
Anxiolytic and Antioxidant Effect of Phytoecdysteroids and Polyphenols from Chenopodium quinoa on an In Vivo Restraint Stress Model

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Acute Oral Toxicity of FFIs from Chenopodium quinoa
2.2. 36-Day Study of the Quinoa FFI Adaptogenic Properties in a Model of Immobilization-Induced Emotional Stress
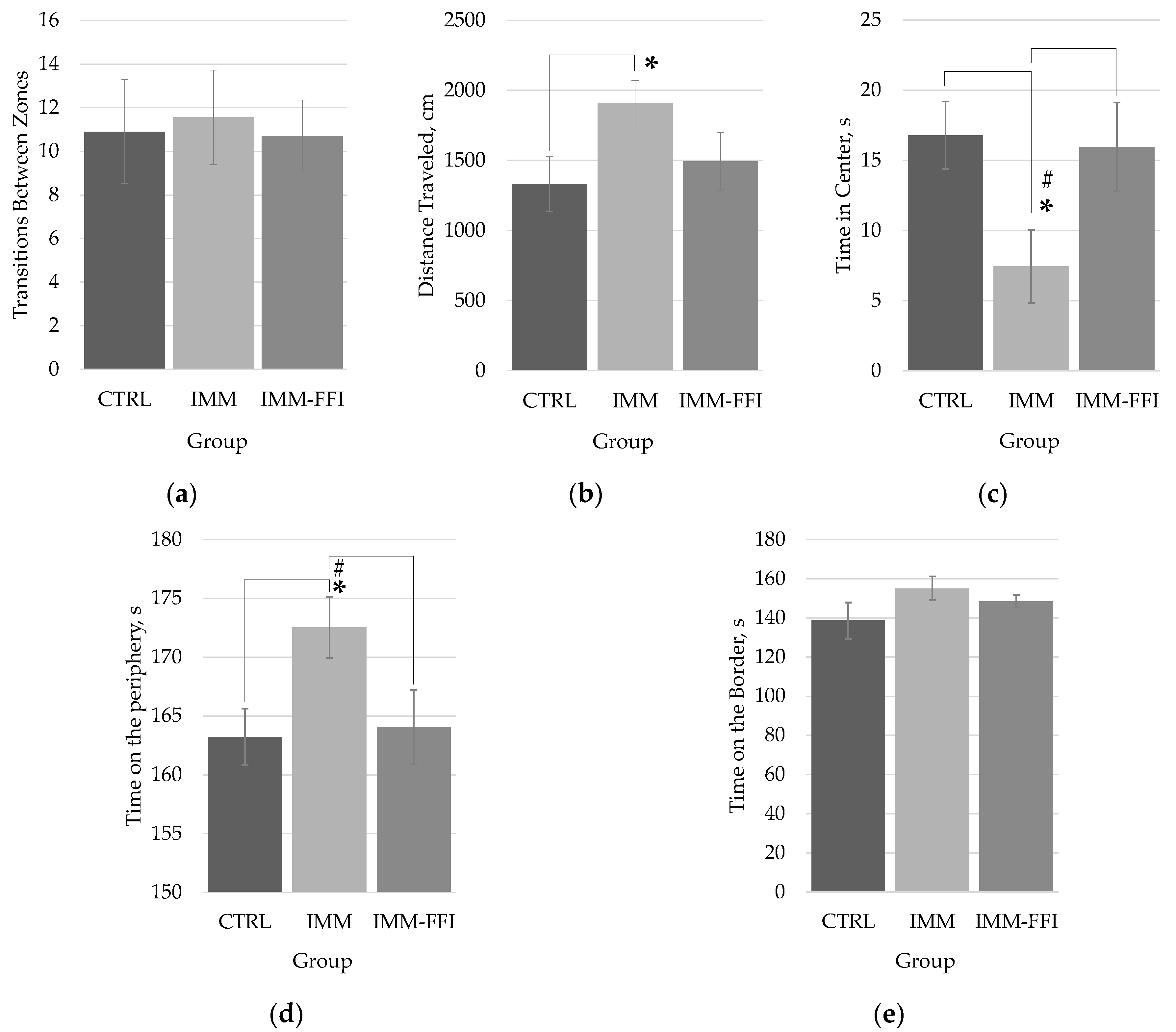
2.2.1. Integral Indicators
2.2.2. Memory Function and Behavioral Responses
2.2.3. Biochemical Indices
3. Discussion
4. Materials and Methods
4.1. Preparation and Characterization of the FFIs from Chenopodium quinoa Seeds
4.2. Animals and Experimental Design
4.2.1. Animals and Ethics
4.2.2. Acute Oral Toxicity: Fixed Dose Procedure
4.2.3. 36-Day Study of the Quinoa FFI Adaptogenic Properties in a Model of Immobilization-Induced Emotional Stress
4.3. Methods for Assessing Biochemical Parameters
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Head, K.A.; Kelly, G.S. Nutrients and Botanicals for Treatment of Stress: Adrenal Fatigue, Neurotransmitter Imbalance, Anxiety, and Restless Sleep. Altern. Med. Rev. 2009, 14, 114–140. [Google Scholar]
- Panossian, A.G.; Efferth, T.; Shikov, A.N.; Pozharitskaya, O.N.; Kuchta, K.; Mukherjee, P.K.; Banerjee, S.; Heinrich, M.; Wu, W.; Guo, D.; et al. Evolution of the Adaptogenic Concept from Traditional Use to Medical Systems: Pharmacology of Stress- and Aging-related Diseases. Med. Res. Rev. 2021, 41, 630–703. [Google Scholar] [CrossRef]
- Sidorova, Y.S.; Petrov, N.A.; Shipelin, V.A.; Mazo, V.K. Spinach and Quinoa—Prospective Food Sources of Biologically Active Substances. Vopr. Pitan. [Probl. Nutr.] 2020, 89, 100–106. [Google Scholar] [CrossRef]
- Graf, B.L.; Poulev, A.; Kuhn, P.; Grace, M.H.; Lila, M.A.; Raskin, I. Quinoa Seeds Leach Phytoecdysteroids and Other Compounds with Anti-Diabetic Properties. Food Chem. 2014, 163, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, J.; Kong, Z.; Huan, X.; Li, L.; Zhang, P.; Wang, Q.; Guo, Y.; Zhu, W.; Qin, P. Transcriptomics and Metabolomics Analyses of the Mechanism of Flavonoid Synthesis in Seeds of Differently Colored Quinoa Strains. Genomics 2022, 114, 138–148. [Google Scholar] [CrossRef]
- Antognoni, F.; Potente, G.; Biondi, S.; Mandrioli, R.; Marincich, L.; Ruiz, K.B. Free and Conjugated Phenolic Profiles and Antioxidant Activity in Quinoa Seeds and Their Relationship with Genotype and Environment. Plants 2021, 10, 1046. [Google Scholar] [CrossRef]
- Melini, V.; Melini, F. Modelling and Optimization of Ultrasound-Assisted Extraction of Phenolic Compounds from Black Quinoa by Response Surface Methodology. Molecules 2021, 26, 3616. [Google Scholar] [CrossRef]
- Lin, M.; Han, P.; Li, Y.; Wang, W.; Lai, D.; Zhou, L. Quinoa secondary metabolites and their biological activities or functions. Molecules 2019, 24, 2512. [Google Scholar] [CrossRef] [Green Version]
- Filho, A.M.M.; Pirozi, M.R.; Borges, J.T.D.S.; Pinheiro Sant’Ana, H.M.; Chaves, J.B.P.; Coimbra, J.S.D.R. Quinoa: Nutritional, Functional, and Antinutritional Aspects. Crit. Rev. Food Sci. Nutr. 2017, 57, 1618–1630. [Google Scholar] [CrossRef]
- Ong, E.S.; Pek, C.J.N.; Tan, J.C.W.; Leo, C.H. Antioxidant and Cytoprotective Effect of Quinoa (Chenopodium quinoa Willd.) with Pressurized Hot Water Extraction (PHWE). Antioxidants 2020, 9, 1110. [Google Scholar] [CrossRef]
- Han, Y.; Chi, J.; Zhang, M.; Zhang, R.; Fan, S.; Huang, F.; Xue, K.; Liu, L. Characterization of Saponins and Phenolic Compounds: Antioxidant Activity and Inhibitory Effects on α-Glucosidase in Different Varieties of Colored Quinoa (Chenopodium quinoa Willd.). Biosci. Biotechnol. Biochem. 2019, 83, 2128–2139. [Google Scholar] [CrossRef]
- Song, C.; Lv, W.; Li, Y.; Nie, P.; Lu, J.; Geng, Y.; Heng, Z.; Song, L. Alleviating the Effect of Quinoa and the Underlying Mechanism on Hepatic Steatosis in High-Fat Diet-Fed Rats. Nutr. Metab. 2021, 18, 106. [Google Scholar] [CrossRef]
- Foucault, A.-S.; Even, P.; Lafont, R.; Dioh, W.; Veillet, S.; Tomé, D.; Huneau, J.-F.; Hermier, D.; Quignard-Boulangé, A. Quinoa Extract Enriched in 20-Hydroxyecdysone Affects Energy Homeostasis and Intestinal Fat Absorption in Mice Fed a High-Fat Diet. Physiol. Behav. 2014, 128, 226–231. [Google Scholar] [CrossRef]
- Noratto, G.D.; Murphy, K.; Chew, B.P. Quinoa Intake Reduces Plasma and Liver Cholesterol, Lessens Obesity-Associated Inflammation, and Helps to Prevent Hepatic Steatosis in Obese Db/Db Mouse. Food Chem. 2019, 287, 107–114. [Google Scholar] [CrossRef]
- Yoon, B.H.; Jung, J.W.; Lee, J.-J.; Cho, Y.-W.; Jang, C.-G.; Jin, C.; Oh, T.H.; Ryu, J.H. Anxiolytic-like effects of sinapic acid in mice. Life Sci. 2007, 81, 234–240. [Google Scholar] [CrossRef]
- Graf, B.L.; Rojo, L.E.; Delatorre-Herrera, J.; Poulev, A.; Calfio, C.; Raskin, I. Phytoecdysteroids and Flavonoid Glycosides among Chilean and Commercial Sources of Chenopodium quinoa: Variation and Correlation to Physico-Chemical Characteristics. J. Sci. Food Agric. 2016, 96, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Gorelick-Feldman, J.; MacLean, D.; Ilic, N.; Poulev, A.; Lila, M.A.; Cheng, D.; Raskin, I. Phytoecdysteroids Increase Protein Synthesis in Skeletal Muscle Cells. J. Agric. Food Chem. 2008, 56, 3532–3537. [Google Scholar] [CrossRef]
- Isenmann, E.; Ambrosio, G.; Joseph, J.F.; Mazzarino, M.; de la Torre, X.; Zimmer, P.; Kazlauskas, R.; Goebel, C.; Botrè, F.; Diel, P.; et al. Ecdysteroids as Non-Conventional Anabolic Agent: Performance Enhancement by Ecdysterone Supplementation in Humans. Arch. Toxicol. 2019, 93, 1807–1816. [Google Scholar] [CrossRef]
- Franco, R.R.; de Almeida Takata, L.; Chagas, K.; Justino, A.B.; Saraiva, A.L.; Goulart, L.R.; de Melo Rodrigues Ávila, V.; Otoni, W.C.; Espindola, F.S.; da Silva, C.R. A 20-Hydroxyecdysone-Enriched Fraction from Pfaffia glomerata (Spreng.) Pedersen Roots Alleviates Stress, Anxiety, and Depression in Mice. J. Ethnopharmacol. 2021, 267, 113599. [Google Scholar] [CrossRef]
- Baev, A.Y.; Charishnikova, O.S.; Khasanov, F.A.; Nebesnaya, K.S.; Makhmudov, A.R.; Rakhmedova, M.T.; Khushbaktova, Z.A.; Syrov, V.N.; Levitskaya, Y.V. Ecdysterone Prevents Negative Effect of Acute Immobilization Stress on Energy Metabolism of Rat Liver Mitochondria. J. Steroid Biochem. Mol. Biol. 2022, 219, 106066. [Google Scholar] [CrossRef]
- Gholipour, P.; Komaki, A.; Ramezani, M.; Parsa, H. Effects of the Combination of High-Intensity Interval Training and Ecdysterone on Learning and Memory Abilities, Antioxidant Enzyme Activities, and Neuronal Population in an Amyloid-β-Induced Rat Model of Alzheimer’s Disease. Physiol. Behav. 2022, 251, 113817. [Google Scholar] [CrossRef]
- OECD. Test No. 425: Acute Oral Toxicity: Up-and-Down Procedure; OECD Publishing: Paris, France, 2008. [Google Scholar] [CrossRef]
- El Hazzam, K.; Hafsa, J.; Sobeh, M.; Mhada, M.; Taourirte, M.; El Kacimi, K.; Yasri, A. An Insight into Saponins from Quinoa (Chenopodium quinoa Willd.): A Review. Molecules 2020, 25, 1059. [Google Scholar] [CrossRef] [Green Version]
- Boncler, M.; Golanski, J.; Lukasiak, M.; Redzynia, M.; Dastych, J.; Watala, C. A new approach for the assessment of the toxicity of polyphenol-rich compounds with the use of high content screening analysis. PLoS ONE 2017, 12, e0180022. [Google Scholar] [CrossRef] [Green Version]
- Lin, B.; Qi, X.; Fang, L.; Zhao, L.; Zhang, R.; Jing, J.; Zhang, S.; Yang, X.; Hou, Z.; Xue, P. In vivo acute toxicity and mutagenic analysis of crude saponins from Chenopodium quinoa Willd. husks. RSC Adv. 2021, 11, 4829–4841. [Google Scholar] [CrossRef]
- Mennen, L.I.; Walker, R.; Bennetau-Pelissero, C.; Scalbert, A. Risks and safety of polyphenol consumption. Am. J. Clin. Nutr. 2005, 81, 326S–329S. [Google Scholar] [CrossRef] [Green Version]
- Zorin, S.N.; Sidorova, Y.S.; Petrov, N.A.; Perova, I.B.; Malinkin, A.D.; Bokov, D.O.; Bessonov, V.V.; Mazo, V.K. A New Functional Food Ingredient Enriched by Phytoecdisteroids and Polyphenols from Quinoa Grains (Chenopodium quinoa Willd.). Res. J. Pharm. Technol. 2021, 14, 4321–4328. [Google Scholar] [CrossRef]
- Flandreau, E.I.; Tóth, M.A. Animal Models of PTSD: A Critical Review. Curr. Top. Behav. Neurosci. 2018, 38, 47–68. [Google Scholar] [CrossRef]
- Verbitsky, A.; Dopfel, D.; Zhang, N. Rodent models of post-traumatic stress disorder: Behavioral assessment. Transl. Psychiatry 2020, 10, 132. [Google Scholar] [CrossRef]
- Poleschuk, T.S.; Sultanov, R.M.; Ermolenko, E.; Shulgina, L.V.; Kasyanov, S.P. Protective action of alkylglycerols under stress. Stress 2019, 23, 213–220. [Google Scholar] [CrossRef]
- Domingues, M.; Casaril, A.M.; Birmann, P.T.; Bampi, S.R.; Lourenço, D.d.A.; Vieira, B.M.; Dapper, L.H.; Lenardão, E.J.; Sonego, M.; Collares, T.; et al. Effects of a selanylimidazopyridine on the acute restraint stress-induced depressive- and anxiety-like behaviors and biological changes in mice. Behav. Brain Res. 2019, 366, 96–107. [Google Scholar] [CrossRef]
- Chen, H.-J.C.; Lee, J.K.; Yip, T.; Sernia, C.; Lavidis, N.A.; Spiers, J.G. Sub-acute restraint stress progressively increases oxidative/nitrosative stress and inflammatory markers while transiently upregulating antioxidant gene expression in the rat hippocampus. Free Radic. Biol. Med. 2019, 130, 446–457. [Google Scholar] [CrossRef] [Green Version]
- Sidorova, Y.S.; Shipelin, V.A.; Petrov, N.A.; Zorin, S.N.; Mazo, V.K. Adaptogenic Properties of a Phytoecdysteroid-Rich Extract from the Leaves of Spinacia oleracea L. Plants 2021, 10, 2555. [Google Scholar] [CrossRef]
- Chaoui, N.; Anarghou, H.; Laaroussi, M.; Essaidi, O.; Najimi, M.; Chigr, F. Long lasting effect of acute restraint stress on behavior and brain anti-oxidative status. AIMS Neurosci. 2022, 9, 57–75. [Google Scholar] [CrossRef]
- Chigr, F.; Rachidi, F.; Tardivel, C.; Najimi, M.; Moyse, E. Modulation of orexigenic and anorexigenic peptides gene expression in the rat DVC and hypothalamus by acute immobilization stress. Front. Cell. Neurosci. 2014, 8, 198. [Google Scholar] [CrossRef] [Green Version]
- Martí, O.; Martí, J.; Armario, A. Effects of chronic stress on food intake in rats: Influence of stressor intensity and duration of daily exposure. Physiol. Behav. 1994, 55, 747–753. [Google Scholar] [CrossRef]
- Das, A.; Rai, D.; Dikshit, M.; Palit, G.; Nath, C. Nature of stress: Differential effects on brain acetylcholinesterase activity and memory in rats. Life Sci. 2005, 77, 2299–2311. [Google Scholar] [CrossRef]
- Chen, X.; He, X.; Sun, J.; Wang, Z. Phytochemical Composition, Antioxidant Activity, α-Glucosidase and Acetylcholinesterase Inhibitory Activity of Quinoa Extract and Its Fractions. Molecules 2022, 27, 2420. [Google Scholar] [CrossRef]
- Thakare, V.N.; Dhakane, V.D.; Patel, B.M. Attenuation of acute restraint stress-induced depressive like behavior and hippocampal alterations with protocatechuic acid treatment in mice. Metab. Brain Dis. 2017, 32, 401–413. [Google Scholar] [CrossRef]
- Hu, J.; Luo, C.X.; Chu, W.H.; Shan, Y.A.; Qian, Z.-M.; Zhu, G.; Yu, Y.B.; Feng, H. 20-Hydroxyecdysone protects against oxidative stress-induced neuronal injury by scavenging free radicals and modulating NF-κB and JNK pathways. PLoS ONE 2012, 7, e50764. [Google Scholar] [CrossRef]
- Reeves, P.G. Components of the AIN-93 Diets as Improvements in the AIN-76A Diet. J. Nutr. 1997, 127, 838S–841S. [Google Scholar] [CrossRef] [Green Version]
- Committee for the Update of the Guide for the Care and Use of Laboratory Animals. Guide Laboratory for the Care and Use of Animals; National Academies Press (US): Washington, DC, USA, 2011; ISBN 978-0-309-15400-0. [Google Scholar]
- Sharanova, N.E.; Kirbaeva, N.V.; Toropygin, I.Y.; Khryapova, E.V.; Koplik, E.V.; Soto, C.K.; Pertsov, S.S.; Vasiliev, A.V. Effect of Acute Emotional Stress on Proteomic Profile of Selected Brain Areas and Lysosomal Proteolysis in Rats with Different Behavioral Activity. Bull. Exp. Biol. Med. 2016, 161, 355–358. [Google Scholar] [CrossRef] [PubMed]
- Pertsov, S.S.; Koplik, E.V.; Kalinichenko, L.S.; Alekseeva, I.V. Effects of Melatonin on Lipid Peroxidation in Blood in Rats with Different Behavioral Characteristics in Acute Emotional Stress. Neurosci. Behav. Physiol. 2016, 46, 133–137. [Google Scholar] [CrossRef]
- Apryatin, S.A.; Shipelin, V.A.; Trusov, N.V.; Mzhelskaya, K.V.; Evstratova, V.S.; Kirbaeva, N.V.; Soto, J.S.; Fesenko, Z.S.; Gainetdinov, R.R.; Gmoshinski, I.V. Comparative analysis of the influence of a high-fat/high-carbohydrate diet on the level of anxiety and neuromotor and cognitive functions in Wistar and DAT-KO rats. Physiol. Rep. 2019, 7, e13987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mzhelskaya, K.V.; Shipelin, V.A.; Shumakova, A.A.; Musaeva, A.D.; Soto, J.S.; Riger, N.A.; Trusov, N.V.; Kirbaeva, N.V.; Apryatin, S.A.; Gmoshinski, I.V. Effects of quercetin on the neuromotor function and behavioral responses of Wistar and Zucker rats fed a high-fat and high-carbohydrate diet. Behav. Brain Res. 2020, 378, 112270. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator | Group | ||
|---|---|---|---|
| CTRL | IMM | IMM-FFI | |
| Body weight, g | 130 ± 3 | 129 ± 4 | 131 ± 3 |
| Time in open arms, s | 49 ± 7 | 51 ± 10 | 40 ± 9 |
| Time in closed arms, s | 225 ± 9 | 214 ± 15 | 237 ± 12 |
| Distance covered, cm | 1103 ± 72 | 1248 ± 144 | 1175 ± 129 |
| Number of transitions | 19 ± 2 | 23 ± 4 | 21 ± 3 |
| Group | First Test CRPA Formation | Second Test after 24 h Short-Term Memory | Third Test after 14 Days Long-Term Memory | |||
|---|---|---|---|---|---|---|
| Latency, s | Entered Animals, % | Latency, s | Number of Animals Entered | Latency, s | Number of Animals Entered | |
| CTRL | 18 ± 3 | 100 | 117 ± 22 | 5 (41%) | 43 ± 15 | 11 (92%) |
| IMM | 27 ± 8 | 100 | 156 ± 16 | 2 (16%) | 75 ± 22 | 8 (67%) |
| IMM-FFI | 23 ± 4 | 100 | 126 ± 23 | 4 (33%) | 88 ± 23 | 7 (58%) |
| Indicator | Group | ||
|---|---|---|---|
| CTRL | IMM | IMM-FFI | |
| HDL, mmol/L | 0.83 ± 0.05 | 0.96 ± 0.05 | 1.00 ± 0.04 1 |
| LDL mmol/L | 0.23 ± 0.02 | 0.23 ± 0.01 | 0.20 ± 0.01 |
| Triglycerides, mmol/L | 1.09 ± 0.12 | 0.90 ± 0.05 | 0.82 ± 0.03 1 |
| Cholesterol, mmol/L | 2.03 ± 0.10 | 2.17 ± 0.08 | 2.20 ± 0.11 |
| AlAT, U/L | 77.8 ± 4.5 | 89.1 ± 3.4 | 75.6 ± 2.6 2 |
| AsAT, U/L | 185.1 ± 17.5 | 131.2 ± 23.9 | 211.0 ± 14.3 2 |
| Total bilirubin, mmol/L | 4.73 ± 0.31 | 6.07 ± 0.50 1 | 5.57 ± 0.52 |
| Total protein, g/L | 65.7 ± 0.9 | 65.0 ± 0.5 | 63.7 ± 0.6 |
| Phosphorus, mmol/L | 2.90 ± 0.05 | 2.96 ± 0.06 | 2.92 ± 0.05 |
| Calcium, mmol/L | 2.64 ± 0.02 | 2.61 ± 0.02 | 2.63 ± 0.02 |
| Magnesium, mmol/L | 0.96 ± 0.03 | 1.01 ± 0.04 | 0.96 ± 0.03 |
| Alkaline phosphatase, U/L | 342 ± 25.3 | 349 ± 20.6 | 391 ± 35.1 |
| Urea, mmol/L | 5.25 ± 0.11 | 5.06 ± 0.07 | 5.05 ± 0.12 |
| Uric acid, mmol/L | 86.29 ± 6.1 | 98.27 ± 4.4 | 87.09 ± 4.3 |
| Creatinine, mmol/L | 25.04 ± 0.65 | 26.41 ± 1.00 | 25.37 ± 0.68 |
| Glucose, mmol/L | 5.29 ± 0.45 | 5.06 ± 0.20 | 5.93 ± 0.16 2 |
| Corticosterone, ng/mL | 19.2 ± 4.4 | 48.2 ± 10.5 1 | 56.4 ± 9.4 1 |
| Indicator | Group | ||
|---|---|---|---|
| CTRL | IMM | IMM-FFI | |
| Calcium, µg/day | 5.5 ± 1.2 | 5.8 ± 1.8 | 4.9 ± 0.7 |
| Creatinine, mg/day | 44.2 ± 3.3 | 43.5 ± 2.1 | 44.7 ± 1.7 |
| Magnesium, µg/day | 27.0 ± 2.8 | 35.2 ± 2.7 | 33.7 ± 3.1 |
| Uric acid, mg/day | 21.9 ± 1.7 | 23.8 ± 1.4 | 22.2 ± 0.9 |
| Urea, mg/day | 3.9 ± 0.7 | 5.3 ± 0.5 | 5.1 ± 0.3 |
| Phosphorus, µg/day | 560.5 ± 38.7 | 695.5 ± 40.1 1 | 709.4 ± 46.2 1 |
| Prostaglandin E2, ng/day | 20.1 ± 2.0 | 19.3 ± 3.3 | 19.9 ± 2.5 |
| Indicator | Group | ||
|---|---|---|---|
| CTRL | IMM | IMM-FFI | |
| Malondialdehyde, mg/mL | 9.7 ± 1.5 | 17.2 ± 3.6 1 | 11.4 ± 1.6 |
| Glutathione reductase, ng/mL | 113.2 ± 10.5 | 93.6 ± 7.0 | 96.9 ± 7.3 |
| Superoxide dismutase, units/mL | 1755.7 ± 92.9 | 1997.1 ± 236.5 | 2369.6 ± 225.4 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sidorova, Y.S.; Shipelin, V.A.; Petrov, N.A.; Zorin, S.N.; Mazo, V.K. Anxiolytic and Antioxidant Effect of Phytoecdysteroids and Polyphenols from Chenopodium quinoa on an In Vivo Restraint Stress Model. Molecules 2022, 27, 9003. https://doi.org/10.3390/molecules27249003
Sidorova YS, Shipelin VA, Petrov NA, Zorin SN, Mazo VK. Anxiolytic and Antioxidant Effect of Phytoecdysteroids and Polyphenols from Chenopodium quinoa on an In Vivo Restraint Stress Model. Molecules. 2022; 27(24):9003. https://doi.org/10.3390/molecules27249003
Chicago/Turabian StyleSidorova, Yuliya S., Vladimir A. Shipelin, Nikita A. Petrov, Sergey N. Zorin, and Vladimir K. Mazo. 2022. "Anxiolytic and Antioxidant Effect of Phytoecdysteroids and Polyphenols from Chenopodium quinoa on an In Vivo Restraint Stress Model" Molecules 27, no. 24: 9003. https://doi.org/10.3390/molecules27249003
APA StyleSidorova, Y. S., Shipelin, V. A., Petrov, N. A., Zorin, S. N., & Mazo, V. K. (2022). Anxiolytic and Antioxidant Effect of Phytoecdysteroids and Polyphenols from Chenopodium quinoa on an In Vivo Restraint Stress Model. Molecules, 27(24), 9003. https://doi.org/10.3390/molecules27249003