
Inhibition of Aflatoxin B1 Synthesis in Aspergillus flavus by Mate (Ilex paraguariensis), Rosemary (Rosmarinus officinalis) and Green Tea (Camellia sinensis) Extracts: Relation with Extract Antioxidant Capacity and Fungal Oxidative Stress Response Modulation


 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
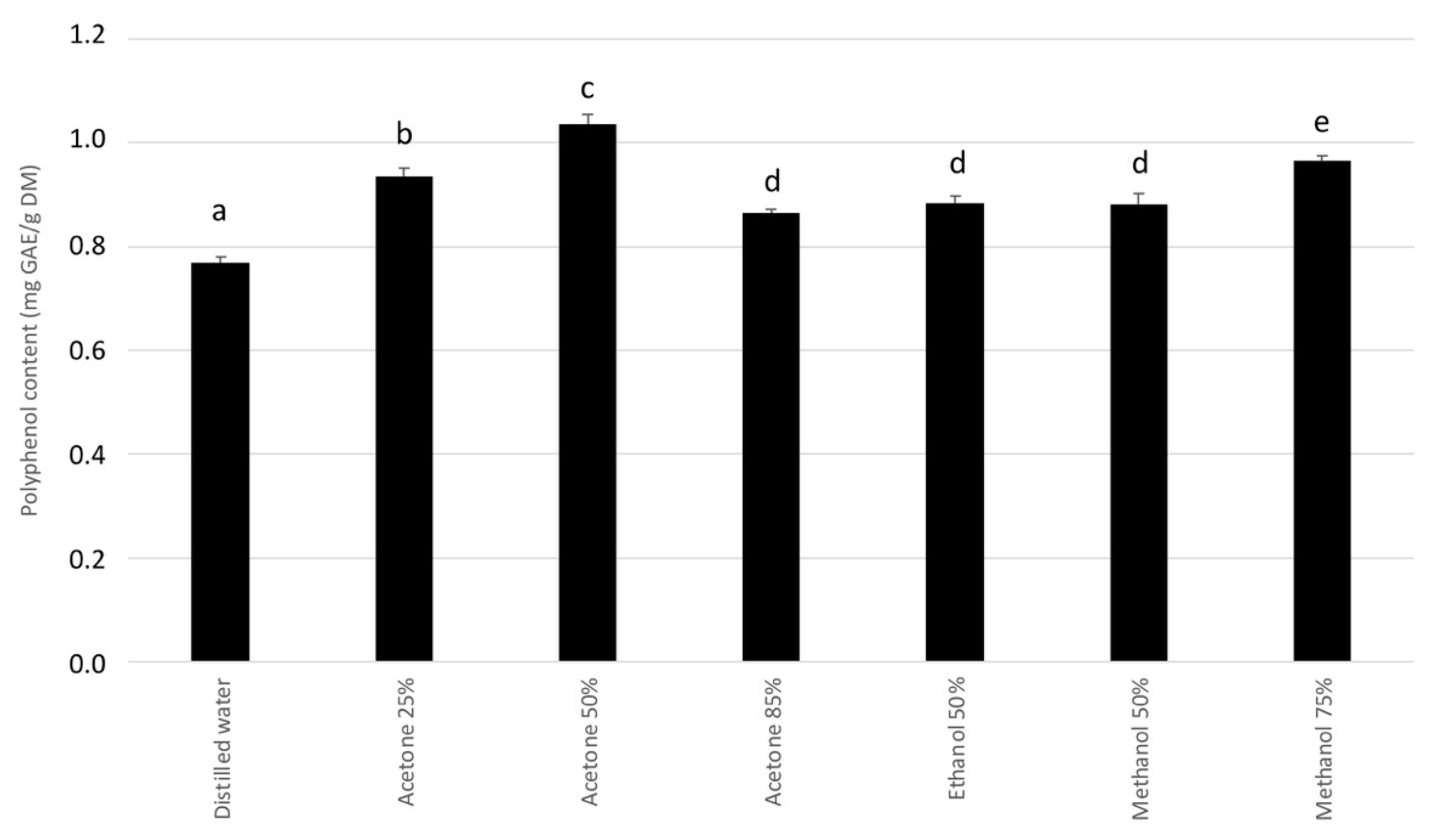
2.1. Choice of Extraction Solvent
2.2. Phenolic Content and Antioxidative Activity of Extracts
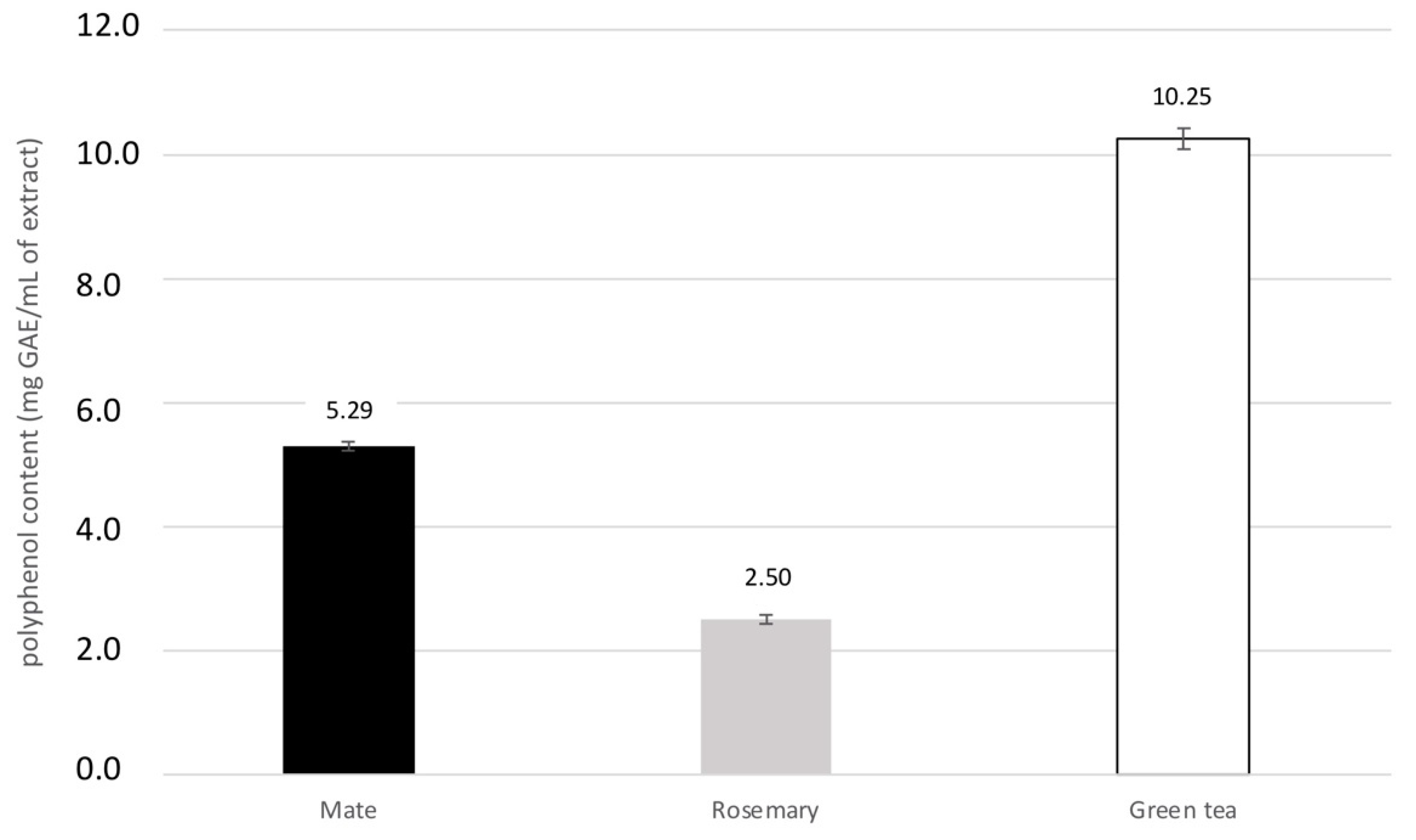
2.2.1. Total Phenolic Content
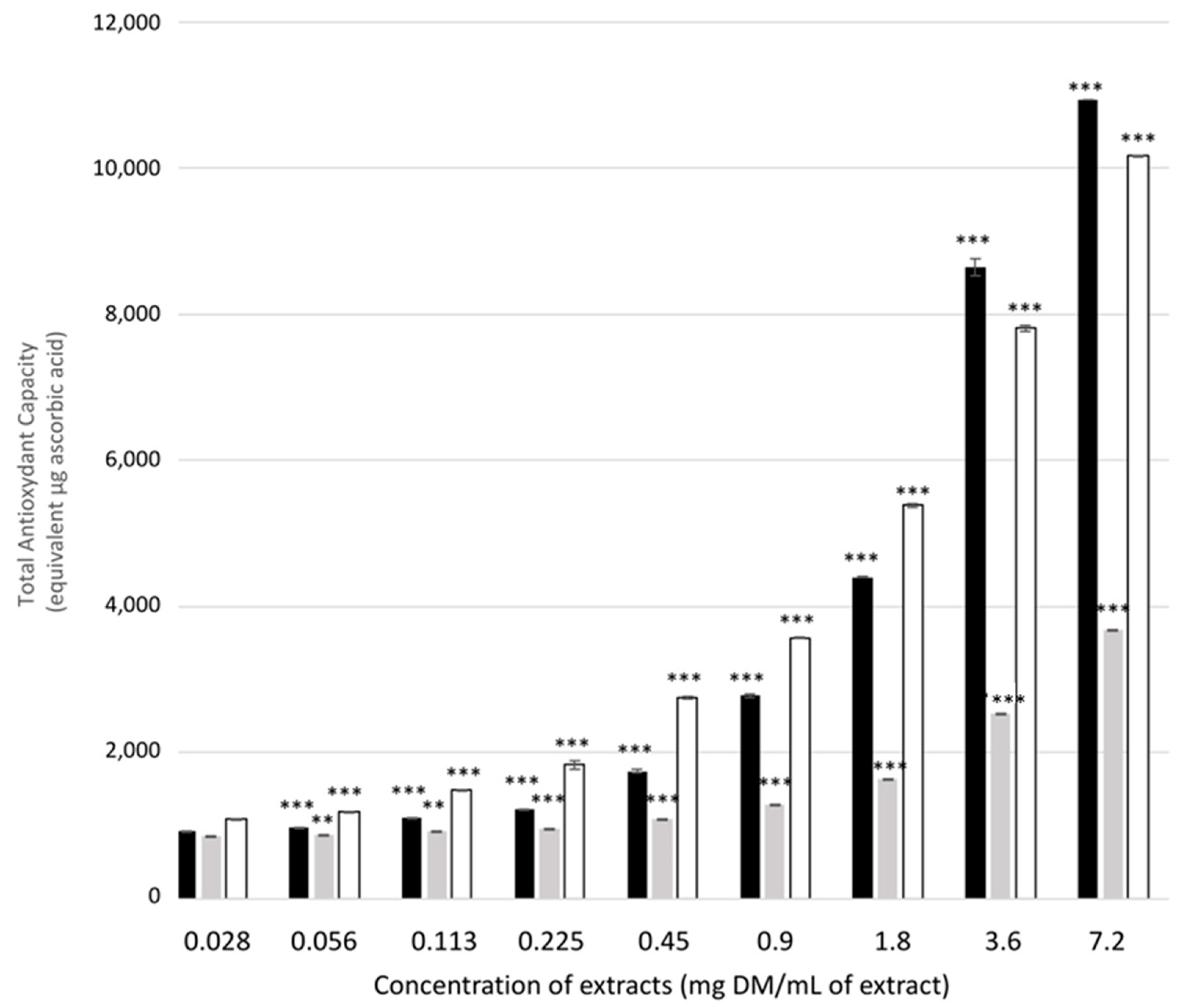
2.2.2. Total Antioxidant Activity of Plant Extracts
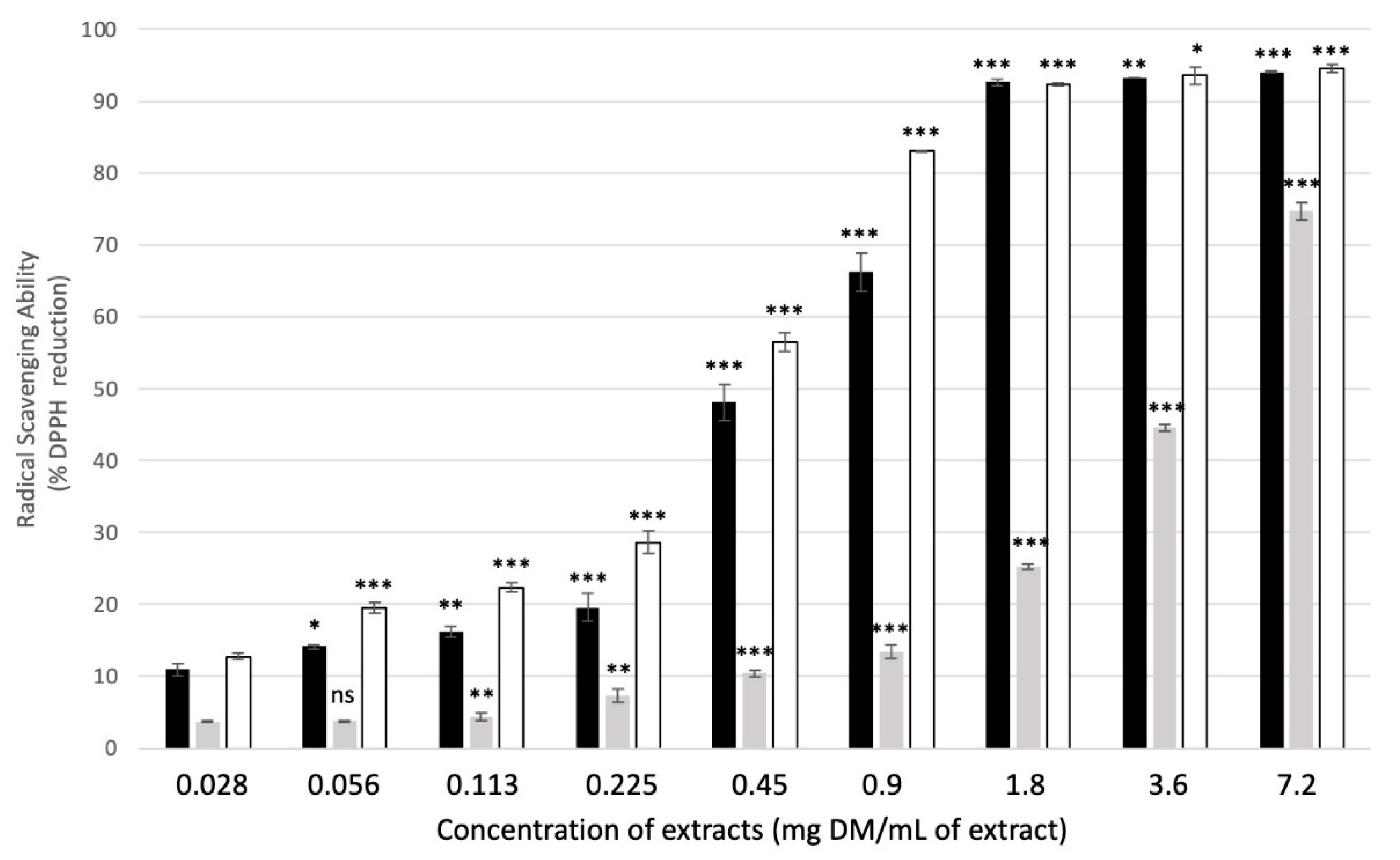
2.2.3. Antiradical Activity of Plant Extracts
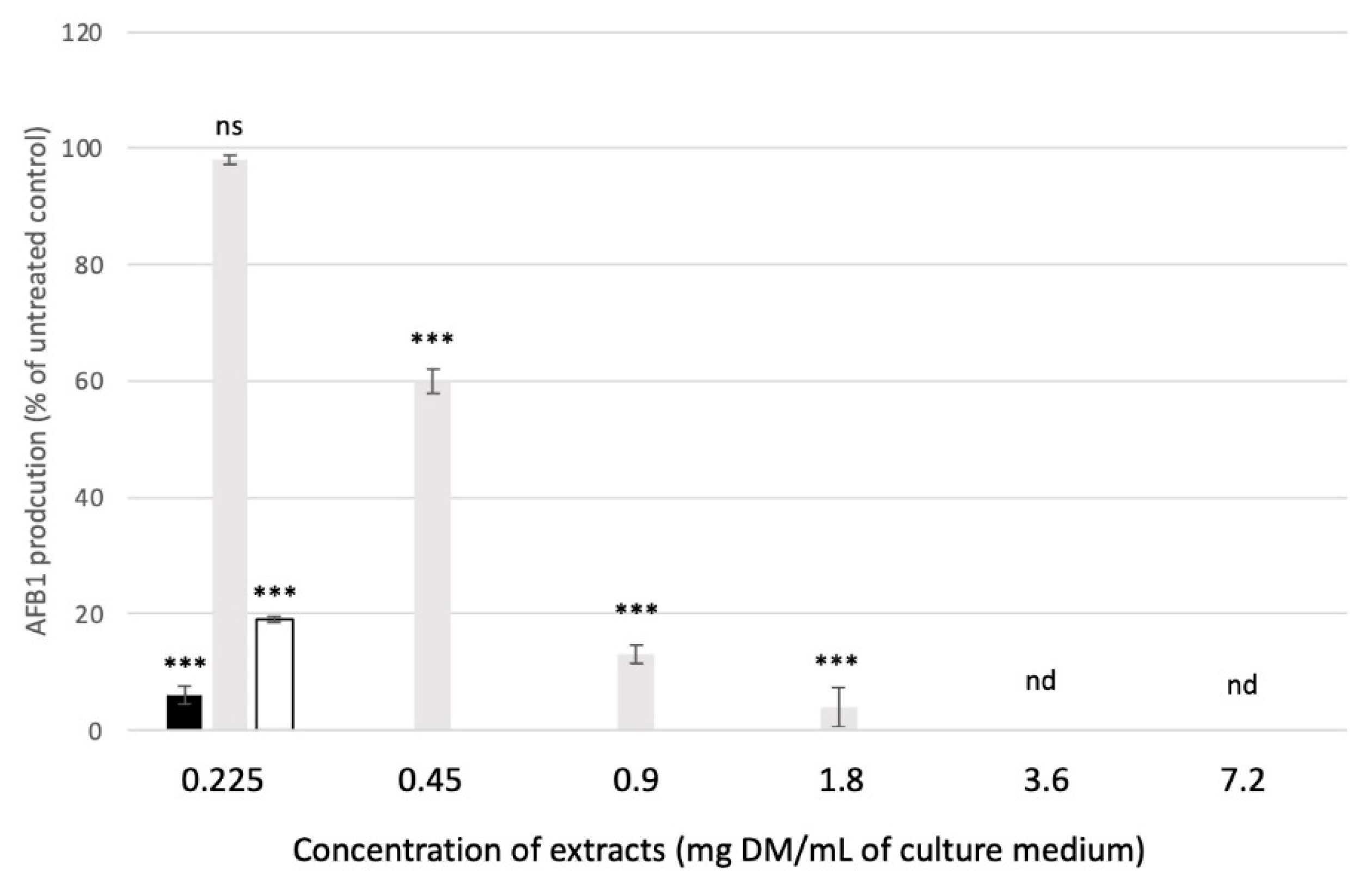
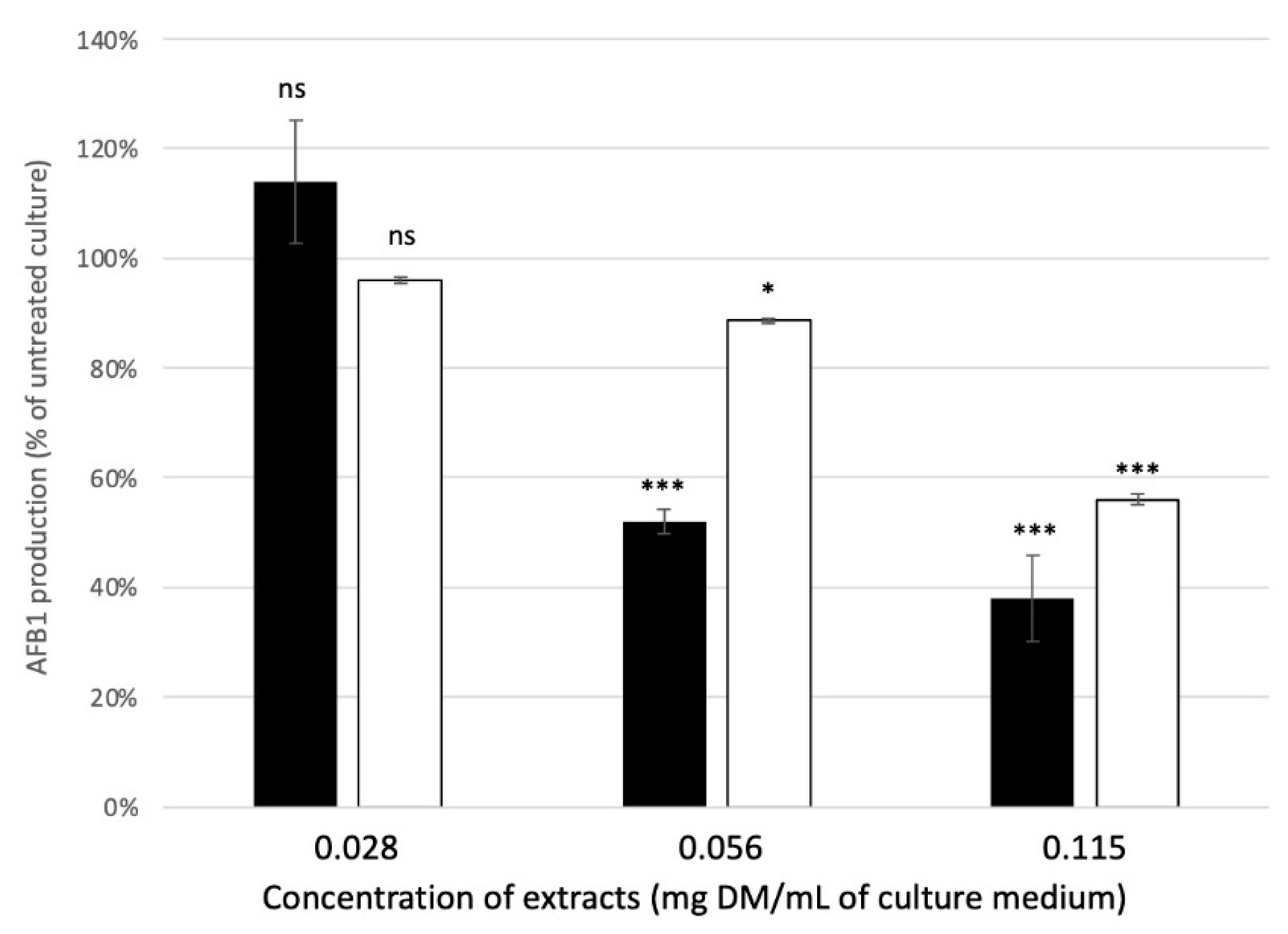
2.3. Impact of Plant Extracts on the Growth of Aspergillus flavus and on the Production of AFB1
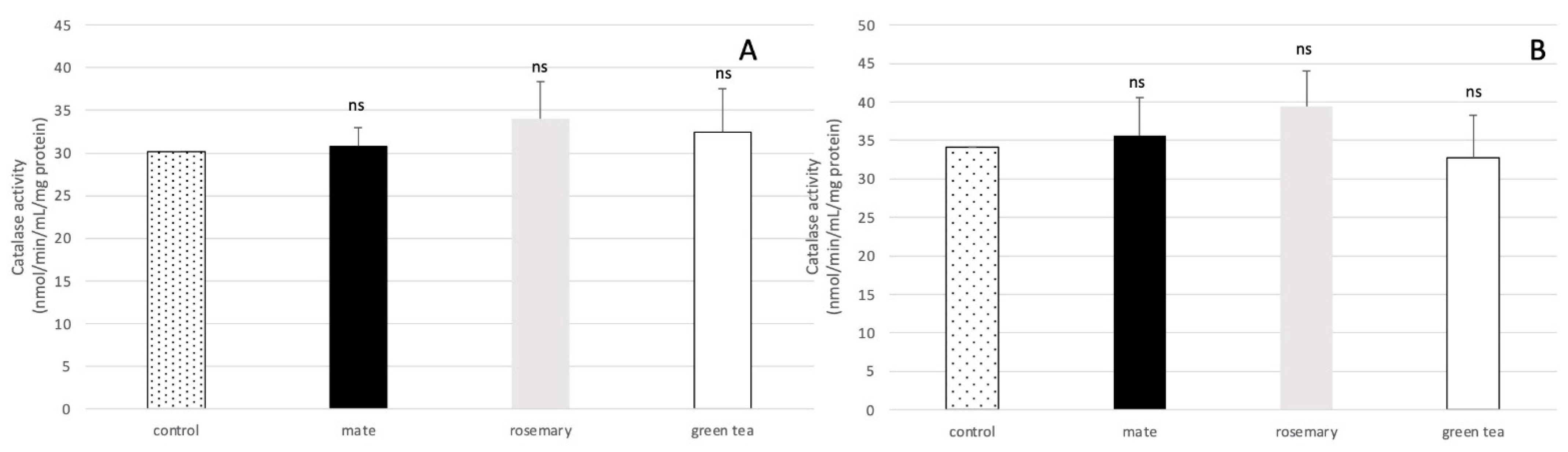
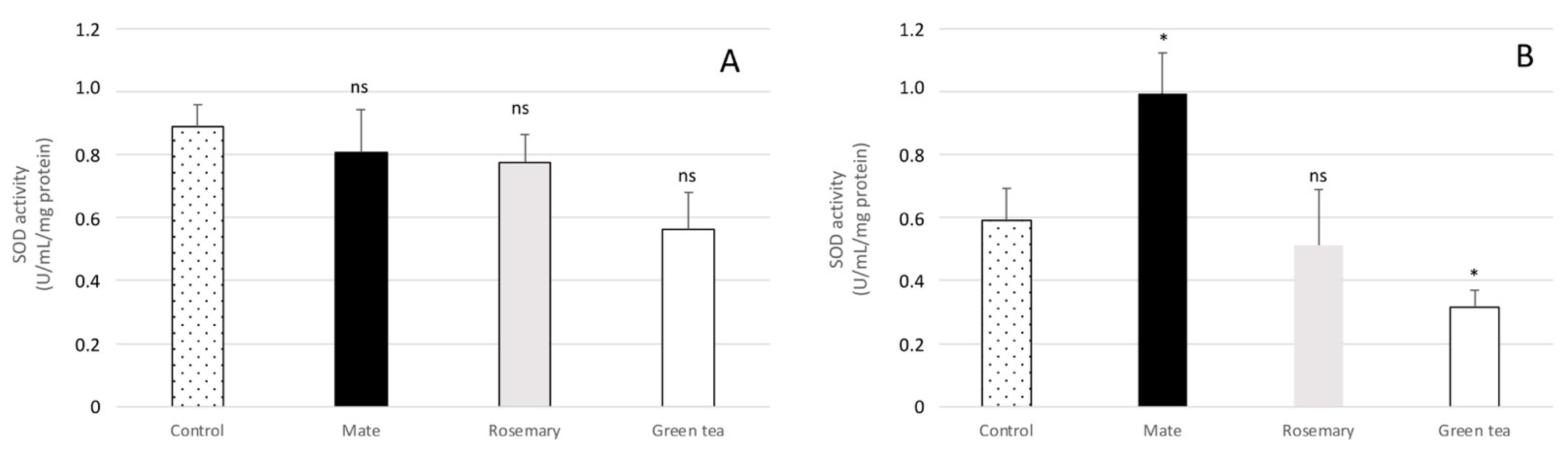
2.4. Effect of Plant Extracts on Oxidative Stress Enzymatic Response in Aspergillus flavus
2.5. Correlation between Anti-AFB1 Potential of the Three Extracts and Phenolic Content, Antioxidant and Antiradical Activities
2.6. Impact of Rutin on AFB1 Synthesis in Aspergillus flavus
3. Materials and Methods
3.1. Chemicals
3.2. Preparation of Plant Extracts
3.3. Determination of Total Phenolic Content
3.4. Determination of Total Antioxidant Activity
3.5. Radical Scavenging Ability of Plant Extracts
3.6. Impact of Extracts on Fungal Growth and AFB1 Synthesis
3.7. AFB1 Quantification
3.8. Analysis of Fungal Enzymatic Activities
3.8.1. Sample Preparation
3.8.2. Measurement of Superoxide Dismutase Activity
3.8.3. Measurement of Catalase Activity
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Nov, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklen, F.; Mahakarnchanakul, W.; Samson, R.A.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2019, 93, 37–59. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.H.; Phillips, T.D.; Jolly, P.E.; Stiles, J.K.; Jolly, C.M.; Aggarwal, D. Human aflatoxicosis in developing countries: A review of toxicology, exposure, potential health consequences, and interventions. Am. J. Clin. Nutr. 2004, 80, 1106–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnonlonfin, G.J.B.; Hell, K.; Adjovi, Y.; Fandohan, P.; Koudande, D.O.; Mensah, G.A.; Sanni, A.; Brimer, L. A review on aflatoxin contamination and its implication in the developing world: A sub-Saharan African perspective. Crit. Rev. Food Sci. Nutr. 2013, 53, 349–365. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Rodriguez, A.; Magan, N. Effect of climate change on Aspergillus flavus and aflatoxin B 1 production. Front. Microbiol. 2014, 5, 348. [Google Scholar] [CrossRef] [Green Version]
- Battilani, P.; Toscano, P.; Van der Fels-Klerx, J.; Moretti, A.; Camardo Leggieri, M.; Brera, C.; Rortais, T.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef] [Green Version]
- Bailly, S.; Carvajal, A.; Lorber, S.; Puel, O.; Oswald, I.P.; Bailly, J.D.; Orlando, B. Occurrence and identification of Aspergillus section Flavi in the context of the emergence of aflatoxins in French maize. Toxins 2018, 10, 525. [Google Scholar] [CrossRef] [Green Version]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Ann. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef]
- Villers, P. Aflatoxin and safe storage. Front. Microbiol. 2014, 5, 158. [Google Scholar] [CrossRef] [Green Version]
- Codex Alimentarius. Code of practice for the prevention and reduction of aflatoxin contamination in peanuts CAC/RCP 55-2004. In General Principles of Food Hygiene; FAO: Rome, Italy, 2004. [Google Scholar]
- Lagogianni, S.; Tsitsigiannis, I. Effective chemical management for prevention of aflatoxins in maize. Phytopathol. Mediterr. 2018, 57, 186–197. [Google Scholar] [CrossRef]
- Parikhani, M.P.; Borkhani, F.R.; Fami, H.S.; Motiee, N.; Hosseinpoor, A. Major barriers to application of Good Agricultural Practices (GAPs) technologies in sustainability of livestock units. Int. J. Agric. Manag. Dev. 2015, 5, 169–178. [Google Scholar] [CrossRef]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdisc. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, I.; Imadi, S.R.; Shazadi, K.; Gul, A.; Hakeem, K.R. Effects of pesticides on environment. In Plant, Soil and Microbes; Hakeem, K.R., Ed.; Springer: Cham, Switzerland, 2015; pp. 253–269. [Google Scholar] [CrossRef]
- Nicolopoulou-Stamati, P.; Maipas, S.; Kotampasi, C.; Stamatis, P.; Hens, L. Chemical pesticides and human health: The urgent need for a new concept in agriculture. Front. Public Health 2016, 4, 148. [Google Scholar] [CrossRef] [Green Version]
- Moreira da Silva, J.F.; Peluzio, J.M.; Prado, G.; Cruz Madeira, J.E.G.; Oliveira Silva, M.; de Morais, P.B.; Rosa, C.A.; Pimenta, R.S.; Nicoli, J.R. Use of probiotics to control aflatoxin production in peanut grains. Sci. World J. 2015, 2015, 959138. [Google Scholar] [CrossRef] [Green Version]
- Mauro, A.; Garcia-Cela, E.; Pietri, A.; Cotty, P.J.; Battilani, P. Biological control products for aflatoxin prevention in Italy: Commercial field evaluation of atoxigenic Aspergillus flavus active ingredients. Toxins 2018, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Nazareth, T.D.M.; Luz, C.; Torrijos, R.; Quiles, J.M.; Bittencourt, L.F.; Mañes, J.; Meca, G. Potential application of lactic acid bacteria to reduce aflatoxin B1 and fumonisin B1 occurrence on corn kernels and corn ears. Toxins 2020, 12, 21. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Zhang, Q.; Zhang, W.; Mao, J.; Li, P. Control of aflatoxigenic molds by antagonistic microorganisms: Inhibitory behaviors, bioactive compounds, related mechanism and influencing factors. Toxins 2020, 12, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojiambo, P.S.; Battilani, P.; Cary, J.W.; Blum, B.H.; Carbone, I. Cultural and genetic approaches to manage aflatoxin contamination: Recent insights provide opportunities for improved control. Phytopathology 2018, 108, 1024–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loi, M.; Paciolla, C.; Logrieco, A.F.; Mulè, G. Plant bioactive compounds in pre- and postharvest management for aflatoxins reduction. Front. Microbiol. 2020, 11, 243. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, R.; Caceres, I.; Puel, O.; Bailly, S.; Atoui, A.; Oswald, I.P.; El Khoury, A.; Bailly, J.D. Identification of the anti-aflatoxinogenic activity of Micromeria graeca and elucidation of its molecular mechanism in Aspergillus flavus. Toxins 2017, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, C.; Cadenillas, L.; El Mahgubi, A.; Caceres, I.; Durrieu, V.; Mathieu, C.; Bailly, J.D. Mimosa tenuiflora aqueous extract: Role of condensed tannins in anti-aflatoxin B1 activity in Aspergillus flavus. Toxins 2021, 13, 391. [Google Scholar] [CrossRef]
- Caceres, I.; El Khoury, R.; Medina, A.; Lippi, Y.; Naylies, C.; Atoui, A.; El Khoury, A.; Oswald, I.P.; Bailly, J.D.; Puel, O. Deciphering the anti-aflatoxinogenic properties of eugenol using a large-scale q-PCR approach. Toxins 2016, 8, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caceres, I.; El Khoury, R.; Bailly, S.; Oswald, I.P.; Puel, O.; Bailly, J.D. Piperine inhibits aflatoxin B1 production in Aspergillus flavus by modulating fungal oxidative stress response. Fungal Genet. Biol. 2017, 107, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Caceres, I.; Al Khoury, A.; El Khoury, R.; Lorber, S.; Oswald, I.P.; El Khoury, A.; Atoui, A.; Puel, O.; Bailly, J.D. Aflatoxin biosynthesis and genetic regulation: A review. Toxins 2020, 12, 150. [Google Scholar] [CrossRef] [Green Version]
- Kujawska, M. Yerba Mate (Ilex paraguariensis) beverage: Nutraceutical ingredient or conveyor for the intake of medicinal plants. Evidence from Paraguayan Folk Medicine. Evid. Based Complement Alternat. Med. 2018, 2018, 6849317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira da Silva, T.; Dillenburg Meinhart, A.; Cristina, T.; Lima de Souza, T.C.; Teixeira Filho, J.; Teixeira Godoy, H. Phenolic compounds from yerba mate-based beverages—A multivariate optimization. Food Chem. 2016, 190, 1159–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Minero, F.J.; Bravo-Diaz, L.; Ayala-Gomez, A. Rosmarinus officinalis L. (Rosemary): An ancient plant with uses in personal healthcare and cosmetics. Cosmetics 2020, 7, 77. [Google Scholar] [CrossRef]
- Meegahakumbura, M.K.; Wambulwa, M.C.; Li, M.M.; Thapa, K.K.; Sun, Y.S.; Möller, M.; Xu, J.C.; Yang, J.B.; Liu, J.; Liu, B.Y.; et al. Domestication origin and breeding history of the tea plant (Camellia sinensis) in China and India based on nuclear microsatellites and cpDNA sequence data. Front. Plant Sci. 2018, 8, 2270. [Google Scholar] [CrossRef] [Green Version]
- Yi, M.; Wu, X.; Zhuang, W.; Xia, L.; Chen, Y.; Zhao, R.; Wan, Q.; Du, L.; Zhou, Y. Tea consumption and health outcomes: Umbrella review of meta-analysis of observational studies in humans. Mol. Nutr. Food Res. 2019, 63, 1900389. [Google Scholar] [CrossRef]
- Musial, C.; Kuban-jankowska, A.; Gorska-Ponikowska, M. Beneficial properties of green tea catechins. Int. J. Mol. Sci. 2020, 21, 1744. [Google Scholar] [CrossRef] [Green Version]
- Van Ngo, T.; Scarlett, C.J.; Bowyer, M.C.; Ngo, P.D.; Van Vuong, Q. Impact of different extraction solvents on bioactive compounds and antioxidant capacity from the root of Salacia chinensis L. J. Food Quality 2017, 2017, 9305047. [Google Scholar] [CrossRef] [Green Version]
- Bae, I.K.; Ham, H.M.; Jeong, M.H.; Kim, D.H.; Kim, H.J. Simultaneous determination of 15 phenolic compounds and caffeine in teas and mate using RP-HPLC/UV detection: Method development and optimization of extraction process. Food Chem. 2015, 172, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Markovicz Bastos, D.H.; Saldanha, L.A.; Catharino, R.R.; Sawaya, A.C.H.F.; Cunha, I.B.S.; Carvalho, P.O.; Eberlin, M.N. Phenolic antioxidants identified by ESI-MS from Yerba mate (Ilex paraguariensis) and green tea (Camelia sinensis) extracts. Molecules 2007, 12, 423–432. [Google Scholar] [CrossRef]
- Heck, C.I.; Schmalko, M.; Gonzalez de Mejia, E. Effect of growing and drying conditions on the phenolic composition of mate teas (Ilex paraguariensis). J. Agric. Food Chem. 2008, 56, 8394–8403. [Google Scholar] [CrossRef] [PubMed]
- Kabran, G.R.M.; Ambeu N’ta, C.; Mamyrbekova-Bekro, J.A.; Bekro, Y.A. Phenols et flavonoïdes totaux dans les extraits organiques de dix plantes utilisees dans la traditherapie du cancer du sein en Côte d’Ivoire. Eur. J. Sci. Res. 2012, 68, 182–190. [Google Scholar]
- Popovici, C.; Saykova, I.; Tylkowski, B. Evaluation de l’activité antioxydant des composés phénoliques par la réactivité avec le radical libre DDPH. Rev. Gen. Indust. 2009, 4, 25–39. [Google Scholar]
- Dasgupta, N.; De, B. Antioxidant activity of Piper betle L. leaf extract in vitro. Food Chem. 2004, 88, 219–224. [Google Scholar] [CrossRef]
- Li, H.Y.; Hao, Z.B.; Wang, X.L.; Huang, L.; Li, J.P. Antioxidant activities of extracts and fractions from Lysimachia foenum-graecum Hance. Biores. Technol. 2009, 100, 970–974. [Google Scholar] [CrossRef]
- Afonso, V.; Champy, R.; Mitrovic, D.; Collin, P.; Lomri, A. Reactive oxygen species and superoxide dismutases: Role in joint diseases. Joint Bone Spine 2007, 74, 324–329. [Google Scholar] [CrossRef]
- Unachukwu, U.J.; Ahmed, S.; Kavalier, A.; Lyles, J.T.; Kennelly, E.J. White and green teas (Camellia sinensis var. sinensis): Variation in phenolic, methylxanthine and antioxidant profiles. J. Food Sci. 2010, 75, 541–548. [Google Scholar] [CrossRef] [Green Version]
- Dudonne, S.; Vitrac, X.; Coutiere, P.; Woillez, M.; Merillon, J.M. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD and ORAC assyas. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Ben Miri, Y.; Djenane, D. Evaluation of protective impact of Algerian Cuminum cyminum L. and Coriandrum sativum L. essential oils on Aspergillus flavus growth and Aflatoxin B1 production. Pakistan J. Biol. Sci. 2018, 21, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Gorran, A.; Farzaneh, M.; Shivazad, M.; Rezaeian, M.; Ghassempour, A. Aflatoxin B1 reduction of Aspergillus flavus by three medicinal plants (Lamiaceae). Food Control 2013, 31, 218–223. [Google Scholar] [CrossRef]
- Shukla, R.; Singh, P.; Prakash, B.; Dubey, N.K. Antifungal, aflatoxin inhibition and antioxidant activity of Callistemon lanceolatus (Sm.) sweet essential oil and its major component 1,8-cineole against fungal isolates from chickpea seeds. Food Control 2012, 25, 27–33. [Google Scholar] [CrossRef]
- Al Khoury, A.; Sleiman, R.; Atoui, A.; Hindhieh, P.; Maroun, R.; Bailly, J.D.; El Khoury, A. Antifungal and anti-aflatoxigenic properties of organs of Cannabis sativa L.: Relation to phenolic content and antioxidant capacities. Arch. Microbiol. 2021, 203, 4485–4492. [Google Scholar] [CrossRef]
- Reverberi, M.; Fabbri, A.A.; Zjalic, S.; Ricelli, A.; Punelli, F.; Fanelli, C. Antioxidant enzymes stimulation in Aspergillus parasiticus by Lentinula edodes inhibits aflatoxin production. Appl. Microbiol. Biotechnol. 2005, 69, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Al Sereiti, M.R.; Abu-Amer, K.M.; Sen, P. Pharmacology of rosemary (Rosmarinus officinalis) and its therapeutic potentials. Indian J. Exp. Biol. 1999, 37, 124–130. [Google Scholar]
- Bressan Waller, S.; Martins Madrid, I.; Hoffmann, J.F.; Picoli, T.; Brum Cleff, M.; Clasen Chaves, F.; de Faria, R.O.; Araujo Meireles, M.C.; Braga de Mello, J.R. Chemical composition and cytotoxicity of extracts of marjoram and rosemary and their activity against Sporothrix brasiliensis. J. Med. Microbiol. 2017, 66, 1076–1083. [Google Scholar] [CrossRef]
- Kochman, J.; Jakubbczyk, K.; Antoniewicz, J.; Mruk, H.; Janda, K. Health benefits and chemical composition of Matcha green tea: A review. Molecules 2020, 26, 85. [Google Scholar] [CrossRef] [PubMed]
- Paula Lima, J.; Farah, A.; King, B.; de Paulis, T.; Martin, P.R. Distribution of major chlorogenic acids and related compounds in Brazilian greenand toasted Ilex paraguensis (mate) leaves. J. Agric. Food Chem. 2016, 64, 2361–2370. [Google Scholar] [CrossRef]
- Paster, N.; Juven, B.J.; Harshemesh, H. Antimicrobial activity and inhibition of aflatoxin B1 formation by olive plant tissue constituents. J. Appl. Microbiol. 1988, 64, 293–297. [Google Scholar]
- Campello Telles, A.; Kupski, L.; Badiale Furlong, E. Phenolic compound in beans as protection against mycotoxins. Food Chem. 2017, 214, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Z.; Wang, Y.; Wang, J.; Yang, P. Quercetin Inhibits the Proliferation and Aflatoxins Biosynthesis of Aspergillus flavus. Toxins 2019, 11, 154. [Google Scholar] [CrossRef] [PubMed]
- Muradic, S.; Salihovic, M.; Klepo, L.; Sofic, E.; Kalher, K. Quantification of some phenolic acid and rutin in the leaves of rosemary from Turkey and Croatia. Planta. Med. 2010, 76, 120. [Google Scholar] [CrossRef]
- Yang, J.; Guo, J.; Yuan, J. In vitro antioxidant properties of rutin. LWT-Food Sci. Technol. 2008, 4, 1060–1066. [Google Scholar] [CrossRef]
- Bianchin, M.; Pereira, D.; de Florio Almeida, J.; de Moura, C.; Siminionatto Pinheiro, R.; Serafini Heldt, L.F.; Isidoro Haminiuk, C.W.; Teresinha Carpes, S. Antioxidant properties of lyophilized rosemary and sage extracts and its effect to prevent lipid oxidation in poultry pate. Molecules 2020, 25, 5160. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Abi-Khattar, A.M.; Rajha, H.N.; Abdel-Massih, R.M. Intensification of polyphenol extraction from olive. Antioxidants 2019, 8, 227. [Google Scholar] [CrossRef] [Green Version]
- El Mahgubi, A.; Puel, O.; Bailly, S.; Tadrist, S.; Querin, A.; Ouadia, A.; Oswald, I.P.; Bailly, J.D. Distribution and toxigenicity of Aspergillus section Flavi in spices marketed in Morocco. Food Control 2013, 32, 143–148. [Google Scholar] [CrossRef]
- Miklos, G.; Angeli, C.; Ambrus, A.; Nagy, A.; Kardos, V.; Zentai, A.; Kerekes, K.; Farkas, Z.; Jozwiak, A.; Bartok, T. Detection of aflatoxins in different matrices and food-chain positions. Front. Microbiol. 2020, 11, 1916. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Growth | |||
|---|---|---|---|
| (% of Untreated Control) | |||
| Mate | Rosemary | Green Tea | |
| Concentration of extract | |||
| (mg DM/mL a) | |||
| 0 | 100% | 100% | 100% |
| 0.0281 | 96% *** | nt b | 96% ** |
| 0.0563 | 97% * | nt | 97% ** |
| 0.113 | 96% ** | nt | 96% ** |
| 0.225 | 98% *** | 98% ** | 88% *** |
| 0.45 | 96% ** | 96% ** | 91% ** |
| 0.9 | 99% ** | 97% * | 95% * |
| 1.8 | 95% * | 94% *** | 92% ** |
| 3.6 | 100% ** | 92% ** | 90% * |
| 7.2 | 98% ** | 84% ** | 82% * |
| Anti-Aflatoxigenic Activity | |||
|---|---|---|---|
| Mate | Rosemary | Green Tea | |
| Polyphenol content | 0.825 | 0.753 | 0.935 |
| Antiradical activity | 0.711 | 0.660 | 0.853 |
| Antioxidant activity | 0.5 | 0.668 | 0.660 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Khoury, A.; El Khoury, A.; Rocher, O.; Hindieh, P.; Puel, O.; Maroun, R.G.; Atoui, A.; Bailly, J.-D. Inhibition of Aflatoxin B1 Synthesis in Aspergillus flavus by Mate (Ilex paraguariensis), Rosemary (Rosmarinus officinalis) and Green Tea (Camellia sinensis) Extracts: Relation with Extract Antioxidant Capacity and Fungal Oxidative Stress Response Modulation. Molecules 2022, 27, 8550. https://doi.org/10.3390/molecules27238550
Al Khoury A, El Khoury A, Rocher O, Hindieh P, Puel O, Maroun RG, Atoui A, Bailly J-D. Inhibition of Aflatoxin B1 Synthesis in Aspergillus flavus by Mate (Ilex paraguariensis), Rosemary (Rosmarinus officinalis) and Green Tea (Camellia sinensis) Extracts: Relation with Extract Antioxidant Capacity and Fungal Oxidative Stress Response Modulation. Molecules. 2022; 27(23):8550. https://doi.org/10.3390/molecules27238550
Chicago/Turabian StyleAl Khoury, Anthony, André El Khoury, Ophélie Rocher, Pamela Hindieh, Olivier Puel, Richard G. Maroun, Ali Atoui, and Jean-Denis Bailly. 2022. "Inhibition of Aflatoxin B1 Synthesis in Aspergillus flavus by Mate (Ilex paraguariensis), Rosemary (Rosmarinus officinalis) and Green Tea (Camellia sinensis) Extracts: Relation with Extract Antioxidant Capacity and Fungal Oxidative Stress Response Modulation" Molecules 27, no. 23: 8550. https://doi.org/10.3390/molecules27238550
APA StyleAl Khoury, A., El Khoury, A., Rocher, O., Hindieh, P., Puel, O., Maroun, R. G., Atoui, A., & Bailly, J. -D. (2022). Inhibition of Aflatoxin B1 Synthesis in Aspergillus flavus by Mate (Ilex paraguariensis), Rosemary (Rosmarinus officinalis) and Green Tea (Camellia sinensis) Extracts: Relation with Extract Antioxidant Capacity and Fungal Oxidative Stress Response Modulation. Molecules, 27(23), 8550. https://doi.org/10.3390/molecules27238550