
Chemistry, Biosynthesis and Pharmacology of Sarsasapogenin: A Potential Natural Steroid Molecule for New Drug Design, Development and Therapy


 , , , ,
, , , ,  ,
,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Phytochemistry of Sarsasapogenin
3. Isolation of Sarsasapogenin
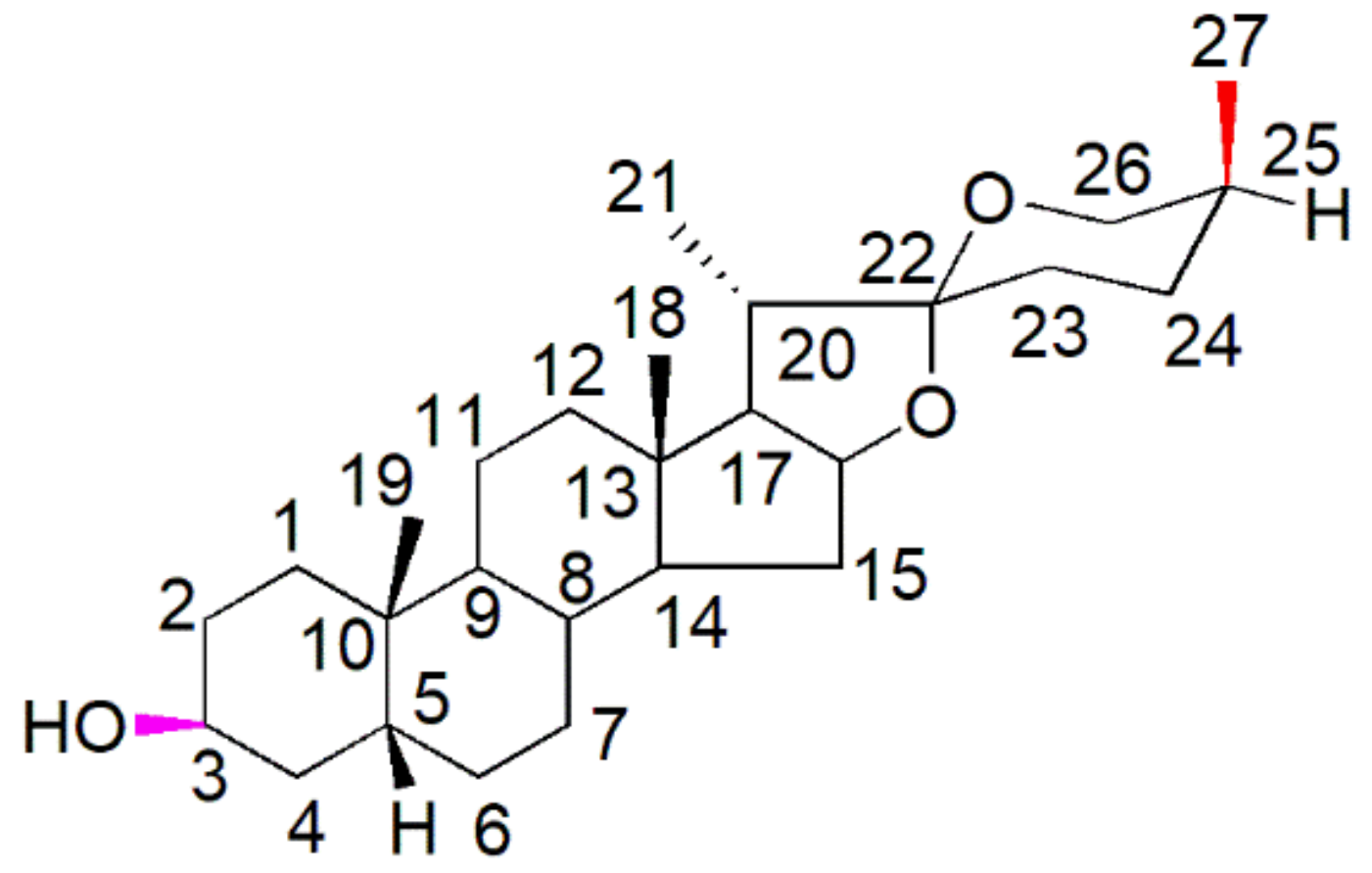
4. Chemistry of Sarsasapogenin
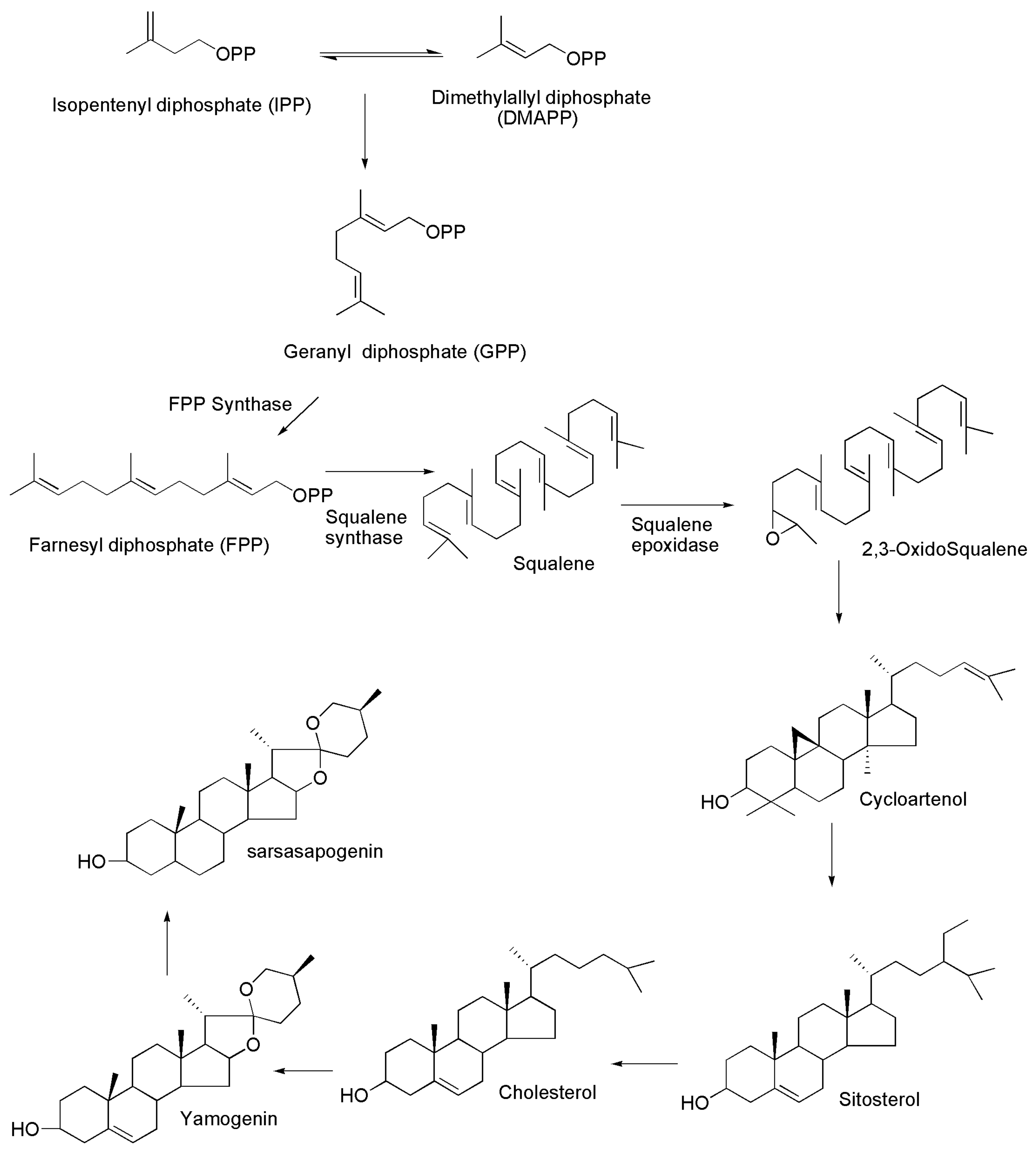
5. Biosynthesis of Sarsasapogenin
6. Physicochemical and Drug-Likeness Properties of Sarsasapogenin
7. Biological Properties of Sarsasapogenin
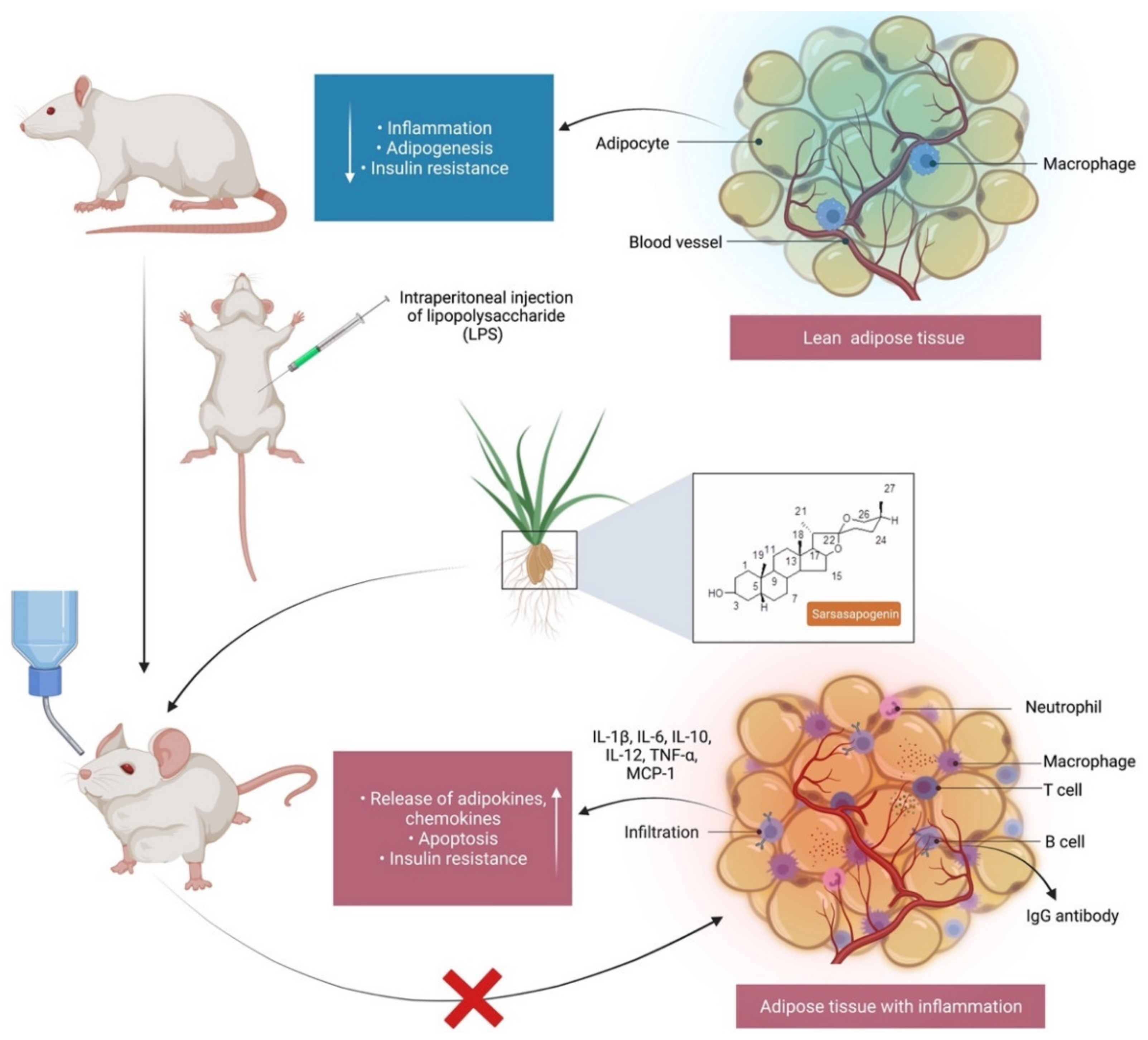
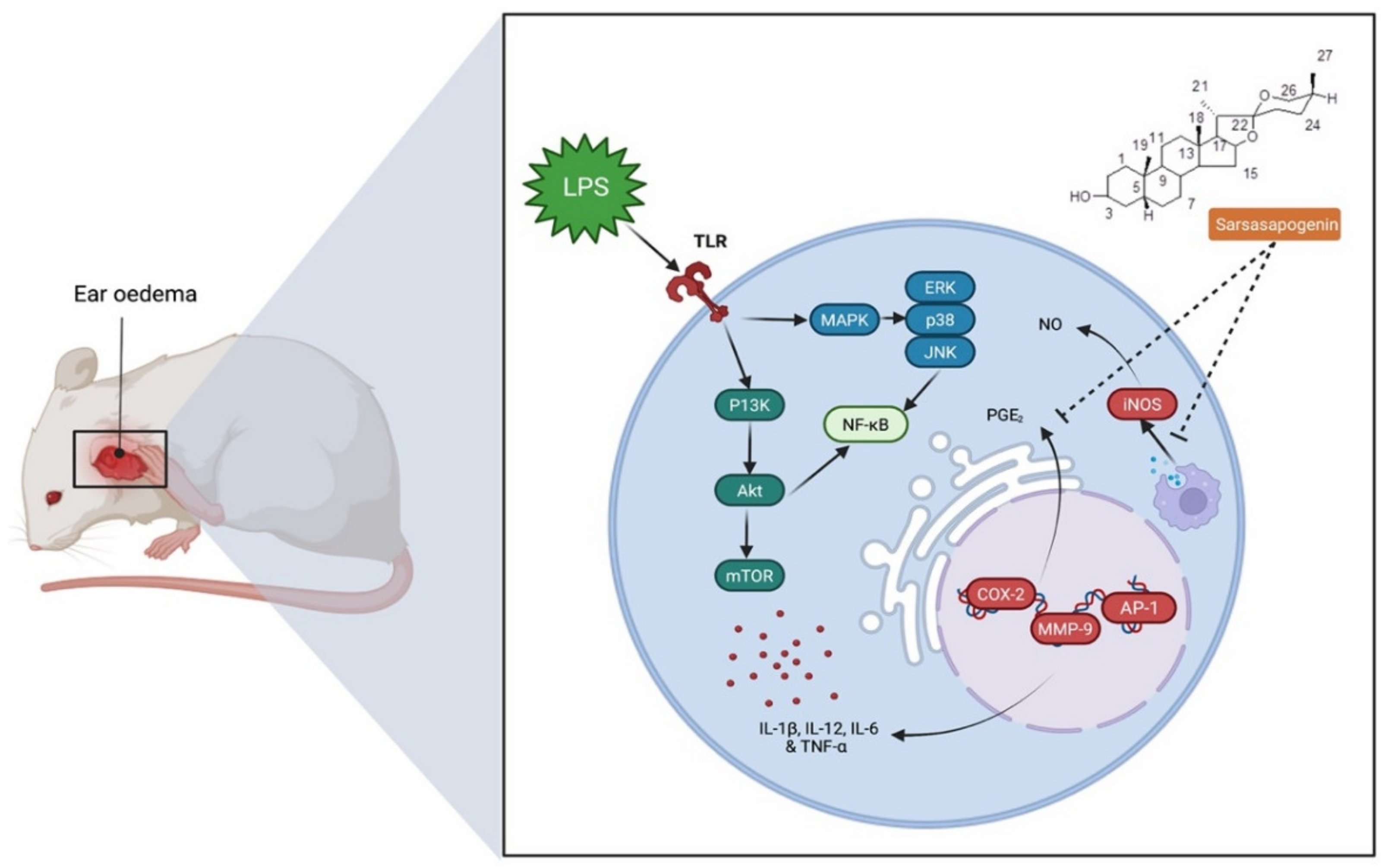
7.1. Anti-Inflammatory Activity
7.2. Anticancer Activity
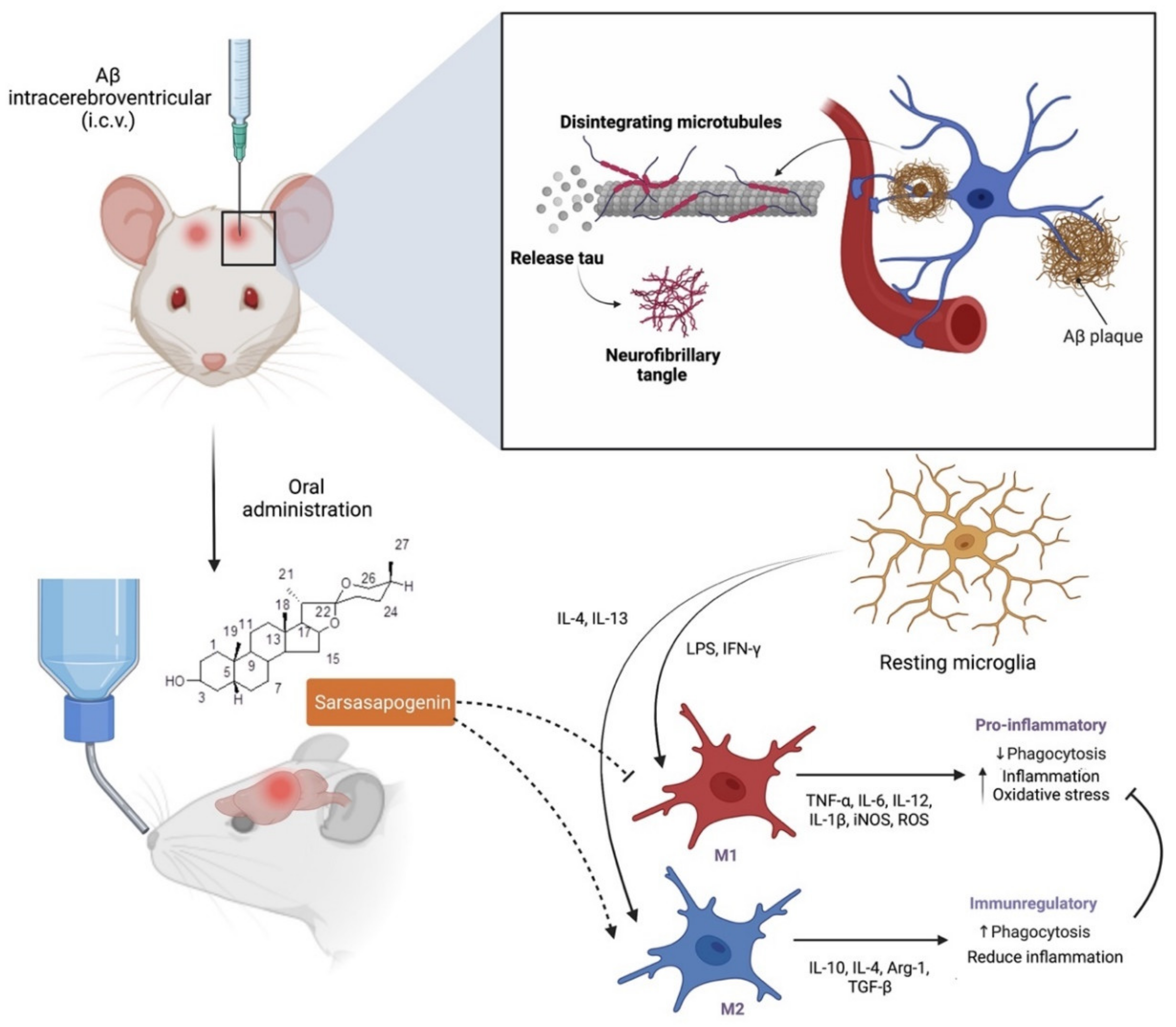
7.3. Neuroprotection
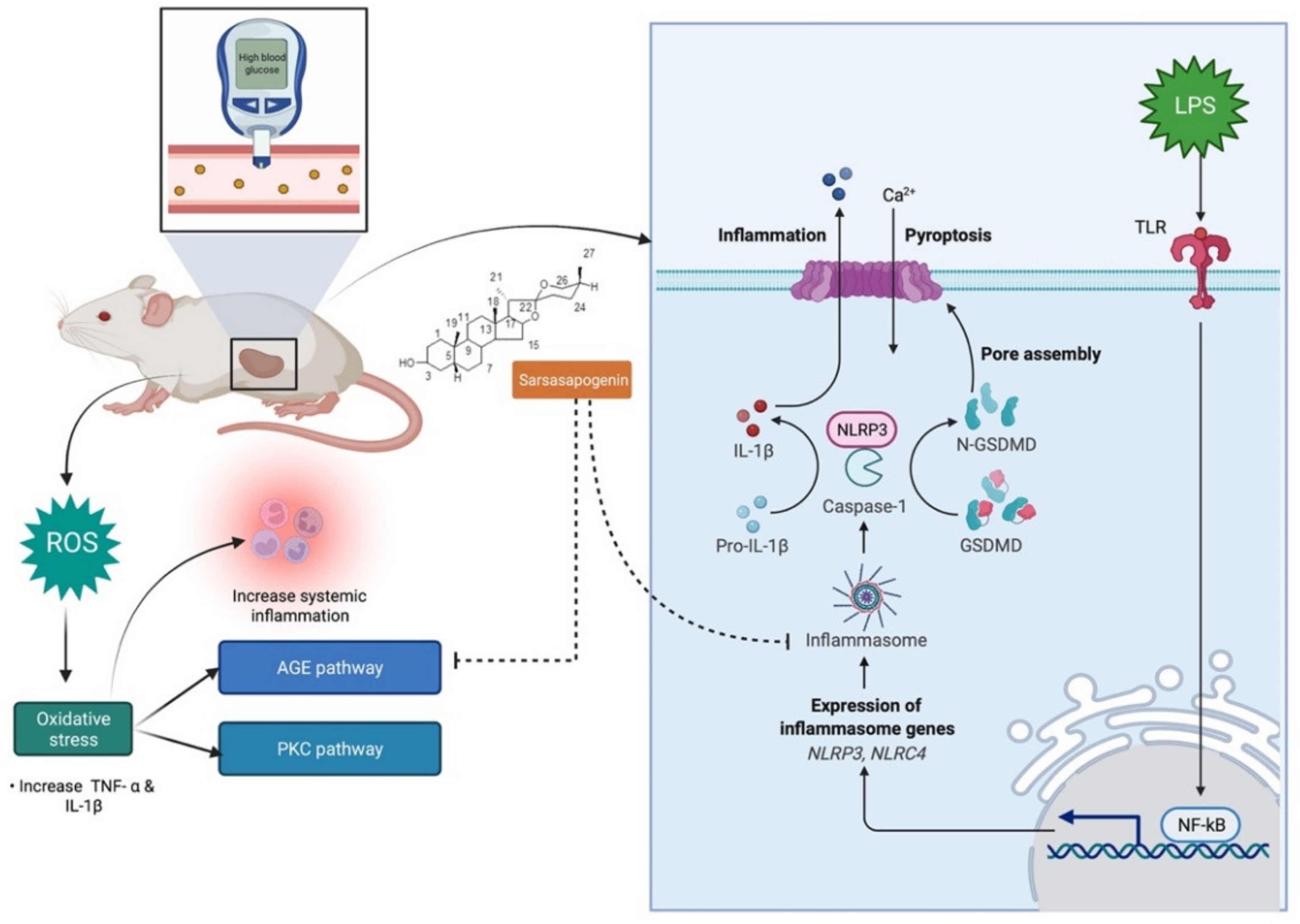
7.4. Antidiabetic Activity
7.5. Anti-Osteoclastogenic Activity
7.6. Treatment for Precocious Puberty
8. Bioavailability and Pharmacokinetics of Sarsasapogenin
9. Metabolism of Sarsasapogenin
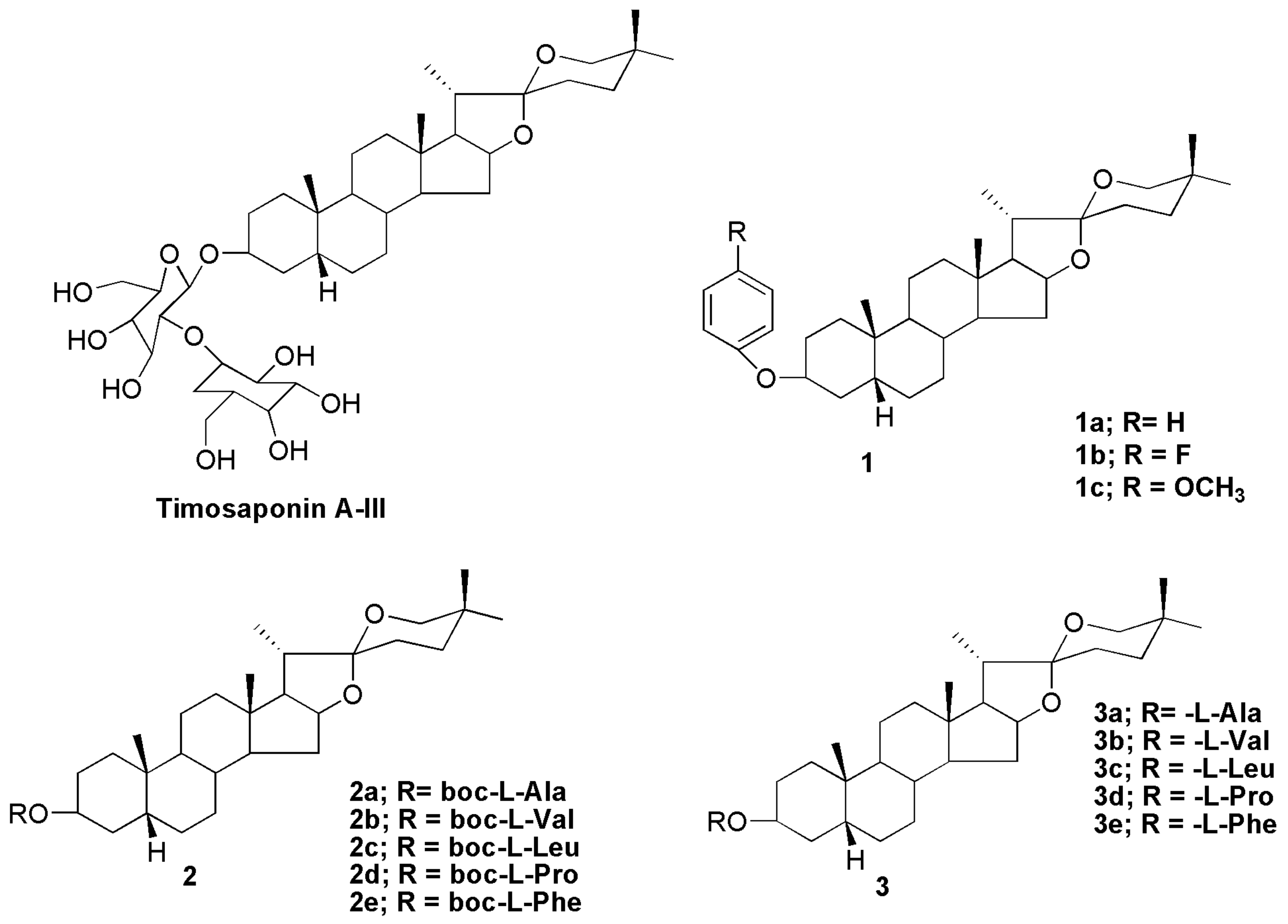
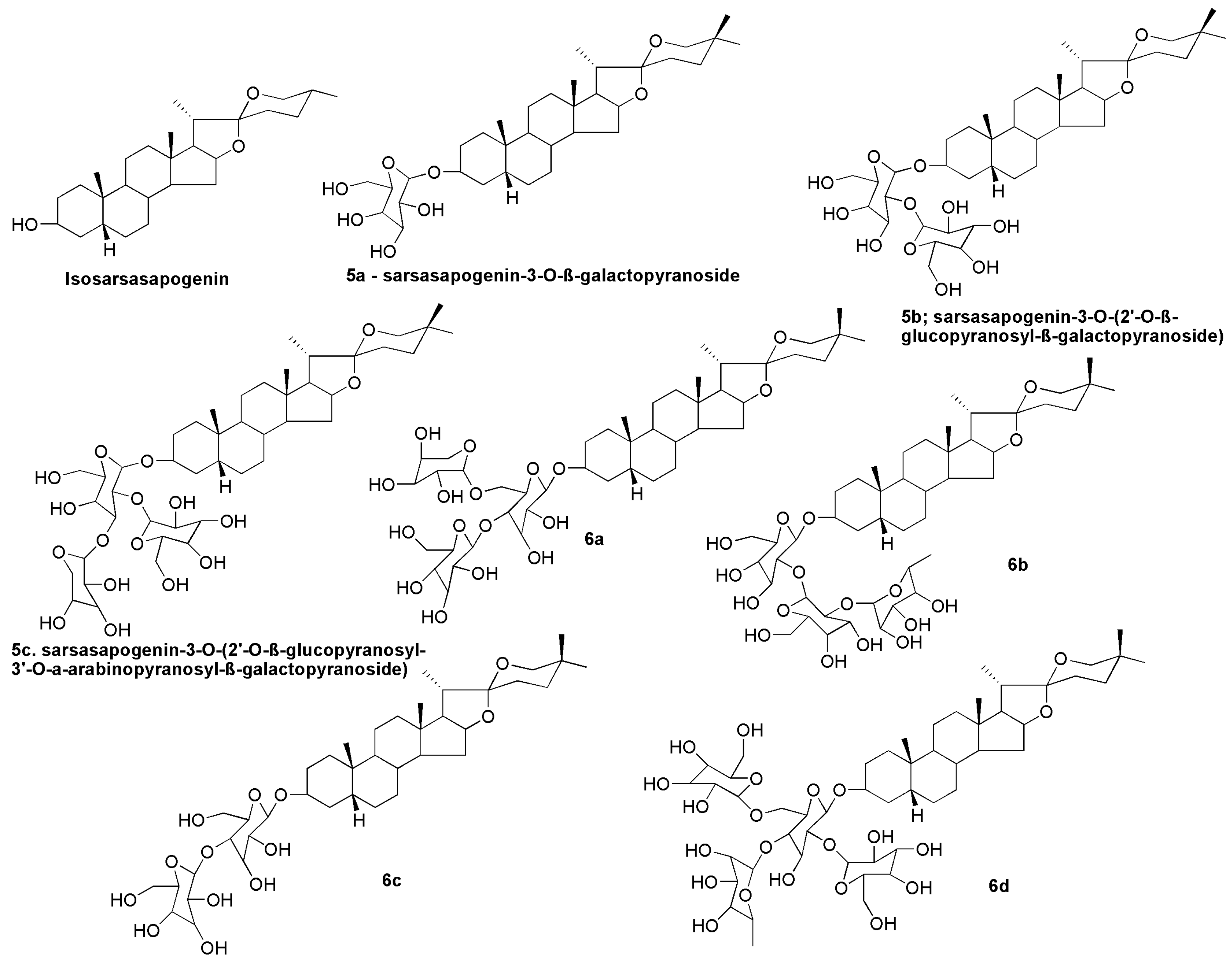
10. Sarsasapogenin-Derived Compounds
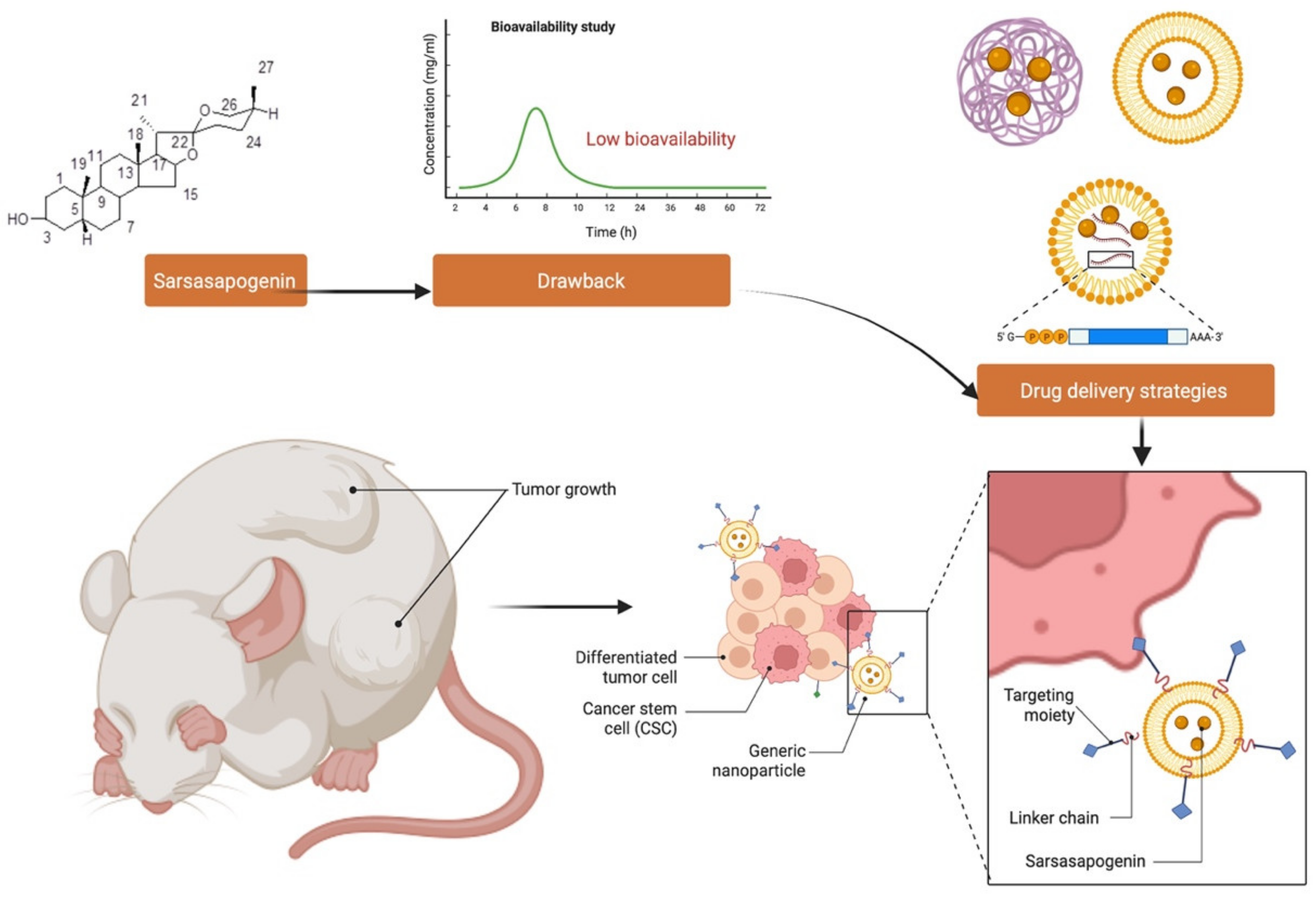
11. Challenges and Opportunities in Developing Sarsasapogenin as a Novel Drug Molecule
12. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hassan, R.; Abdul, B. A Medicinal Plants (Importance and Uses). Pharm. Anal. Acta 2012, 3, 2153–2435. [Google Scholar]
- Li, X.; Jiang, H.; Xu, L.; Liu, Y.-Q.; Tang, J.-w.; Shi, J.-S.; Yu, X.-j.; Wang, X.; Du, L.; Lu, Q. Sarsasapogenin restores podocyte autophagy in diabetic nephropathy by targeting GSK3β signaling pathway. Biochem. Pharmacol. 2021, 192, 114675. [Google Scholar] [CrossRef]
- Othman, S.N.N.; Lum, P.T.; Sekar, M.; Mazlan, N.A.; Yusri, P.Z.S.; Ghazali, N.F.; Idi, H.M.; Azman, S.; Ismail, M.; Noor, A.A.M. Molecules of interest–embelin–a review. Res. J. Pharm. Technol. 2020, 13, 3485–3493. [Google Scholar] [CrossRef]
- Watroly, M.N.; Sekar, M.; Fuloria, S.; Gan, S.H.; Jeyabalan, S.; Wu, Y.S.; Subramaniyan, V.; Sathasivam, K.V.; Ravi, S.; Rani, N.N.I.M. Chemistry, biosynthesis, physicochemical and biological properties of rubiadin: A promising natural anthraquinone for new drug discovery and development. Drug Des. Dev. Ther. 2021, 15, 4527–4549. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.P.; Ahmad, F.; Chatterjee, D.; Bajpai, R.; Sengar, N. Natural products: Drug discovery and development. Drug Discov. Dev. Targets Mol. Med. 2021, 11–65. [Google Scholar] [CrossRef]
- Jeong, J.-J.; Jang, S.-E.; Hyam, S.R.; Han, M.J.; Kim, D.-H. The rhizome mixture of Anemarrhena asphodeloides and Coptidis chinensis ameliorates acute and chronic colitis in mice by inhibiting the binding of lipopolysaccharide to TLR4 and IRAK1 phosphorylation. Evid.-Based Complementary Altern. Med. 2014, 2014, 809083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jigden, B.; Wang, H.; Samdan, N.; Yang, D.-C. Molecular identification of oriental medicinal plant Anemarrhena asphodeloides Bunge (‘Jimo’) by multiplex PCR. Mol. Biol. Rep. 2010, 37, 955. [Google Scholar] [CrossRef]
- Nakashima, N.; Kimura, I.; Kimura, M.; Matsuura, H. Isolation of pseudoprototimosaponin AIII from rhizomes of Anemarrhena asphodeloides and its hypoglycemic activity in streptozotocin-induced diabetic mice. J. Nat. Prod. 1993, 56, 345–350. [Google Scholar] [CrossRef]
- Wang, S.; Su, R.; Nie, S.; Sun, M.; Zhang, J.; Wu, D.; Moustaid-Moussa, N. Application of nanotechnology in improving bioavailability and bioactivity of diet-derived phytochemicals. J. Nutr. Biochem. 2014, 25, 363–376. [Google Scholar] [CrossRef] [Green Version]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; Chinese Medical Science and Technology Press: Beijing, China, 2010; Volume 1, p. 292. [Google Scholar]
- Wang, J.; Liu, J.; Guo, X.; Zhang, J.; Yu, B. A sensitive indirect competitive enzyme-linked immunosorbent assay for the detection of sarsasapogenin in rat plasma. Acta Pharmacol. Sin. 2010, 31, 984–989. [Google Scholar] [CrossRef]
- Dong, D.; Zhou, N.; Liu, R.; Xiong, J.; Pan, H.; Sun, S.; Ma, L.; Wang, R. Sarsasapogenin-AA13 inhibits LPS-induced inflammatory responses in macrophage cells in vitro and relieves dimethylbenzene-induced ear edema in mice. Acta Pharmacol. Sin. 2017, 38, 699–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Dong, D.; Jiao, Q.; Pan, H.; Ma, L.; Wang, R. Sarsasapogenin-AA 13 ameliorates Aβ-induced cognitive deficits via improving neuroglial capacity on Aβ clearance and antiinflammation. CNS Neurosci. Ther. 2017, 23, 498–509. [Google Scholar] [CrossRef] [Green Version]
- Feng, B.; Zhao, X.-Y.; Song, Y.-Z.; Liang, W.-N.; Liu, J.-L. Sarsasapogenin reverses depressive-like behaviors and nicotinic acetylcholine receptors induced by olfactory bulbectomy. Neurosci. Lett. 2017, 639, 173–178. [Google Scholar] [CrossRef]
- Ma, D.; Zhang, J.; Sugahara, K.; Sagara, Y.; Kodama, H. Effect of sarsasapogenin and its derivatives on the stimulus coupled responses of human neutrophils. Clin. Chim. Acta 2001, 314, 107–112. [Google Scholar] [CrossRef]
- Wang, W.; Wang, D.; Wang, Z.; Yao, G.; Li, X.; Gao, P.; Li, L.; Zhang, Y.; Wang, S.; Song, S. Synthesis of new sarsasapogenin derivatives with cytotoxicity and apoptosis-inducing activities in human breast cancer MCF-7 cells. Eur. J. Med. Chem. 2017, 127, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Zhao, X.-C.; Wang, S.-J.; Gao, P.-Y.; Li, L.-Z.; Ikejima, T.; Song, S.-J. Synthesis and biological evaluation of novel sarsasapogenin derivatives as potential anti-tumor agents. Steroids 2015, 93, 25–31. [Google Scholar] [CrossRef]
- Gu, G.; Fang, M.; Liu, J.; Gu, L. Concise synthesis and antitumor activities of trisaccharide steroidal saponins. Carbohydr. Res. 2011, 346, 2406–2413. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Zhang, B.; Fu, Z.; Ma, Y.; Gao, Y.; Shen, F.; Huang, C.; Li, Y. The rapid antidepressant and anxiolytic-like effects of YY-21 involve enhancement of excitatory synaptic transmission via activation of mTOR signaling in the mPFC. Eur. Neuropsychopharmacol. 2016, 26, 1087–1098. [Google Scholar] [CrossRef]
- Sy, L.-K.; Lok, C.-N.; Wang, J.-Y.; Liu, Y.; Cheng, L.; Wan, P.-K.; Leung, C.-T.; Cao, B.; Kwong, W.-L.; Chang, R.C.-C. Identification of “sarsasapogenin-aglyconed” timosaponins as novel Aβ-lowering modulators of amyloid precursor protein processing. Chem. Sci. 2016, 7, 3206–3214. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Wang, R.; Zhou, N.; Xu, T.; Zhao, J.; Zhang, Y.; Pan, H. Sarsasapogenin derivative, its preparation method and application. Invention Patent CN 104513289, 15 April 2015. [Google Scholar]
- Moon, E.; Kim, A.-J.; Kim, S.Y. Sarsasapogenin increases melanin synthesis via induction of tyrosinase and microphthalmia-associated transcription factor expression in melan-a cells. Biomol. Ther. 2012, 20, 340. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Zhao, K.; Zhu, J.; Wang, Y.; Sun, P.; Yang, Q.; Zhang, T.; Han, W.; Hu, W.; Yang, W. Sarsasapogenin suppresses RANKL-induced osteoclastogenesis in vitro and prevents lipopolysaccharide-induced bone loss in vivo. Drug Des. Dev. Ther. 2020, 14, 3435. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Van Khang, P.; Dong, D.; Wang, R.; Ma, L. Synthesis and SAR study of novel sarsasapogenin derivatives as potent neuroprotective agents and NO production inhibitors. Bioorganic Med. Chem. Lett. 2017, 27, 662–665. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.-X.; Luo, Y.-F.; Li, X.; Zuo, D.-Y.; Wu, Y.-L. Antidepressant-Like Effects of sarsasapogenin from Anemarrhena asphodeloides B UNGE (Liliaceae). Biol. Pharm. Bull. 2006, 29, 2304–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.W.; Hao, Y.C.; Chen, Y.J.; Yin, S.Y.; Zhang, M.Y.; Kong, L.; Wang, T.Y. Protective effects of sarsasapogenin against early stage of diabetic nephropathy in rats. Phytother. Res. 2018, 32, 1574–1582. [Google Scholar] [CrossRef]
- Hu, K.; Sun, W.; Li, Y.; Zhang, B.; Zhang, M.; Guo, C.; Chang, H.; Wang, X. Study on the mechanism of sarsasapogenin in treating precocious puberty by regulating the HPG Axis. Evid. Based Complementary Altern. Med. 2020, 2020, 1978043. [Google Scholar] [CrossRef]
- Lim, S.-M.; Jeong, J.-J.; Kang, G.-D.; Kim, K.-A.; Choi, H.-S.; Kim, D.-H. Timosaponin AIII and its metabolite sarsasapogenin ameliorate colitis in mice by inhibiting NF-κB and MAPK activation and restoring Th17/Treg cell balance. Int. Immunopharmacol. 2015, 25, 493–503. [Google Scholar] [CrossRef]
- Bao, W.; Pan, H.; Lu, M.; Ni, Y.; Zhang, R.; Gong, X. The apoptotic effect of sarsasapogenin from Anemarrhena asphodeloides on HepG2 human hepatoma cells. Cell Biol. Int. 2007, 31, 887–892. [Google Scholar] [CrossRef]
- Tang, Z.-Z.; Zhang, Y.-M.; Zheng, T.; Huang, T.-T.; Ma, T.-F.; Liu, Y.-W. Sarsasapogenin alleviates diabetic nephropathy through suppression of chronic inflammation by down-regulating PAR-1: In vivo and in vitro study. Phytomedicine 2020, 78, 153314. [Google Scholar] [CrossRef]
- Pei, L.; Ye, Y.; Zhao, W.; Ye, Q.; Ge, S.; Jiang, Z.; Liang, X.; Gan, H.; Ma, L. A validated UPLC–MS/MS method for quantitative determination of a potent neuroprotective agent Sarsasapogenin-AA13 in rat plasma: Application to pharmacokinetic studies. Biomed. Chromatogr. 2020, 34, e4775. [Google Scholar] [CrossRef]
- Kashyap, P.; Muthusamy, K.; Niranjan, M.; Trikha, S.; Kumar, S. Sarsasapogenin: A steroidal saponin from Asparagus racemosus as multi target directed ligand in Alzheimer’s disease. Steroids 2020, 153, 108529. [Google Scholar] [CrossRef]
- Paczkowski, C.; Wojciechowski, Z.A. The occurrence of UDPG-dependent glucosyltransferase specific for sarsasapogenin in Asparagus officinalis. Phytochemistry 1988, 27, 2743–2747. [Google Scholar] [CrossRef]
- Huang, X.F.; Lin, Y.Y.; Kong, L.Y. Steroids from the roots of Asparagus officinalis and their cytotoxic activity. J. Integr. Plant Biol. 2008, 50, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Okanishi, T.; Akahori, A.; Yasuda, F.; Takeuchi, Y.; Iwao, T. Steroidal sapogenins of sixteen Liliaceae plants. Chem. Pharm. Bull. 1975, 23, 575–579. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Lu, Y.; Qian, W.; Pei, Z. Asparagus cochinchinensis (Lour.) Merr. (Tiandong, Chinese Asparagus). In Dietary Chinese Herbs; Springer: Berlin/Heidelberg, Germany, 2015; pp. 83–88. [Google Scholar]
- Flåøyen, A.; Wilkins, A.; Sandvik, M. Ruminal metabolism in sheep of saponins from Yucca schidigera. Vet. Res. Commun. 2002, 26, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Ingawale, D.K.; Mandlik, S.K.; Patel, S.S. Combination of sarsasapogenin and fluticasone attenuates ovalbumin-induced airway inflammation in a mouse asthma model. Immunopharmacol. Immunotoxicol. 2020, 42, 128–137. [Google Scholar] [CrossRef]
- Power, F.B.; Salway, A.H. XXV.—Chemical examination of sarsaparilla root. J. Chem. Soc. Trans. 1914, 105, 201–219. [Google Scholar] [CrossRef]
- Mandlik, D.S.; Mandlik, S.K.; Patel, S. Protective effect of sarsasapogenin in TNBS induced ulcerative colitis in rats associated with downregulation of pro-inflammatory mediators and oxidative stress. Immunopharmacol. Immunotoxicol. 2021, 43, 571–583. [Google Scholar] [CrossRef]
- Marker, R.E.; Wagner, R.; Ulshafer, P.R.; Wittbecker, E.L.; Goldsmith, D.P.; Ruof, C.H. Steroidal sapogenins1a. J. Am. Chem. Soc. 1947, 69, 2167–2230. [Google Scholar] [CrossRef]
- Uhlig, S.; Wisløff, H.; Petersen, D. Identification of cytotoxic constituents of Narthecium ossifragum using bioassay-guided fractionation. J. Agric. Food Chem. 2007, 55, 6018–6026. [Google Scholar] [CrossRef]
- Ceh, L.; Hauge, J.G. Alveld-producing saponins. I. Chemical studies. Acta Vet. Scand. 1981, 22, 391–402. [Google Scholar] [CrossRef]
- Duckstein, S.M.; Stintzing, F.C. LC–MSn characterization of steroidal saponins in Helleborus niger L. roots and their conversion products during fermentation. Steroids 2015, 93, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Li, Z.; Xue, R.; Jiang, J.; Huang, C. Stereoisomerism metabolites found in rats after oral administration of timosaponin B-II using HPLC-Q-TOF-MS and NMR methods. RSC Adv. 2015, 5, 60650–60657. [Google Scholar] [CrossRef]
- Jadhav, A.; Bhutani, K. Steroidal saponins from the roots of Asparagus adscendens Roxb and Asparagus racemosus Willd. Indian J. Chem. 2006, 45B, 1515–1524. [Google Scholar]
- Srivastava, P.L.; Daramwar, P.P.; Krithika, R.; Pandreka, A.; Shankar, S.S.; Thulasiram, H.V. Functional characterization of novel sesquiterpene synthases from Indian sandalwood, Santalum album. Sci. Rep. 2015, 5, 10095. [Google Scholar] [CrossRef] [Green Version]
- Eisenreich, W.; Rohdich, F.; Bacher, A. Deoxyxylulose phosphate pathway to terpenoids. Trends Plant Sci. 2001, 6, 78–84. [Google Scholar] [CrossRef]
- Vickers, C.E.; Gershenzon, J.; Lerdau, M.T.; Loreto, F. A unified mechanism of action for volatile isoprenoids in plant abiotic stress. Nat. Chem. Biol. 2009, 5, 283–291. [Google Scholar] [CrossRef]
- Lange, B.M.; Rujan, T.; Martin, W.; Croteau, R. Isoprenoid biosynthesis: The evolution of two ancient and distinct pathways across genomes. Proc. Natl. Acad. Sci. USA 2000, 97, 13172–13177. [Google Scholar] [CrossRef] [Green Version]
- Bridges, N.; Christopher, J.; Hindmarsh, P.; Brook, C. Sexual precocity: Sex incidence and aetiology. Arch. Dis. Child. 1994, 70, 116–118. [Google Scholar] [CrossRef] [Green Version]
- Cantas-Orsdemir, S.; Eugster, E.A. Update on central precocious puberty: From etiologies to outcomes. Expert Rev. Endocrinol. Metab. 2019, 14, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Zhou, W.; Wei, L.; Li, J.; Jia, J.; Li, F.; Smith, S.M.; Xu, J. Regulation of the cholesterol biosynthetic pathway and its integration with fatty acid biosynthesis in the oleaginous microalga Nannochloropsis oceanica. Biotechnol. Biofuels 2014, 7, 81. [Google Scholar] [CrossRef] [Green Version]
- Gholami, A.; De Geyter, N.; Pollier, J.; Goormachtig, S.; Goossens, A. Natural product biosynthesis in Medicago species. Nat. Prod. Rep. 2014, 31, 356–380. [Google Scholar] [CrossRef] [PubMed]
- Haralampidis, K.; Trojanowska, M.; Osbourn, A.E. Biosynthesis of triterpenoid saponins in plants. Adv. Biochem. Eng. Biotechnol. 2002, 75, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Kemp, M.W.; Saito, M.; Newnham, J.P.; Nitsos, I.; Okamura, K.; Kallapur, S.G. Preterm birth, infection, and inflammation advances from the study of animal models. Reprod. Sci. 2010, 17, 619–628. [Google Scholar] [CrossRef] [PubMed]
- James, M.W.; Hawkey, C.J. Assessment of non-steroidal anti-inflammatory drug (NSAID) damage in the human gastrointestinal tract. Br. J. Clin. Pharmacol. 2003, 56, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.Y.; Cui, S.C.; Zheng, T.N.; Ma, H.J.; Xie, Z.F.; Jiang, H.W.; Li, Y.F.; Zhu, K.X.; Huang, C.G.; Li, J.; et al. Sarsasapogenin improves adipose tissue inflammation and ameliorates insulin resistance in high-fat diet-fed C57BL/6J mice. Acta Pharmacol. Sin. 2021, 42, 272–281. [Google Scholar] [CrossRef]
- Ni, Y.; Gong, X.G.; Lu, M.; Chen, H.M.; Wang, Y. Mitochondrial ROS burst as an early sign in sarsasapogenin-induced apoptosis in HepG2 cells. Cell Biol. Int. 2008, 32, 337–343. [Google Scholar] [CrossRef]
- Bai, J.; Cederbaum, A.I. Overexpression of CYP2E1 in mitochondria sensitizes HepG2 cells to the toxicity caused by depletion of glutathione. J. Biol. Chem. 2006, 281, 5128–5136. [Google Scholar] [CrossRef] [Green Version]
- Ling, Y.-H.; Liebes, L.; Zou, Y.; Perez-Soler, R. Reactive oxygen species generation and mitochondrial dysfunction in the apoptotic response to Bortezomib, a novel proteasome inhibitor, in human H460 non-small cell lung cancer cells. J. Biol. Chem. 2003, 278, 33714–33723. [Google Scholar] [CrossRef] [Green Version]
- Chandra, D.; Liu, J.-W.; Tang, D.G. Early Mitochondrial activation and cytochrome c up-regulation during apoptosis*210. J. Biol. Chem. 2002, 277, 50842–50854. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Zhang, Y.; Zhang, R.; Gong, X. Sarsasapogenin induces apoptosis via the reactive oxygen species-mediated mitochondrial pathway and ER stress pathway in HeLa cells. Biochem. Biophys. Res. Commun. 2013, 441, 519–524. [Google Scholar] [CrossRef]
- Zhang, Y.-M.; Zheng, T.; Huang, T.-T.; Gu, P.-P.; Gou, L.-s.; Ma, T.-F.; Liu, Y.-W. Sarsasapogenin attenuates Alzheimer-like encephalopathy in diabetes. Phytomedicine 2021, 91, 153686. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xia, Z.; Sun, Q.; Orsi, A.; Rees, D. A new approach to the pharmacological regulation of memory: Sarsasapogenin improves memory by elevating the low muscarinic acetylcholine receptor density in brains of memory-deficit rat models. Brain Res. 2005, 1060, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.; Moriguchi, S.; Tagashira, H.; Fukunaga, K. Rivastigmine improves hippocampal neurogenesis and depression-like behaviors via 5-HT1A receptor stimulation in olfactory bulbectomized mice. Neuroscience 2014, 272, 116–130. [Google Scholar] [CrossRef]
- Andreasen, J.T.; Nielsen, E.Ø.; Redrobe, J.P. Chronic oral nicotine increases brain [3H] epibatidine binding and responsiveness to antidepressant drugs, but not nicotine, in the mouse forced swim test. Psychopharmacology 2009, 205, 517–528. [Google Scholar] [CrossRef]
- Lendvai, B.; Kassai, F.; Szájli, Á.; Némethy, Z. α7 nicotinic acetylcholine receptors and their role in cognition. Brain Res. Bull. 2013, 93, 86–96. [Google Scholar] [CrossRef]
- Séguéla, P.; Wadiche, J.; Dineley-Miller, K.; Dani, J.A.; Patrick, J.W. Molecular cloning, functional properties, and distribution of rat brain alpha 7: A nicotinic cation channel highly permeable to calcium. J. Neurosci. 1993, 13, 596–604. [Google Scholar] [CrossRef] [Green Version]
- Wada, E.; Wada, K.; Boulter, J.; Deneris, E.; Heinemann, S.; Patrick, J.; Swanson, L.W. Distribution of alpha2, alpha3, alpha4, and beta2 neuronal nicotinic receptor subunit mRNAs in the central nervous system: A hybridization histochemical study in the rat. J. Comp. Neurol. 1989, 284, 314–335. [Google Scholar] [CrossRef]
- Zhang, M.-Y.; Li, Y.; Yin, S.-Y.; Kong, L.; Liu, X.-L.; Yin, X.-X.; Liu, Y.-W. Sarsasapogenin suppresses Aβ overproduction induced by high glucose in HT-22 cells. Naunyn Schmiedeberg’s Arch. Pharmacol. 2018, 391, 159–168. [Google Scholar] [CrossRef]
- Shlipak, M. Diabetic nephropathy: Preventing progression. BMJ Clin. Evid. 2010, 7, 606. [Google Scholar]
- Qiu, Y.Y.; Tang, L.Q. Roles of the NLRP3 inflammasome in the pathogenesis of diabetic nephropathy. Pharmacol. Res. 2016, 114, 251–264. [Google Scholar] [CrossRef]
- Zheng, N.; Shi, X.; Chen, X.; Lv, W. Associations between inflammatory markers, hemostatic markers, and microvascular complications in 182 Chinese patients with type 2 diabetes mellitus. Lab. Med. 2015, 46, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tang, Z.-Z.; Zhang, Y.-M.; Kong, L.; Xiao, W.-F.; Ma, T.-F.; Liu, Y.-W. Thrombin/PAR-1 activation induces endothelial damages via NLRP1 inflammasome in gestational diabetes. Biochem. Pharmacol. 2020, 175, 113849. [Google Scholar] [CrossRef]
- Kong, L.; Liu, Y.; Zhang, Y.M.; Li, Y.; Gou, L.S.; Ma, T.F.; Liu, Y.W. Sarsasapogenin ameliorates diabetes-associated memory impairment and neuroinflammation through down-regulation of PAR-1 receptor. Phytother. Res. 2021, 35, 3167–3180. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.; Horowitz, M.; Choi, Y. Osteoimmunology: Interactions of the bone and immune system. Endocr. Rev. 2008, 29, 403–440. [Google Scholar] [CrossRef]
- Arai, F.; Miyamoto, T.; Ohneda, O.; Inada, T.; Sudo, T.; Brasel, K.; Miyata, T.; Anderson, D.M.; Suda, T. Commitment and differentiation of osteoclast precursor cells by the sequential expression of c-Fms and receptor activator of nuclear factor κB (RANK) receptors. J. Exp. Med. 1999, 190, 1741–1754. [Google Scholar] [CrossRef]
- Takayanagi, H. Osteoimmunology: Shared mechanisms and crosstalk between the immune and bone systems. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Darnay, B.G.; Besse, A.; Poblenz, A.T.; Lamothe, B.; Jacoby, J.J. TRAFs in RANK signaling. Adv. Exp. Med. Biol. 2007, 597, 152–159. [Google Scholar] [PubMed]
- Mizukami, J.; Takaesu, G.; Akatsuka, H.; Sakurai, H.; Ninomiya-Tsuji, J.; Matsumoto, K.; Sakurai, N. Receptor activator of NF-κB ligand (RANKL) activates TAK1 mitogen-activated protein kinase kinase kinase through a signaling complex containing RANK, TAB2, and TRAF6. Mol. Cell. Biol. 2002, 22, 992–1000. [Google Scholar] [CrossRef] [Green Version]
- Wong, B.R.; Besser, D.; Kim, N.; Arron, J.R.; Vologodskaia, M.; Hanafusa, H.; Choi, Y. TRANCE, a TNF family member, activates Akt/PKB through a signaling complex involving TRAF6 and c-Src. Mol. Cell 1999, 4, 1041–1049. [Google Scholar] [CrossRef]
- Kim, M.S.; Yang, Y.-M.; Son, A.; Tian, Y.S.; Lee, S.-I.; Kang, S.W.; Muallem, S.; Shin, D.M. RANKL-mediated reactive oxygen species pathway that induces long lasting Ca2+ oscillations essential for osteoclastogenesis. J. Biol. Chem. 2010, 285, 6913–6921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Kim, N. Regulation of NFATc1 in osteoclast differentiation. J. Bone Metab. 2014, 21, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano-Guillen, L.; Argente, J. Central precocious puberty, functional and tumor-related. Best Pract. Res. Clin. Endocrinol. Metab. 2019, 33, 101262. [Google Scholar] [CrossRef] [PubMed]
- Guaraldi, F.; Beccuti, G.; Gori, D.; Ghizzoni, L. Management of endocrine disease: Long-term outcomes of the treatment of central precocious puberty. Eur. J. Endocrinol. 2016, 174, R79–R87. [Google Scholar] [CrossRef]
- Rostami, S.; Kohan, L.; Mohammadianpanah, M. The LEP G-2548A gene polymorphism is associated with age at menarche and breast cancer susceptibility. Gene 2015, 557, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Day, F.R.; Thompson, D.J.; Helgason, H.; Chasman, D.I.; Finucane, H.; Sulem, P.; Ruth, K.S.; Whalen, S.; Sarkar, A.K.; Albrecht, E. Genomic analyses identify hundreds of variants associated with age at menarche and support a role for puberty timing in cancer risk. Nat. Genet. 2017, 49, 834–841. [Google Scholar] [CrossRef]
- Chen, M.; Eugster, E.A. Central precocious puberty: Update on diagnosis and treatment. Pediatric Drugs 2015, 17, 273–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendirci, H.N.P.; Ağladıoğlu, S.Y.; Önder, A.; Baş, V.N.; Çetinkaya, S.; Aycan, Z. Effects of GnRH analogue treatment on anterior pituitary hormones in children with central precocious puberty. J. Pediatric Endocrinol. Metab. 2015, 28, 1145–1151. [Google Scholar]
- Yen, S. Clinical applications of gonadotropin-releasing hormone and gonadotropin-releasing hormone analogs. Fertil. Steril. 1983, 39, 257–266. [Google Scholar]
- Li, W.-J.; Gong, C.-X.; Guo, M.-J.; Xing, J.; Li, T.; Song, W.-H.; Luo, X.-P.; Wu, D.; Liang, J.-P.; Cao, B.-Y. Efficacy and safety of domestic leuprorelin in girls with idiopathic central precocious puberty: A multicenter, randomized, parallel, controlled trial. Chin. Med. J. 2015, 128, 1314. [Google Scholar] [CrossRef]
- Latronico, A.C.; Brito, V.N.; Carel, J.-C. Causes, diagnosis, and treatment of central precocious puberty. Lancet Diabetes Endocrinol. 2016, 4, 265–274. [Google Scholar] [CrossRef]
- Carel, J.-C.; Eugster, E.A.; Rogol, A.; Ghizzoni, L.; Palmert, M.R. Consensus statement on the use of gonadotropin-releasing hormone analogs in children. Pediatrics 2009, 123, e752–e762. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Liu, Z.; Hu, J.; Lai, X.; Xia, P. Quantitative determination of sarsasapogenin in rat plasma using liquid chromatography-tandem mass spectrometry. J. Chromatogr. B 2016, 1022, 213–219. [Google Scholar] [CrossRef]
- Liu, Z.; Qin, W.; Zhu, Z.; Liu, Y.; Sun, F.; Chai, Y.; Xia, P. Development and validation of liquid chromatography-tandem mass spectrometry method for simultaneous determination of six steroidal saponins in rat plasma and its application to a pharmacokinetics study. Steroids 2015, 96, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, Y.; Yao, G.; Wang, W.; Shang, X.; Zhang, Y.; Wang, X.; Wang, S.; Song, S. Synthesis of new sarsasapogenin derivatives with antiproliferative and apoptotic effects in MCF-7 cells. Steroids 2018, 131, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Díaz, A.E.C.; Herfindal, L.; Rathe, B.A.; Sletta, K.Y.; Vedeler, A.; Haavik, S.; Fossen, T. Cytotoxic saponins and other natural products from flowering tops of Narthecium ossifragum L. Phytochemistry 2019, 164, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.-W.; Zhang, Z.; Long, H.-L.; Zhang, Y.-J. Steroidal saponins from the genus Smilax and their biological activities. Nat. Prod. Bioprospect. 2017, 7, 283–298. [Google Scholar] [CrossRef] [Green Version]
- Bonferoni, M.C.; Rossi, S.; Sandri, G.; Ferrari, F. Nanoparticle formulations to enhance tumor targeting of poorly soluble polyphenols with potential anticancer properties. In Proceedings of the Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2017; pp. 205–214. [Google Scholar]
- Lagoa, R.; Silva, J.; Rodrigues, J.R.; Bishayee, A. Advances in phytochemical delivery systems for improved anticancer activity. Biotechnol. Adv. 2020, 38, 107382. [Google Scholar] [CrossRef]
- Pistollato, F.; Bremer-Hoffmann, S.; Basso, G.; Cano, S.S.; Elio, I.; Vergara, M.M.; Giampieri, F.; Battino, M. Targeting glioblastoma with the use of phytocompounds and nanoparticles. Target. Oncol. 2016, 11, 1–16. [Google Scholar] [CrossRef]
- Rahimi, H.R.; Nedaeinia, R.; Shamloo, A.S.; Nikdoust, S.; Oskuee, R.K. Novel delivery system for natural products: Nano-curcumin formulations. Avicenna J. Phytomed. 2016, 6, 383. [Google Scholar]
- Siddiqui, I.A.; Sanna, V. Impact of nanotechnology on the delivery of natural products for cancer prevention and therapy. Mol. Nutr. Food Res. 2016, 60, 1330–1341. [Google Scholar] [CrossRef] [PubMed]
- Zuraini, N.Z.A.; Sekar, M.; Wu, Y.S.; Gan, S.H.; Bonam, S.R.; Rani, N.N.I.M.; Begum, M.Y.; Lum, P.T.; Subramaniyan, V.; Fuloria, N.K. Promising nutritional fruits against cardiovascular diseases: An overview of experimental evidence and understanding their mechanisms of action. Vasc. Health Risk Manag. 2021, 17, 739–769. [Google Scholar] [CrossRef] [PubMed]
- Yap, K.M.; Sekar, M.; Fuloria, S.; Wu, Y.S.; Gan, S.H.; Rani, N.N.I.M.; Subramaniyan, V.; Kokare, C.; Lum, P.T.; Begum, M.Y. Drug delivery of natural products through nanocarriers for effective breast cancer therapy: A comprehensive review of literature. Int. J. Nanomed. 2021, 16, 7891–7941. [Google Scholar] [CrossRef] [PubMed]
- Davatgaran-Taghipour, Y.; Masoomzadeh, S.; Farzaei, M.H.; Bahramsoltani, R.; Karimi-Soureh, Z.; Rahimi, R.; Abdollahi, M. Polyphenol nanoformulations for cancer therapy: Experimental evidence and clinical perspective. Int. J. Nanomed. 2017, 12, 2689–2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Family | Genus | Plant Species | References |
|---|---|---|---|
| Asparagaceae | Anemarrhena | Anemarrhena asphodeloides | Wang et al. [11]; Moon et al. [22]; Peng et al. [23]; Pan et al. [24]; Ren et al. [25]; Liu et al. [26]; Hu et al. [27]; Lim et al. [28]; Bao et al. [29]; Tang et al. [30]; Pei et al. [31] |
| Asparagus | Asparagus racemosus | Kashyap et al. [32] | |
| Asparagus officinalis | Paczkowski and Wojciechowski [33]; Huang et al. [34] | ||
| Asparagus cochinchinensis | Okanishi et al. [35]; Zhang et al. [36] | ||
| Yucca | Yucca schidigera | Flåøyen et al. [37] | |
| Smilacaceae | Smilax | Smilax china | Ingawale et al. [38] |
| Smilax ornata | Ingawale et al. [38]; Power and Salway [39]; Mandlik et al. [40] | ||
| Smilax aspera | Ingawale et al. [38]; Mandlik et al. [40] | ||
| Smilax febrifuga | Ingawale et al. [38]; Mandlik et al. [40] | ||
| Smilax aristolochiifolia | Ingawale et al. [38]; Mandlik et al. [40] | ||
| Smilax officinalis | Ingawale et al. [38] | ||
| Smilax medica | Marker et al. [41] | ||
| Nartheciaceae | Narthecium | Narthecium ossifragum | Uhlig et al. [42]; Ceh and Hauge [43] |
| Ranunculaceae | Helleborus | Helleborus niger | Duckstein and Stintzing [44] |
| Carbon | Signal (δ) | Proton | Signal (δ) (Multiplicity) |
|---|---|---|---|
| 1 | 30.1 | 1 | 1.49 (m), 1.24 (m) |
| 2 | 37.0 | 2 | 1.72 (m), 1.47 (m) |
| 3 | 67.2 | 3 | 4.10 (br. s) |
| 4 | 33.7 | 4 | 1.67 (m), 1.41 (m) |
| 5 | 36.7 | 5 | 1.42 (m) |
| 6 | 26.7 | 6 | 2.1 (m), 2.7 (m) |
| 7 | 26.7 | 7 | 1.52 (m), 1.27 (m) |
| 8 | 35.4 | 8 | 1.56 (m) |
| 9 | 40.0 | 9 | 1.40 (d) |
| 10 | 35.4 | 10 | - |
| 11 | 21.0 | 11 | 1.35 (m) |
| 12 | 40.5 | 12 | 1.14 (m), 1.65 (m) |
| 13 | 40.8 | 13 | - |
| 14 | 56.6 | 14 | 1.18 (m) |
| 15 | 31.9 | 15 | 1.73 (m), 1.46 (m) |
| 16 | 81.2 | 16 | 4.40 (td, J = 7.9, 7.3, 6.2 Hz) |
| 17 | 62.2 | 17 | 1.79 (m) |
| 18 | 16.2 | 18 | 0.75 (s) |
| 19 | 24.1 | 19 | 0.97 (s) |
| 20 | 42.3 | 20 | 1.26 (m) |
| 21 | 14.5 | 21 | 1.07 (d, J = 7.1 Hz) |
| 22 | 109.9 | 22 | - |
| 23 | 28.0 | 23 | 1.95 (m) |
| 24 | 25.9 | 24 | 1.66 (m), 1.44 (m) |
| 25 | 27.2 | 25 | 1.77 (m) |
| 26 | 65.3 | 26 | 3.94 (dd, J = 11.0, 2.8 Hz), 3.29 (d, J = 10.9 Hz) |
| 27 | 16.6 | 27 | 0.98 (d, J = 6.4 Hz) |
| Property/Rule | Result |
|---|---|
| Molecular formula | C27H44O3 |
| Molecular weight | 416.6 g/mol |
| Hydrogen bond donors | 1 |
| Hydrogen bond acceptors | 3 |
| Rotatable bonds | 0 |
| Log P (partition coefficient, predicted value) | 7.306 |
| Molar refractivity | 122.07 cm3 |
| Topological polar surface area | 38.7 Å2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mustafa, N.H.; Sekar, M.; Fuloria, S.; Begum, M.Y.; Gan, S.H.; Rani, N.N.I.M.; Ravi, S.; Chidambaram, K.; Subramaniyan, V.; Sathasivam, K.V.; et al. Chemistry, Biosynthesis and Pharmacology of Sarsasapogenin: A Potential Natural Steroid Molecule for New Drug Design, Development and Therapy. Molecules 2022, 27, 2032. https://doi.org/10.3390/molecules27062032
Mustafa NH, Sekar M, Fuloria S, Begum MY, Gan SH, Rani NNIM, Ravi S, Chidambaram K, Subramaniyan V, Sathasivam KV, et al. Chemistry, Biosynthesis and Pharmacology of Sarsasapogenin: A Potential Natural Steroid Molecule for New Drug Design, Development and Therapy. Molecules. 2022; 27(6):2032. https://doi.org/10.3390/molecules27062032
Chicago/Turabian StyleMustafa, Nur Hanisah, Mahendran Sekar, Shivkanya Fuloria, M. Yasmin Begum, Siew Hua Gan, Nur Najihah Izzati Mat Rani, Subban Ravi, Kumarappan Chidambaram, Vetriselvan Subramaniyan, Kathiresan V. Sathasivam, and et al. 2022. "Chemistry, Biosynthesis and Pharmacology of Sarsasapogenin: A Potential Natural Steroid Molecule for New Drug Design, Development and Therapy" Molecules 27, no. 6: 2032. https://doi.org/10.3390/molecules27062032
APA StyleMustafa, N. H., Sekar, M., Fuloria, S., Begum, M. Y., Gan, S. H., Rani, N. N. I. M., Ravi, S., Chidambaram, K., Subramaniyan, V., Sathasivam, K. V., Jeyabalan, S., Uthirapathy, S., Ponnusankar, S., Lum, P. T., Bhalla, V., & Fuloria, N. K. (2022). Chemistry, Biosynthesis and Pharmacology of Sarsasapogenin: A Potential Natural Steroid Molecule for New Drug Design, Development and Therapy. Molecules, 27(6), 2032. https://doi.org/10.3390/molecules27062032