
Antioxidant Potential and Enhancement of Bioactive Metabolite Production in In Vitro Cultures of Scutellaria lateriflora L. by Biotechnological Methods



Abstract
:1. Introduction
2. Results and Discussion
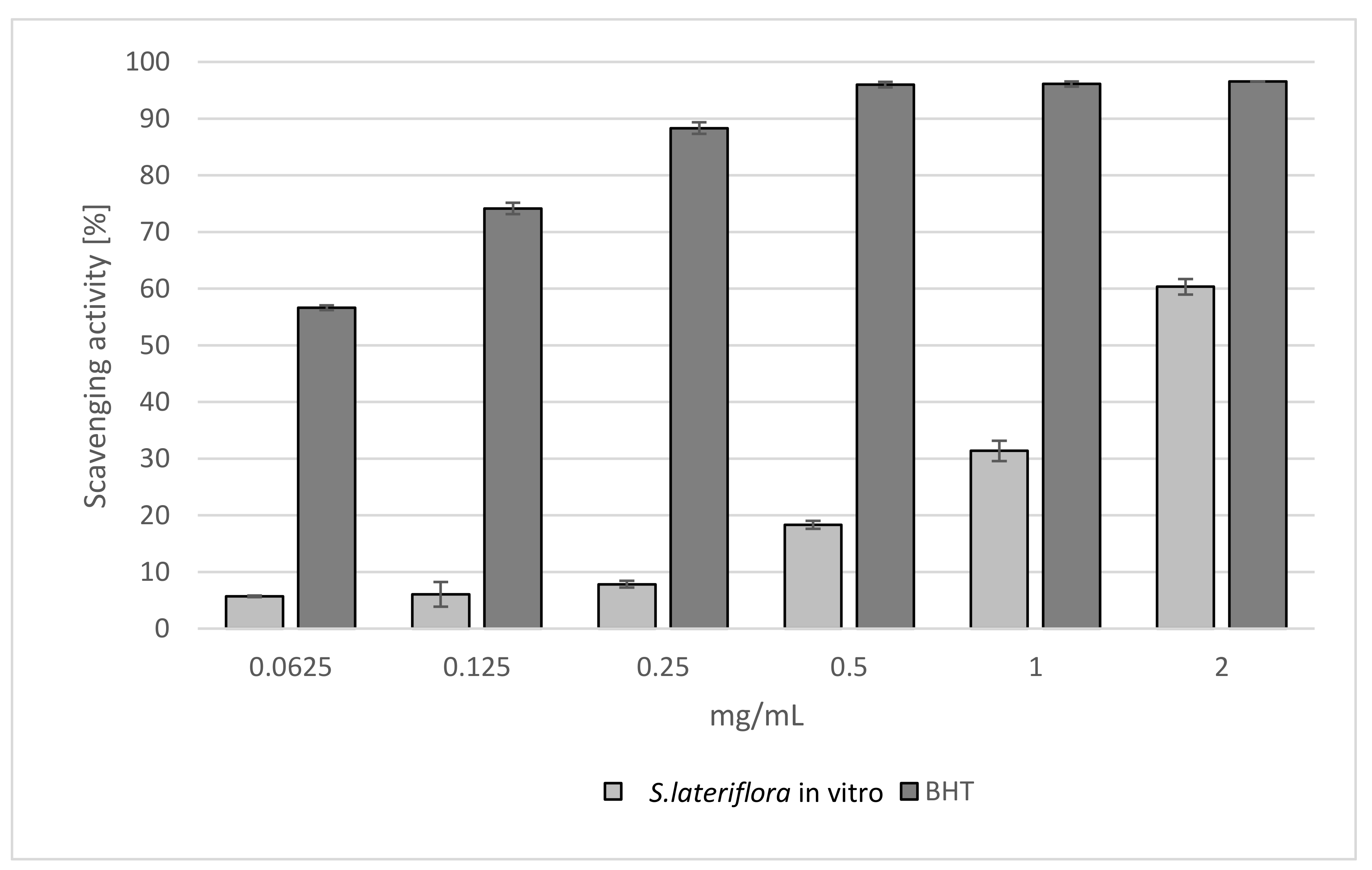
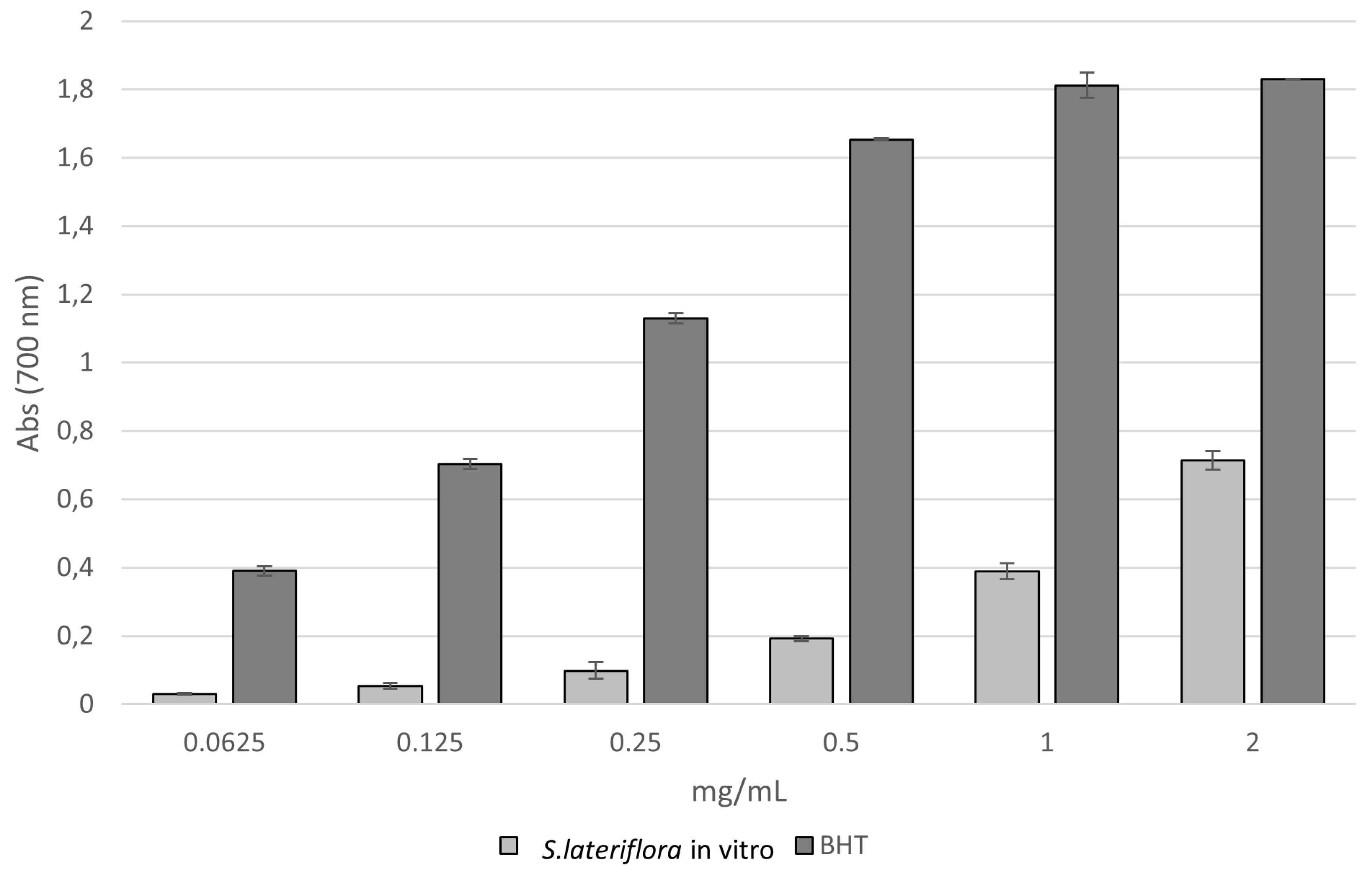
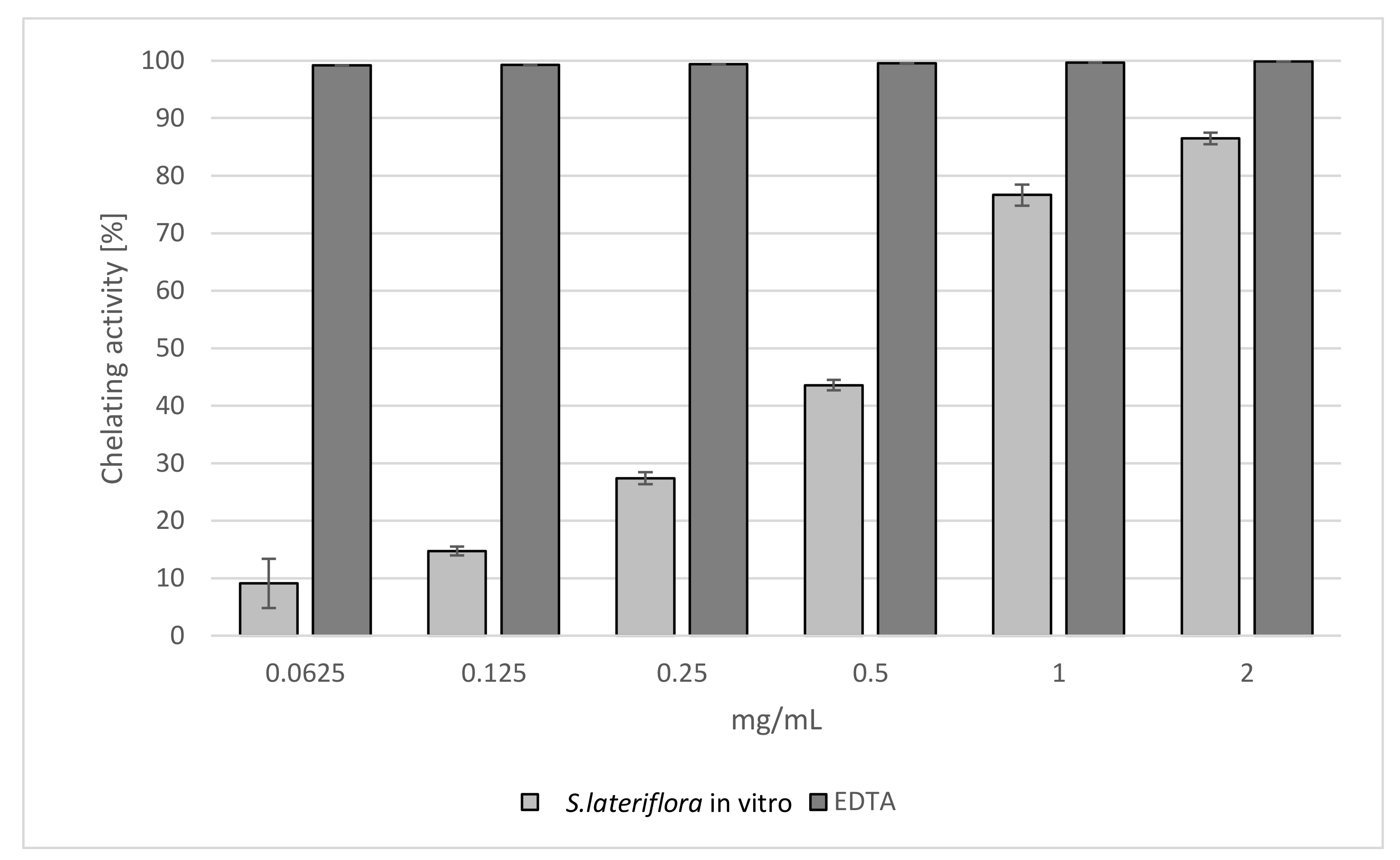
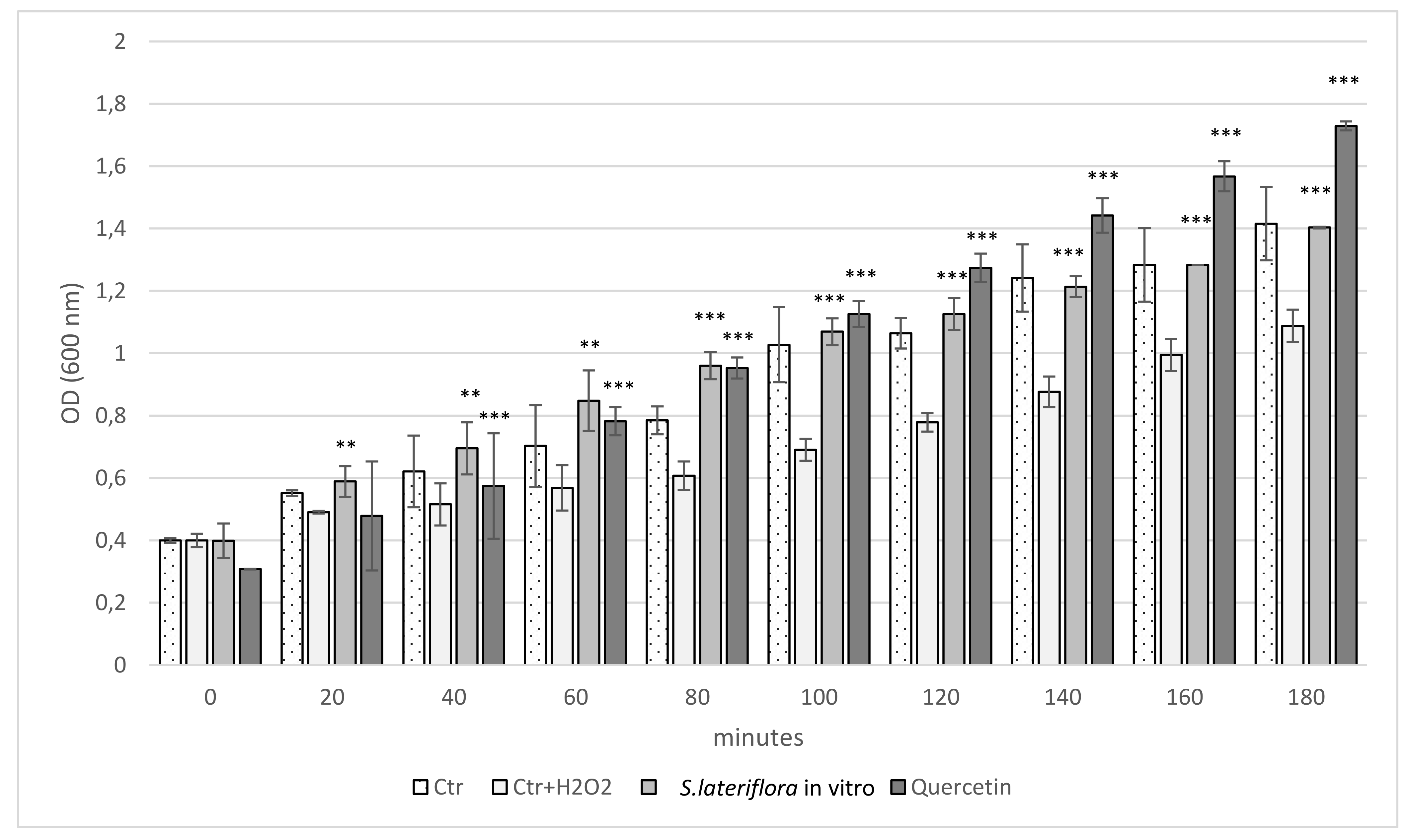
2.1. Antioxidant Activity
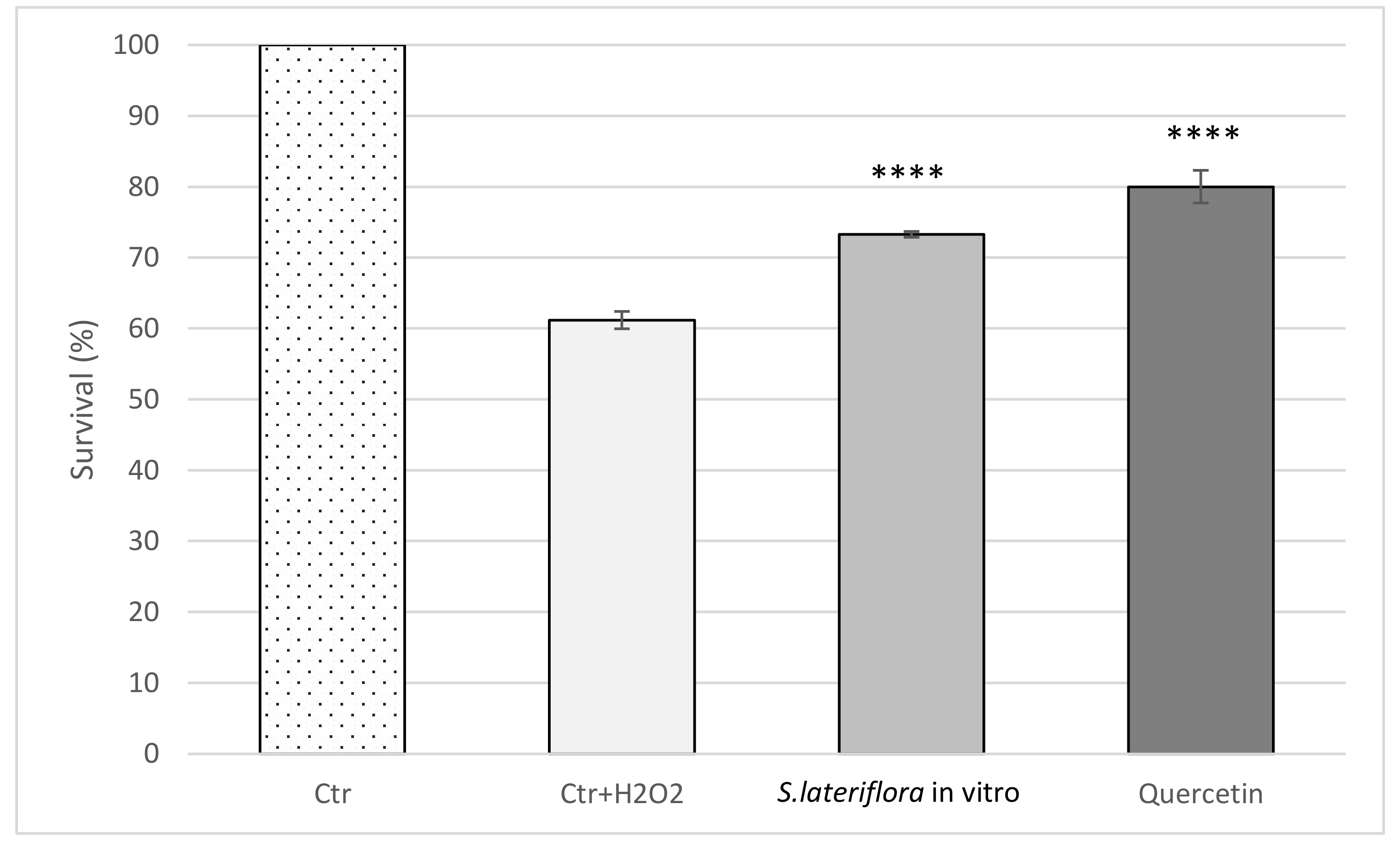
2.2. Artemia salina Lethality Bioassay
2.3. Enhanncing the Production of Secondary Metabolites
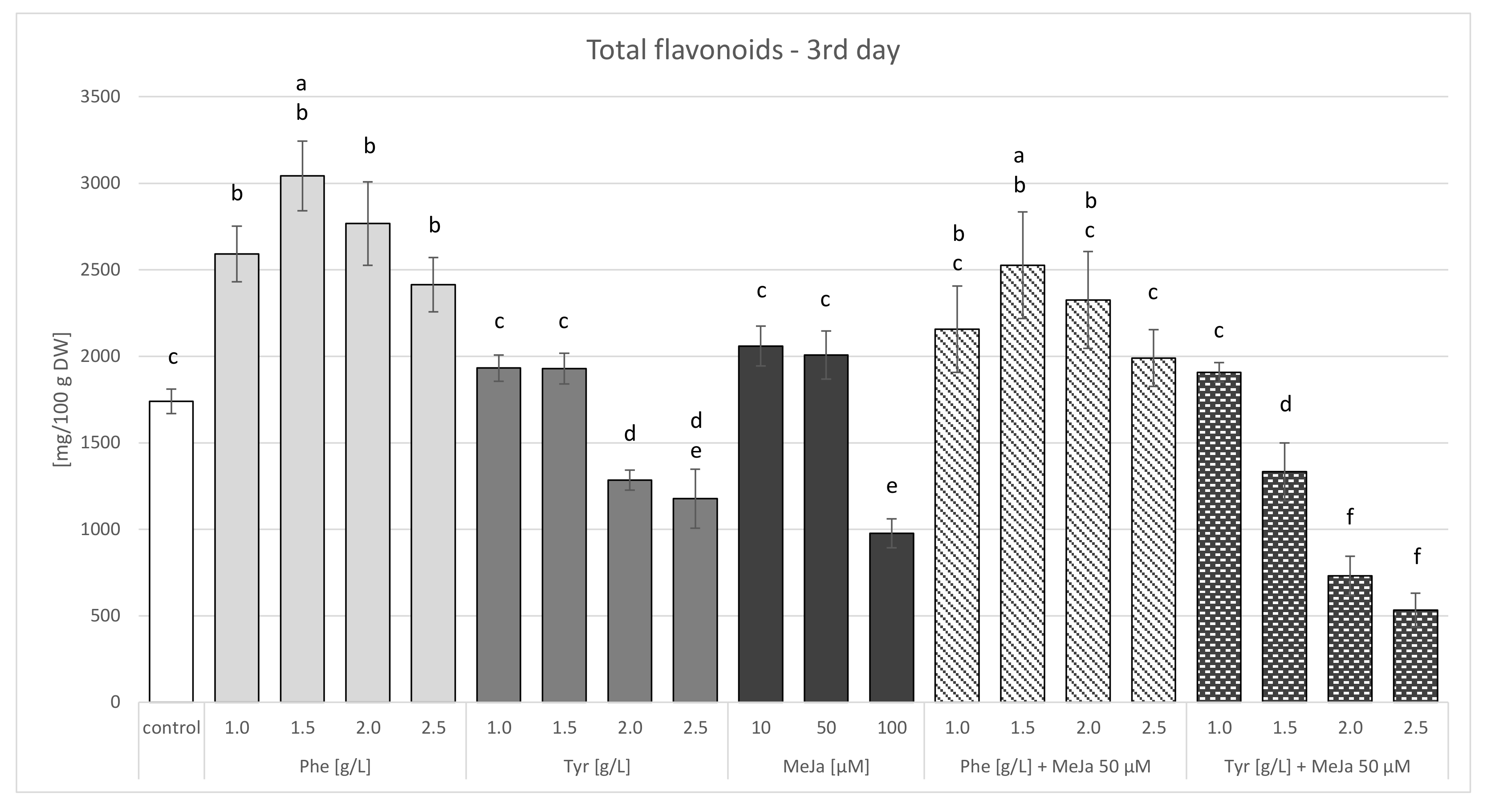
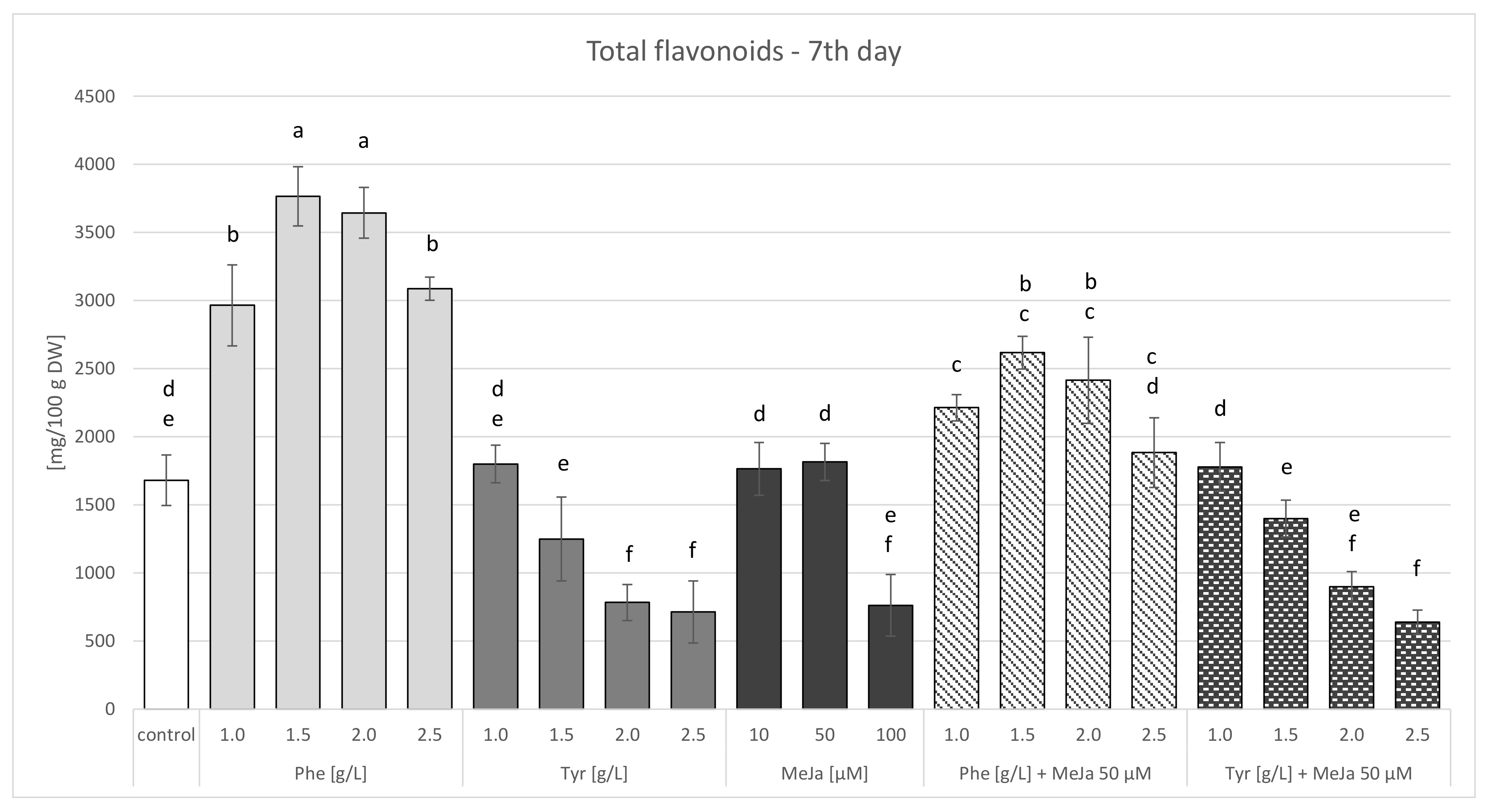
2.3.1. Agar Culture Feeding with Biosynthetic Precursors
2.3.2. Agitated Cultures
Feeding with Biosynthetic Precursors
Elicitation with Methyl Jasmonate
Combined Strategy: Feeding with Biosynthetic Precursors and Elicitation
2.3.3. Cultures in Bioreactors
3. Materials and Methods
3.1. In Vitro Cultures
3.1.1. Agar Cultures
3.1.2. Agitated Cultures
3.1.3. Cultures in Bioreactors
3.2. Reverse-Phase High-Performance Liquid Chromatography (RP-HPLC) Analysis
3.3. Total Phenolic and Flavonoid Content
3.4. Antioxidant Activity
3.4.1. Free Radical Scavenging Activity
3.4.2. Reducing-Power Assay
3.4.3. Ferrous Ions (Fe2+) Chelating Activity
3.4.4. Protective Effect on Escherichia coli Growth and Survival under Peroxide Stress
3.5. Brine Shrimp (Artemia salina) Lethality Bioassay
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| ASE | ascorbic acid equivalent |
| BA | 6-benzyladenine |
| BHT | butylated hydroxytoluene |
| Ctr | control |
| DPPH | 1,1-diphenyl-2-picrylhydrazyl |
| DW | dry weight |
| GAE | gallic acid equivalents |
| HAT | hydrogen-atom transfer |
| LS | Linsmaier and Skoog |
| MeJa | methyl jasmonate |
| MS | Murashige and Skoog |
| NAA | 1-naphthaleneacetic acid |
| PGR | plant growth regulator |
| QE | quercetin equivalents |
| SET | single-electron transfer |
| TIS | temporary immersion system |
| TPC | total phenolic content |
| TFC | total flavonoid content |
References
- Shen, J.; Li, P.; Liu, S.; Liu, Q.; Li, Y.; Sun, Y.; He, C.; Xiao, P. Traditional uses, ten-years research progress on phytochemistry and pharmacology, and clinical studies of the genus Scutellaria. J. Ethnopharmacol. 2021, 265, 113198. [Google Scholar] [CrossRef]
- Millspaugh, C.F. Scutellaria. American Medicinal Plants; Dover Publications: New York, NY, USA, 1974; pp. 469–472. [Google Scholar]
- Upton, R.; DAyu, R.H. Skullcap Scutellaria lateriflora L.: An American nervine. J. Herb. Med. 2012, 2, 76–96. [Google Scholar] [CrossRef]
- Liu, W.; Ge, T.; Pan, Z.; Leng, Y.; Lv, J.; Li, B. The effects of herbal medicine on epilepsy. Oncotarget 2017, 8, 48385–48397. Available online: https://www.oncotarget.com/article/16801/ (accessed on 10 December 2021). [CrossRef] [Green Version]
- Sarris, J. Herbal medicines in the treatment of psychiatric disorders: 10-year updated review. Phytother. Res. 2018, 32, 1147–1162. [Google Scholar] [CrossRef]
- Zhang, Z.; Lian, X.-Y.; Li, S.; Stringer, J.L. Characterization of chemical ingredients and anticonvulsant activity of American skullcap (Scutellaria lateriflora). Phytomedicine 2009, 16, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Upton, R.; Graff, A.; Jolliffe, G.; Länger, R. Skullcap aerial parts. In American Herbal Pharmacopoeia: Botanical Pharmacognosy-Microscopic Characterization of Botanical Medicines; CRC Press: Boca Raton, FL, USA, 2011; pp. 599–603. [Google Scholar]
- Awad, R.; Arnason, J.T.; Trudeau, V.; Bergeron, C.; Budzinski, J.W.; Foster, B.C.; Merali, Z. Phytochemical and biological analysis of skullcap (Scutellaria lateriflora L.): A medicinal plant with anxiolytic properties. Phytomedicine 2003, 10, 640–649. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.-H.; Smillie, T.J.; Khan, I.A. Identification of phenolic compounds from Scutellaria lateriflora by liquid chromatography with ultraviolet photodiode array and electrospray ionization tandem mass spectrometry. J. Pharm. Biomed. Anal. 2012, 63, 120–127. [Google Scholar] [CrossRef]
- Bruno, M.; Cruciata, T.M.; Bondi, M.L.; Piozzi, F.; De La Torre, M.C.; Rodriguez, B.; Servettaz, O. Neo-clerodane diterpenoids from Scutellaria lateriflora. Phytochemistry 1998, 48, 687–691. [Google Scholar] [CrossRef]
- Ramawat, K.G.; Mathur, M. Factors affecting the production of secondary metabolites. In Biotechnology: Secondary Metabolites. Plants and Microbes; Ramawat, K.G., Merillon, J.M., Eds.; Science Publishers Inc.: Enfield, NH, USA, 2007; pp. 59–102. [Google Scholar]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res. 2009, 3, 1222–1239. [Google Scholar] [CrossRef]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.M.; Palazon, J. Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef]
- Shah, H.; Mtewa, A.G.; Egbuna, C.; Godwin, A.; Sensaazi, D.C. Biotechnology approach to the production of phytochemicals: An introduction. In Phytochemistry. Marine Sources, Industrial Applications, and Recent Advances; Egbuna, C., Ifemeje, J.C., Kumar, S., Sharif, N., Eds.; Apple Academic Press: Oakville, ON, Canada, 2018; Volume 3, pp. 107–130. [Google Scholar]
- Marsh, Z.; Yang, T.; Nopo-Olazabal, L.; Wu, S.; Ingle, T.; Joshee, N.; Medina-Bolivar, F. Effect of light, methyl jasmonate and cyclodextrin on production of phenolic compounds in hairy root cultures of Scutellaria lateriflora. Phytochemistry 2014, 107, 50–60. [Google Scholar] [CrossRef]
- Kwon, D.Y.; Kim, H.H.; Park, J.S.; Park, S.U.; Park, N.I. Production of Baicalin, Baicalein and Wogonin in Hairy Root Culture of American Skullcap (Scutellaria lateriflora) by Auxin Treatment. Biosci. Biotechnol. Res. Asia 2017, 14, 673–677. [Google Scholar] [CrossRef]
- Stepanova, A.Y.; Solov’eva, A.I.; Malunova, M.V.; Salamaikina, S.A.; Panov, Y.M.; Lelishentsev, A.A. Hairy Roots of Scutellaria spp. (Lamiaceae) as Promising Producers of Antiviral Flavones. Molecules 2021, 26, 3927. [Google Scholar] [CrossRef]
- Cole, I.B.; Farooq, F.T.; Murch, S.J. Protocols for establishment of an in vitro collection of medicinal plants in the genus Scutellaria. In Protocols for In Vitro Cultures and Secondary Metabolite Analysis of Aromatic and Medicinal Plants. Methods in Molecular Biology; Jain, S.M., Saxena, P.K., Eds.; Humana Press: Totawa, NJ, USA, 2009; Volume 547, pp. 155–165. [Google Scholar]
- Tascan, A.; Adelberg, J.; Tascan, M.; Rimando, J.; Yadav, A.K. Hyperhydricity and flavonoid content of Scutellaria species in vitro on polyester-supported liquid culture system. Hortscience 2010, 45, 1723–1728. [Google Scholar] [CrossRef] [Green Version]
- Kawka, B.; Kwiecień, I.; Ekiert, H. Endogenous production of specific flavonoids and verbascoside in agar and agitated microshoot cultures of Scutellaria lateriflora L. and biotransformation potential. Plant Cell Tiss Organ Cult. 2020, 142, 471–482. [Google Scholar] [CrossRef]
- Kawka, B.; Kwiecień, I.; Ekiert, H. Influence of culture medium composition and light conditions on the accumulation of bioactive compounds in shoot cultures of Scutellaria lateriflora L. (American Skullcap) grown in vitro. Appl. Biochem. Biotechnol. 2017, 183, 1414–1425. [Google Scholar] [CrossRef] [Green Version]
- Kawka, B.; Kwiecień, I.; Ekiert, H. Production of specific flavonoids and verbascoside in shoot cultures of Scutellaria baicalensis. In Plant Cell and Tissue Differentiation and Secondary Metabolites: Reference Series in Phytochemistry; Ramawat, K., Ekiert, H., Goyal, S., Eds.; Springer Nature: Geneva, Switzerland, 2021; pp. 249–272. [Google Scholar] [CrossRef]
- Nabi, N.; Singh, S.; Saffeullah, P. Responses of in vitro cell cultures to elicitation: Regulatory role of jasmonic acid and methyl jasmonate: A review. In Vitro Cell Dev. Biol. Plant 2021, 57, 341–355. [Google Scholar] [CrossRef]
- Perez, C.A.; Wei, Y.; Guo, M. Iron-binding and anti-Fenton properties of baicalein and baicalin. J. Inorg. Biochem. 2009, 103, 326–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Ge, R.C.; Zheng, R.L.; Liu, Z.M.; Jia, Z.J. Antioxidative and chelating activities of phenylpropanoid glycosides from Pedicularis striata. Acta Pharmacol. Sin. 1997, 18, 77–80. [Google Scholar]
- Wang, Z.L.; Wang, S.; Kuang, Y.; Hu, Z.M.; Qiao, X.; Ye, M. A comprehensive review on phytochemistry, pharmacology, and flavonoid biosynthesis of Scutellaria baicalensis. Pharm. Biol. 2018, 56, 465–484. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.; Ye, J.; Gao, L.; Liu, Y. The main bioactive compounds of Scutellaria baicalensis Georgi. for alleviation of inflammatory cytokines: A comprehensive review. Biomed. Pharmacother. 2021, 133, 110917. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; He, X.; He, X.; Li, M.; Zhang, R.; Fan, P.; Zhang, Q.; Jia, Z. The genus Scutellaria an ethnopharmacological and phytochemical review. J. Ethnopharmacol. 2010, 128, 279–313. [Google Scholar] [CrossRef]
- Korkina, L.G. Phenylpropanoids as naturally occurring antioxidants: From plant defense to human health. Cell. Mol. Biol. 2007, 53, 15–25. [Google Scholar]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside—A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef]
- Wu, L.; Georgiev, M.I.; Cao, H.; Nahar, L.; El-Seedi, H.R.; Sarker, S.D.; Xiao, J.; Lu, B. Therapeutic potential of phenylethanoid glycosides: A systematic review. Med. Res. Rev. 2020, 40, 2605–2649. [Google Scholar] [CrossRef]
- Yu, D.; Pu, W.; Li, D.; Wang, D.; Liu, Q.; Wang, Y. Phenolic Compounds and Antioxidant Activity of Different Organs of Potentilla fruticosa L. from Two Main Production Areas of China. Chem. Biodivers. 2016, 13, 1140–1148. [Google Scholar] [CrossRef]
- Oktyabrsky, O.; Vysochina, G.; Muzyka, N.; Samoilova, Z.; Kukushkina, T.; Smirnova, G. Assessment of antioxidant activity of plant extracts using microbial test systems. J. Appl. Microbiol. 2009, 106, 1175–1183. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, G.V.; Samoylova, Z.Y.; Muzyka, N.G.; Oktyabrsky, O.N. Influence of polyphenols on Escherichia coli resistance to oxidative stress. Free Radic. Biol. Med. 2009, 46, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Pu, W.; Wang, D.; Zhou, D. Structural characterization and evaluation of the antioxidant activity of phenolic compounds from Astragalus taipaishanensis and their structure activity relationship. Sci. Rep. 2015, 5, 13914. [Google Scholar] [CrossRef] [Green Version]
- Rajabi, S.; Ramazani, A.; Hamidi, M.; Naji, T. Artemia salina as a model organism in toxicity assessment of nanoparticles. DARU J. Pharm. Sci. 2015, 23, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagarto Parra, A.; Silva Yhebra, R.; Guerra Sardiñas, I.; Iglesias Buela, L. Comparative study of the assay of Artemia salina L. and the estimate of the medium lethal dose (LD50 value) in mice, to determine oral acute toxicity of plant extracts. Phytomedicine 2001, 8, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Dziurka, M.; Kubica, P.; Kwiecień, I.; Biesaga-Kościelniak, J.; Ekiert, H.; Abdelmohsen, S.A.M.; Al-Harbi, F.F.; El-Ansary, D.O.; Elansary, H.O.; Szopa, A. In Vitro Cultures of Some Medicinal Plant Species (Cistus × incanus, Verbena officinalis, Scutellaria lateriflora, and Scutellaria baicalensis) as a Rich Potential Source of Antioxidants—Evaluation by CUPRAC and QUENCHER-CUPRAC Assays. Plants 2021, 10, 454. [Google Scholar] [CrossRef] [PubMed]
- El-Shennawy, O.A.; El-Torky, G.M.; El-Mokadem, E.H.; Abass, B.I. Effect of NaCl and Phenylalanine on the Production of some Secondary Metabolites in In Vitro Cultures of Mentha longifolia. Alex. Sci. Exch. J. 2017, 38, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Szopa, A.; Kubica, P.; Komsta, Ł.; Walkowicz-Bożek, A.; Ekiert, H. The effect of feeding culture media with biogenetic precursors on high production of depsides in agitated shoot cultures of black and red aronias. Plant Cell Tissue Organ Cult. 2020, 142, 379–399. [Google Scholar] [CrossRef]
- Herrman, K.M. The shikimate pathway: Early step in the Biosynthesis of Aromatic Compounds. Plant Cell 1995, 7, 907–919. [Google Scholar] [CrossRef]
- Falcone-Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sajjalaguddam, R.R.; Paladugu, A. Phenylalanine enhances Quercetin content in In vitro cultures of Abutilon indicum L. J. Appl. Pharm. Sci. 2015, 5, 80–84. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, A.; Kwiecień, I.; Grabowski, M.; Rajek, K.; Cavo, E.; Taviano, M.F.; Miceli, N. Phenylalanine increases the production of antioxidant phenolic acids in Ginkgo biloba cell cultures. Molecules 2021, 26, 4965. [Google Scholar] [CrossRef]
- Arano-Varela, H.; Cruz-Sosa, F.; Estrada-Zúñiga, M.E.; Fernández, F.J. Effects of phenylalanine and methyl jasmonate on verbascoside production in Buddleja cordata Kunth cell suspension cultures. S. Afr. J. Bot. 2020, 135, 41–49. [Google Scholar] [CrossRef]
- Biswas, T.; Mathur, A.; Gupta, V.; Luqman, S.; Mathur, A.K. Elicitation and phenylalanine precursor feeding based modulation of in vitro anthocyanin production, enzyme activity and gene expression in an Indian ginseng congener-Panax sikkimensis Ban. Ind. Crops Prod. 2020, 145, 111986. [Google Scholar] [CrossRef]
- Edahiro, J.-I.; Nakamura, M.; Seki, M.; Furusaki, S. Enhanced accumulation of anthocyanin in cultured strawberry cells by repetitive feeding of l-phenylalanine into the medium. J. Biosci. Bioeng. 2005, 99, 43–47. [Google Scholar] [CrossRef]
- Skrzypczak-Pietraszek, E.; Słota, J.; Pietraszek, J. The influence of L-phenylalanine, methyl jasmonate and sucrose concentration on the accumulation of phenolic acids in Exacum affine Balf. f. ex Regel shoot culture. Acta Biochim. Pol. 2014, 61, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Klimek-Szczykutowicz, M.; Dziurka, M.; Blažević, I.; Đulović, A.; Miazga-Karska, M.; Klimek, K.; Ekiert, H.; Szopa, A. Precursor-Boosted Production of Metabolites in Nasturtium officinale Microshoots Grown in Plantform Bioreactors, and Antioxidant and Antimicrobial Activities of Biomass Extracts. Molecules 2021, 26, 4660. [Google Scholar] [CrossRef] [PubMed]
- Mathew, R.; Sankar, P.D. Effect of methyl jasmonate and chitosan on growth characteristics of Ocimum basilicum L., Ocimum sanctum L. and Ocimum gratissimum L. cell suspension cultures. Afr. J. Biotechnol. 2012, 11, 4759–4766. [Google Scholar] [CrossRef]
- Thakur, M.; Bhattacharya, S.; Kumar Khosla, P.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Fatemi, F.; Abdollahi, M.R.; Mirzaie-asl, A.; Dastan, D.; Garagounis, C.; Papadopoulou, K. Identification and expression profiling of rosmarinic acid biosynthetic genes from Satureja khuzistanica under carbon nanotubes and methyl jasmonate elicitation. Plant Cell Tissue Organ Cult. 2019, 136, 561–573. [Google Scholar] [CrossRef]
- Krzyzanowska, J.; Czubacka, A.; Pecio, L.; Przybys, M. The effects of jasmonic acid and methyl jasmonate on rosmarinic acid production in Mentha × piperita cell suspension cultures. Plant Cell Tissue Organ Cult. 2012, 108, 73–81. [Google Scholar] [CrossRef]
- Mendoza, D.; Cuaspud, O.; Arias, J.P.; Ruiz, O.; Arias, M. Effect of salicylic acid and methyl jasmonate in the production of phenolic compounds in plant cell suspension cultures of Thevetia peruviana. Biotechnol. Rep. 2018, 19, e00273. [Google Scholar] [CrossRef]
- Tassoni, A.; Durante, L.; Ferri, M. Combined elicitation of methyl-jasmonate and red light on stilbene and anthocyanin biosynthesis. J. Plant Physiol. 2012, 169, 775–781. [Google Scholar] [CrossRef]
- Saeed, S.; Ali, H.; Khan, T.; Kayani, W.; Khan, M.A. Impacts of methyl jasmonate and phenyl acetic acid on biomass accumulation and antioxidant potential in adventitious roots of Ajuga bracteosa (Wall ex Benth) a high valued endangered medicinal plant. Physiol. Mol. Biol. Plants 2017, 23, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Shakya, P.; Marslin, B.; Siram, K.; Beerhues, L.; Franklin, G. Elicitation as a tool to improve the profiles of high-value secondary metabolites and pharmacological properties of Hypericum perforatum. J. Pharm. Pharmacol. 2017, 71, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Klimek-Szczykutowicz, M.; Dziurka, M.; Blažević, I.; Đulović, A.; Apola, A.; Ekiert, H.; Szopa, A. Impacts of elicitors on metabolite production and on antioxidant and tyrosinase inhibition in watercress microshoot cultures. Appl. Microbiol. Biotechnol. 2022, 106, 619–633. [Google Scholar] [CrossRef]
- Szopa, A.; Kokotkiewicz, A.; Król, A.; Luczkiewicz, M.; Ekiert, H. Improved production of dibenzocyclooctadiene lignans in the elicited microshoot cultures of Schisandra chinensis (Chinese magnolia vine). Appl. Microbiol. Biotechnol. 2018, 102, 945–959. [Google Scholar] [CrossRef] [Green Version]
- Towler, M.J.; Kim, Y.; Wyslouzil, B.E.; Correll, M.J.; Weathers, P.J. Design, development, and applications of mist bioreactors for micropropagation and hairy root culture. In Plant Tissue Culture Engineering; Gupta, S.D., Ibaraki, Y., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 119–134. [Google Scholar] [CrossRef]
- Paek, K.Y.; Chakrabarty, D.; Hahn, E.J. Application of bioreactor systems for large scale production of horticultural and medicinal plants. Plant Cell Tissue Organ Cult. 2005, 81, 287–300. [Google Scholar] [CrossRef]
- Szopa, A.; Kokotkiewicz, A.; Bednarz, M.; Jafernik, K.; Luczkiewicz, M.; Ekiert, H. Bioreactor type affects the accumulation of phenolic acids and flavonoids in microshoot cultures of Schisandra chinensis (Turcz.) Baill. Plant Cell Tissue Organ Cult. 2019, 139, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Kubica, P.; Szopa, A.; Kokotkiewicz, A.; Miceli, N.; Taviano, M.F.; Maugeri, A.; Cirmi, S.; Synowiec, A.; Gniewosz, M.; Elansary, H.O.; et al. Production of Verbascoside, Isoverbascoside and Phenolic Acids in Callus, Suspension, and Bioreactor Cultures of Verbena officinalis and Biological Properties of Biomass Extracts. Molecules 2020, 25, 5609. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Linsmaier, E.M.; Skoog, F. Organic growth factor requirements of tobacco tissue cultures. Physiol. Plant 1965, 18, 100–127. [Google Scholar] [CrossRef]
- Sułkowska-Ziaja, K.; Maślanka, A.; Szewczyk, A.; Muszyńska, B. Physiologically active compounds in four species of Phellinus. Nat. Prod. Commun. 2017, 12, 363–366. [Google Scholar] [CrossRef] [Green Version]
- Taviano, M.F.; Marino, A.; Trovato, A.; Bellinghieri, V.; La Barbera, T.M.; Güvenç, A.; Hürkul, M.M.; De Pasquale, R.; Miceli, N. Antioxidant and antimicrobial activities of branches extracts of five Juniperus species from Turkey. Pharm. Biol. 2011, 49, 1014–1022. [Google Scholar] [CrossRef]
- Ohnishi, M.; Morishita, H.; Iwahashi, H.; Shitzuo, T.; Yoshiaki, S.; Kimura, M.; Kido, R. Inhibitory effects of chlorogenic acid on linoleic acid peroxidation and haemolysis. Phytochemistry 1994, 36, 579–583. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction: Antioxidative activities of products of browning reaction prepared from glucosamine. Jpn J. Nutr. Diet 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Decker, E.A.; Welch, B. Role of ferritin as a lipid oxidation catalyst in muscle food. J. Agric. Food Chem. 1990, 38, 674–677. [Google Scholar] [CrossRef]
- Smirnova, G.V.; Vysochina, G.I.; Muzyka, N.G.; Samoylova, Z.Y.; Kukushkina, T.A.; Oktyabrsky, O.N. Evaluation of antioxidant properties of medical plants using microbial test systems. World J. Microbiol. Biotechnol. 2010, 26, 2269–2276. [Google Scholar] [CrossRef]
- Meyer, B.N.; Ferrigni, N.R.; Putnam, J.E.; Jacobson, L.B.; Nichols, D.E.; McLaughlin, J.L. Brine shrimp: A convenient general bioassay for active plant constituents. Planta Med. 1982, 45, 31–34. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TPC mg GAE/g Extract | TFC mg QE/g Extract | DPPH IC50 [mg/mL] | Reducing Power ASE/mL | Fe2+ Chelating Activity IC50 [mg/mL] | |
|---|---|---|---|---|---|
| S. lateriflora extract | 37.270 ± 0.650 | 8.820 ± 0.900 | 1.639 ± 0.008 a | 43.481 ± 0.237 a | 0.609 ± 0.018 a |
| Reference standard | - | - | BHT 0.066 ± 0.008 b | BHT 1.131 ± 0.008 b | BHT 0.007 ± 0.001 b |
| Metabolite [mg/100 g DW] | Control | Phe | Tyr |
|---|---|---|---|
| Baicalein | 239.05 ± 58.668 | 222.48 ± 49.106 | 124.56 ± 77.470 |
| Baicalin | 1692.15 ± 206.525 a | 1658.94 ± 200.664 a | 996.90 ± 228.960 b |
| Wogonin | 74.82 ± 43.251 a | 95.77 ± 29.820 a,b | 35.38 ± 16.405 b |
| Wogonoside | 711.97 ± 4.835 a | 491.16 ± 67.224 b | 128.29 ± 35.132 c |
| Scutellarin | 4.41 ± 0.270 a | 46.79 ± 7.684 b | 72.84 ± 21.906 b |
| Oroxylin A | 45.75 ± 19.362 a | 14.98 ± 3.887 b | 4.41 ± 2.162 c |
| Total flavonoids | 2768.14 ± 332.911 a | 2530.13 ± 358.385 a | 1262.73 ± 382.035 b |
| Verbascoside | 292.00 ± 65.789 a | 457.82 ± 57.807 b | 553.72 ± 135.302 b |
| Metabolite [mg/100 g DW] | 3rd Day | |||||
|---|---|---|---|---|---|---|
| Control | Phe | Tyr | MeJa | Phe + MeJa | Tyr + MeJa | |
| Baicalein | 171.557 ± 12.516 | 533.084 ± 18.753 b | 315.664 ± 28.169 b | 351.450 ± 41.801 c | 527.640 ± 57.302 b | 257.450 ± 8.652 a |
| Baicalin | 445.713 ± 20.473 | 698.826 ± 35.761 b | 442.068 ± 20.460 b | 552.318 ± 23.506 c | 610.602 ± 78.309 b | 507.194 ± 11.708 a |
| Wogonin | 689.250 ± 24.371 | 808.929 ± 64.103 b | 749.526 ± 15.489 b | 676.638 ± 25.215 d | 745.436 ± 77.691 b | 659.016 ± 10.974 a |
| Wogonoside | 424.741 ± 11.473 | 935.389 ± 67.641 b | 407.018 ± 21.356 a | 469.450 ± 23.055 c | 602.067 ± 85.539 b | 442.457 ± 18.941 a |
| Scutellarin | 4.065 ± 0.927 | 72.340 ± 13.252 a | 14.205 ± 1.264 b | 25.840 ± 6.515 d | 17.999 ± 5.157 b | 17.452 ± 1.758 a |
| Oroxylin A | 4.620 ± 0.788 | 34.814 ± 2.934 b | 29.693 ± 4.480 a | 37.418 ± 2.325 d | 33.493 ± 2.452 a | 27.505 ± 2.540 b |
| Total flavonoids | 1739.947 ± 70.548 | 3041.690 ± 201.914 b | 1931.784 ± 75.987 a | 2059.550 ± 114.897 c | 2525.897 ± 307.879 b | 1905.997 ± 55.977 a |
| Verbascoside | 311.197 ± 40.548 | 469.127 ± 91.914 b | 299.480 ± 75.987 a | 381.523 ± 114.897 c | 160.597 ± 37.879 b | 25.460 ± 7.405 b |
| Metabolite [mg/100 g DW] | 7th Day | |||||
|---|---|---|---|---|---|---|
| Control | Phe | Tyr | MeJa | Phe + MeJa | Tyr + MeJa | |
| Baicalein | 162.414 ± 26.104 | 614.788 ± 38.279 b | 162.457 ± 49.878 b | 300.486 ± 40.901 e | 541.608 ± 36.215 b | 264.457 ± 31.590 a |
| Baicalin | 378.595 ± 26.606 | 988.086 ± 56.083 c | 419.209 ± 21.780 a | 474.399 ± 49.675 d | 630.484 ± 17.532 b | 411.619 ± 41.777 a |
| Wogonin | 666.088 ± 71.518 | 964.501 ± 49.668 b | 655.544 ± 30.779 a | 676.857 ± 55.040 e | 782.986 ± 29.100 b | 599.387 ± 51.315 a |
| Wogonoside | 469.987 ± 59.996 | 1148.771 ± 50.697 c | 516.879 ± 56.240 a | 365.481 ± 17.298 e | 620.747 ± 30.154 b | 479.433 ± 51.315 a |
| Scutellarin | 2.539 ± 0.940 | 68.295 ± 4.980 b | 9.321 ± 1.797 a | 10.087 ± 2.591 e | 18.946 ± 3.460 a | 10.042 ± 1.025 b |
| Oroxylin A | 0.214 ± 0.023 | 53.653 ± 4.450 c | 41.758 ± 8.250 b | 43.708 ± 3.768 d | 25.006 ± 7.145 c | 14.562 ± 2.246 a |
| Total flavonoids | 1679.837 ± 185.187 | 3764.881 ± 217.489 b | 1799.470 ± 137.499 a | 1814.878 ± 137.456 e | 2616.874 ± 120.568 b | 1778.499 ±179.168 a |
| Verbascoside | 267.327 ± 15.187 | 474.795 ± 27.489 b | 263.961 ± 37.499 a | 121.553 ± 19.489 d | 170.694 ± 17.156 a | 44.068 ± 4.892 b |
| Metabolite * [mg/100 g DW] | BA/NAA [mg/L] | |
|---|---|---|
| MS 1.0/0.5 | LS 1.0/0.5 | |
| Baicalin | 1388.74 ± 140.169 a | 2191.23 ±399.847 b |
| Wogonin | 169.22 ± 48.145 a | 68.56 ± 13.553 b |
| Wogonoside | 174.99 ± 61.832 a | 53.67 ± 21.344 b |
| Scutellarin | 10.55 ± 3669 | 16.91 ± 5358 |
| Oroxylin A | 39.87 ± 3835 a | 17.49 ± 8345 b |
| Total flavonoids | 1783.38 ± 257.651 | 2347.87 ± 448.447 |
| Verbascoside | 485.45 ± 47.439 a | 310.37 ± 37.778 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwiecień, I.; Miceli, N.; D’Arrigo, M.; Marino, A.; Ekiert, H. Antioxidant Potential and Enhancement of Bioactive Metabolite Production in In Vitro Cultures of Scutellaria lateriflora L. by Biotechnological Methods. Molecules 2022, 27, 1140. https://doi.org/10.3390/molecules27031140
Kwiecień I, Miceli N, D’Arrigo M, Marino A, Ekiert H. Antioxidant Potential and Enhancement of Bioactive Metabolite Production in In Vitro Cultures of Scutellaria lateriflora L. by Biotechnological Methods. Molecules. 2022; 27(3):1140. https://doi.org/10.3390/molecules27031140
Chicago/Turabian StyleKwiecień, Inga, Natalizia Miceli, Manuela D’Arrigo, Andreana Marino, and Halina Ekiert. 2022. "Antioxidant Potential and Enhancement of Bioactive Metabolite Production in In Vitro Cultures of Scutellaria lateriflora L. by Biotechnological Methods" Molecules 27, no. 3: 1140. https://doi.org/10.3390/molecules27031140
APA StyleKwiecień, I., Miceli, N., D’Arrigo, M., Marino, A., & Ekiert, H. (2022). Antioxidant Potential and Enhancement of Bioactive Metabolite Production in In Vitro Cultures of Scutellaria lateriflora L. by Biotechnological Methods. Molecules, 27(3), 1140. https://doi.org/10.3390/molecules27031140