | Cafestol | ↓ Mcl-1 ↓ c-FLIP ↓ MMP ↑ Cytochrome C ↑ Caspase-3 ↓ Bcl-2, Bcl-xL, Mcl-1, c-FLIP ↓ PI3K/Akt pathway | ↑ Mitochondrial damage ↑ Apoptosis | [20] | ||
| Caki-1 cells ACHN cells | Kahweol acetate and cafestol | ↓ Akt and ERK phosphorylation ↓ CCR2, CCR5 & CCR6↓ PD-L1 | ↓ Migration ↓ Proliferation ↓ Epithelial-mesenchymal transition ↑ Apoptosis | [35] | ||
| Caki cells | Cafestol | ↑ Cleavage of PARP ↑ Caspase-3 activity ↓ Mcl-1 ↑ PUMA and Bim | ↑ ABT-737-mediated apoptosis | [36] | ||
| Hepatocellular Carcinoma | Hep3B cells SNU182 cells SNU423 cells | Kahweol | ↑ Cleavage of PARP and caspase 3 ↓ p-Src ↓ expression of p-Akt, p-mTOR, p-p70S6K, and p-4EBP1 ↓ p-STAT3 | ↑ Kahweol-induced apoptosis | [37] | |
| Leukemia | U937 cells | Kahweol | ↑ Caspase 3 ↑ Cytochrome C release ↓ Bcl-2, Bcl-xL, Mcl-1, XIAP ↓ Akt pathways ↑ JNK pathways | ↑ Apoptosis | [38] | |
| NB4, K562, HL60 and KG1 | Cafestol Ara-C | ↑ Caspase 3 ↑ CD11b and CD15 | ↑ Apoptosis ↓ ROS (Reactive Oxygen Species) production by organelles ↓ Clonogenic potential | [39] | ||
| K562 | Kawheol and cafestol | ↑ Granzyme B via ↑ ATF-2, c-Jun, and CREB phosphorylation | ↑ Cytolysis | [40] | ||
| Fibrosarcoma | HT-1080 cells | Kahweol acetate | ↓ PMA-induced MMP-9 via ↓ NF-κB ↓ Akt/JNK1/2/p38 MAPK phosphorylation | ↓ PMA-induced proliferation, invasion, and migration | [30] | |
| Head and Neck Squamous Cell carcinoma | SCC25 CAL27 FaDu | Cafestol Cisplatin | No mention of any pathway | ↑ Apoptosis | [41] | |
| Cancer Type | Animal Model | Cell Line | Kahweol/Derivative | Molecular Target | Functional Impact | Ref. |
|---|---|---|---|---|---|---|
| Prostate Cancer | SCID mice | DU-145 | Kahweol acetate | ↑ Caspase-3 cleavage ↑ PARP cleavage ↓ Bcl-2 ↓ Bcl-xL ↓ AR ↓ CCL2-CCR2 ↓ CCL5-CCR5 | Inhibition of tumour growth | [25] |
| Renal cell carcinoma | BALB/c-nude mice | Caki cells | Cafestol | ↑ Cleavage of PARP ↑ caspase-3 activity ↓ Mcl-1 ↑ PUMA and Bim | ↑ABT-737-mediated apoptosis | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eldesouki, S.; Qadri, R.; Abu Helwa, R.; Barqawi, H.; Bustanji, Y.; Abu-Gharbieh, E.; El-Huneidi, W. Recent Updates on the Functional Impact of Kahweol and Cafestol on Cancer. Molecules 2022, 27, 7332. https://doi.org/10.3390/molecules27217332
Eldesouki S, Qadri R, Abu Helwa R, Barqawi H, Bustanji Y, Abu-Gharbieh E, El-Huneidi W. Recent Updates on the Functional Impact of Kahweol and Cafestol on Cancer. Molecules. 2022; 27(21):7332. https://doi.org/10.3390/molecules27217332
Chicago/Turabian StyleEldesouki, Salma, Rama Qadri, Rashid Abu Helwa, Hiba Barqawi, Yasser Bustanji, Eman Abu-Gharbieh, and Waseem El-Huneidi. 2022. "Recent Updates on the Functional Impact of Kahweol and Cafestol on Cancer" Molecules 27, no. 21: 7332. https://doi.org/10.3390/molecules27217332
APA StyleEldesouki, S., Qadri, R., Abu Helwa, R., Barqawi, H., Bustanji, Y., Abu-Gharbieh, E., & El-Huneidi, W. (2022). Recent Updates on the Functional Impact of Kahweol and Cafestol on Cancer. Molecules, 27(21), 7332. https://doi.org/10.3390/molecules27217332