
Functional Protein Composition in Femoral Glands of Sand Lizards (Lacerta agilis)

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
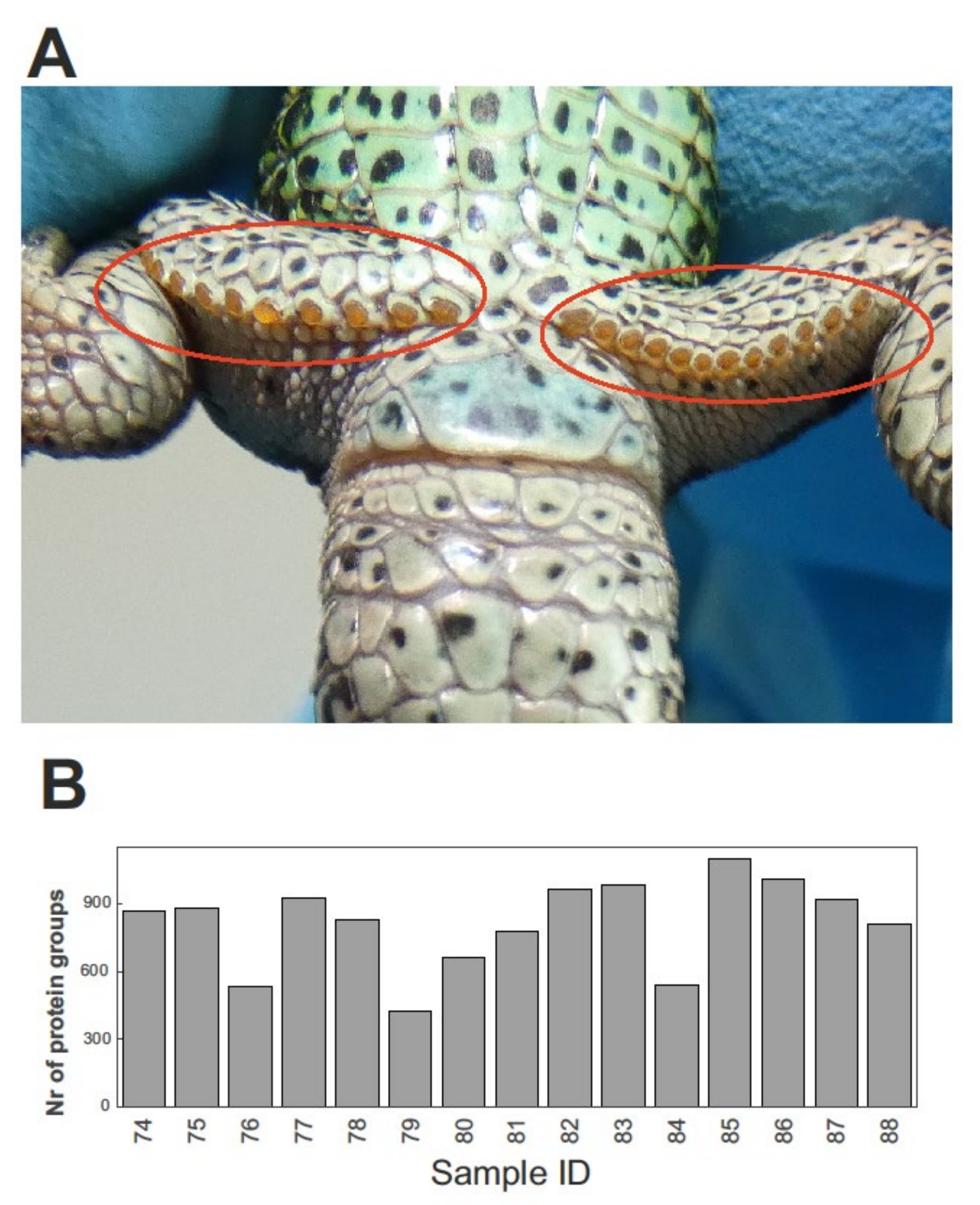
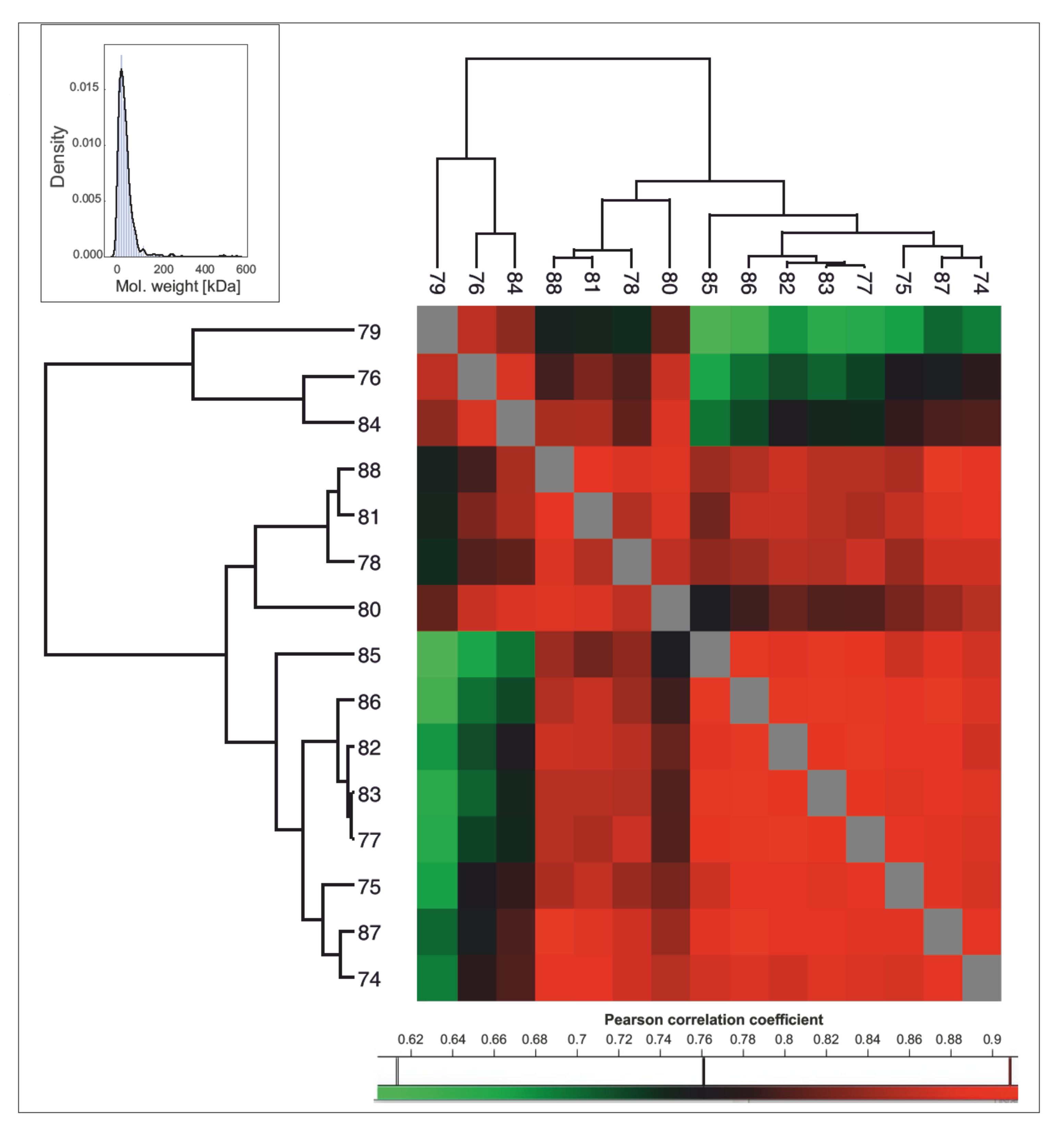
2.1. Protein Identification and Inter-Individual Variation
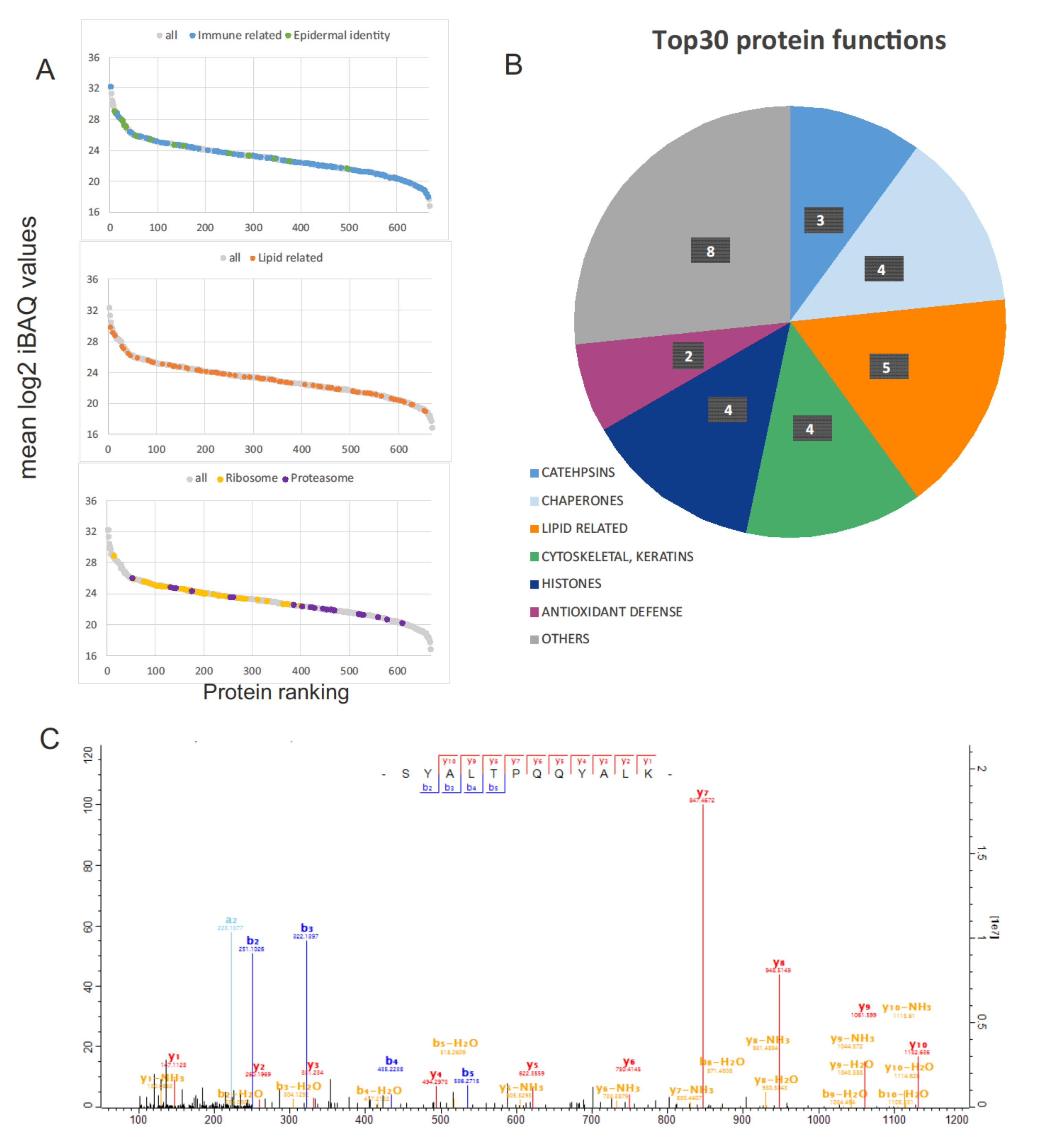
2.2. Protein Functionality and Comparison with the Galápagos Marine Iguana Proteome
2.3. Are Sand Lizard FG Proteins Involved in Intraspecific Chemical Communication?
3. Materials and Methods
3.1. Lizard Capture and Sampling
3.2. LC-MS/MS Analysis
3.3. LC-MS/MS Data Processing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Touhara, K. Sexual communication via peptide and protein pheromones. Curr. Opin. Pharmacol. 2008, 8, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.D. Proteins and peptides as pheromone signals and chemical signatures. Anim. Behav. 2014, 97, 273–280. [Google Scholar] [CrossRef]
- Roberts, S.A.; Simpson, D.M.; Armstrong, S.D.; Davidson, A.J.; Robertson, D.H.; McLean, L.; Beynon, R.J.; Hurst, J.L. Darcin: A male pheromone that stimulates female memory and sexual attraction to an individual male’s odour. BMC Biol. 2010, 8, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houck, L.D. Pheromone communication in amphibians and reptiles. Annu. Rev. Physiol. 2009, 71, 161–176. [Google Scholar] [CrossRef] [Green Version]
- Maex, M.; Van Bocxlaer, I.; Mortier, A.; Proost, P.; Bossuyt, F. Courtship pheromone use in a model urodele, the Mexican axolotl (Ambystoma mexicanum). Sci. Rep. 2016, 6, 20184. [Google Scholar] [CrossRef] [Green Version]
- Van Bocxlaer, I.; Maex, M.; Treer, D.; Janssenswillen, S.; Janssens, R.; Vandebergh, W.; Proost, P.; Bossuyt, F. Beyond sodefrin: Evidence for a multi-component pheromone system in the model newt Cynops pyrrhogaster (Salamandridae). Sci. Rep. 2016, 6, 21880. [Google Scholar] [CrossRef]
- Baeckens, S.; Herrel, A.; Broeckhoven, C.; Vasilopoulou-Kampitsi, M.; Huyghe, K.; Goyens, J.; Van Damme, R. Evolutionary morphology of the lizard chemosensory system. Sci. Rep. 2017, 7, 10141. [Google Scholar] [CrossRef] [Green Version]
- Cooper, W.E. Correlated evolution of prey chemical discrimination with foraging, lingual morphology and vomeronasal chemoreceptor abundance in lizards. Behav. Ecol. Sociobiol. 1997, 41, 257–265. [Google Scholar] [CrossRef]
- Schwenk, K. Of tongues and noses: Chemoreception in lizards and snakes. Trends Ecol. Evol. 1995, 10, 7–12. [Google Scholar] [CrossRef]
- Cole, C.J. Femoral glands in lizards: A review. Herpetologica 1966, 22, 199–206. [Google Scholar]
- Martín, J.; Lopez, P. Pheromones and chemical communication in lizards. In Reproductive Biology and Phylogeny of Lizards and Tuatara; Rheubert, J.L., Siegel, D.S., Trauth, S.E., Eds.; CRC Press: Boca Raton, FL, USA, 2014; Volume 10, pp. 43–77. [Google Scholar]
- Mayerl, C.; Baeckens, S.; Van Damme, R. Evolution and role of the follicular epidermal gland system in non-ophidian squamates. Amphibia-Reptilia 2015, 36, 185–206. [Google Scholar] [CrossRef] [Green Version]
- Matthey, R. Caractères sexuels secondaires du lézard mâle (‘Lacerta agilis’). Bull. Soc. Vaud. Sci. Nat. 1929, 57, 71–81. [Google Scholar]
- Baeckens, S.; Whiting, M.J. Investment in chemical signalling glands facilitates the evolution of sociality in lizards. Proc. R. Soc. B 2021, 288, 20202438. [Google Scholar] [CrossRef] [PubMed]
- Alberts, A.C.; Pratt, N.C.; Phillips, J.A. Seasonal productivity of lizard femoral glands: Relationship to social dominance and androgen levels. Physiol. Behav. 1992, 51, 729–733. [Google Scholar] [CrossRef]
- Carazo, P.; Font, E.; Desfilis, E. Chemosensory assessment of rival competitive ability and scent-mark function in a lizard, Podarcis hispanica. Anim. Behav. 2007, 74, 895–902. [Google Scholar] [CrossRef]
- Khannoon, E.; Breithaupt, T.; El-Gendy, A.; Hardege, J.D. Sexual differences in behavioural response to femoral gland pheromones of Acanthodactylus boskianus. Herpetol. J. 2010, 20, 225–229. [Google Scholar]
- Martín, J.; López, P. Chemoreception, symmetry and mate choice in lizards. Proc. R. Soc. B 2000, 267, 1265–1269. [Google Scholar] [CrossRef]
- Martín, J.; López, P. Condition-dependent pheromone signaling by male rock lizards: More oily scents are more attractive. Chem. Senses 2010, 35, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Alberts, A.C. Chemical properties of femoral gland secretions in the desert iguana, Dipsosaurus dorsalis. J. Chem. Ecol. 1990, 16, 13–25. [Google Scholar] [CrossRef]
- Baeckens, S. Evolution of animal chemical communication: Insights from non-model species and phylogenetic comparative methods. Belg. J. Zool. 2019, 149, 63–93. [Google Scholar] [CrossRef]
- Weldon, P.J.; Flachsbarth, B.; Schulz, S. Natural products from the integument of nonavian reptiles. Nat. Prod. Rep. 2008, 25, 738–756. [Google Scholar] [CrossRef]
- Alberts, A.C. Phylogenetic and adaptive variation in lizard femoral gland secretions. Copeia 1991, 1991, 69–79. [Google Scholar] [CrossRef]
- Mangiacotti, M.; Fumagalli, M.; Cagnone, M.; Viglio, S.; Bardoni, A.M.; Scali, S.; Sacchi, R. Morph-specific protein patterns in the femoral gland secretions of a colour polymorphic lizard. Sci. Rep. 2019, 9, 8412. [Google Scholar] [CrossRef]
- Mangiacotti, M.; Fumagalli, M.; Scali, S.; Zuffi, M.A.; Cagnone, M.; Salvini, R.; Sacchi, R. Inter-and intra-population variability of the protein content of femoral gland secretions from a lacertid lizard. Curr. Zool. 2017, 63, 657–665. [Google Scholar] [CrossRef] [Green Version]
- Mangiacotti, M.; Gaggiani, S.; Coladonato, A.J.; Scali, S.; Zuffi, M.A.L.; Sacchi, R. First experimental evidence that proteins from femoral glands convey identity-related information in a lizard. Acta Ethol. 2019, 22, 57–65. [Google Scholar] [CrossRef]
- Mangiacotti, M.; Baeckens, S.; Scali, S.; Martín, J.; Van Damme, R.; Sacchi, R. Evolutionary and biogeographical support for species-specific proteins in lizard chemical signals. Biol. J. Linn. Soc. 2021, 134, 912–928. [Google Scholar] [CrossRef]
- Tellkamp, F.; Lang, F.; Ibáñez, A.; Abraham, L.; Quezada, G.; Günther, S.; Looso, M.; Tann, F.J.; Müller, D.; Cemic, F.; et al. Proteomics of Galápagos marine iguanas links function of femoral gland proteins to the immune system. Mol. Cell. Proteom. 2020, 19, 1523–1532. [Google Scholar] [CrossRef]
- Blanke, I.; Fernley, H. The Sand Lizard—Between Light and Shadow; Laurenti-Verlag: Bielefeld, Germany, 2015. [Google Scholar]
- Olsson, M.; Madsen, T.; Nordby, J.; Wapstra, E.; Ujvari, B.; Wittsell, H. Major histocompatibility complex and mate choice in sand lizards. Proc. R. Soc. B 2003, 270, S254–S256. [Google Scholar] [CrossRef] [Green Version]
- Baeckens, S.; Martín, J.; García-Roa, R.; Pafilis, P.; Huyghe, K.; Van Damme, R. Environmental conditions shape the chemical signal design of lizards. Funct. Ecol. 2018, 32, 566–580. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized ppb-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Schwanhäusser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [Green Version]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Benes, P.; Vetvicka, V.; Fusek, M. Cathepsin D—Many functions of one aspartic protease. Crit. Rev. Oncol. Hematol. 2008, 68, 12–28. [Google Scholar] [CrossRef] [Green Version]
- Kolli, N.; Garman, S.C. Proteolytic activation of human cathepsin A. J. Biol. Chem. 2014, 289, 11592–11600. [Google Scholar] [CrossRef] [Green Version]
- Conus, S.; Simon, H.U. Cathepsins and their involvement in immune responses. Swiss Med. Wkly. 2010, 140, 4–11. [Google Scholar] [CrossRef]
- Reich, M.; Spindler, K.-D.; Burret, M.; Kalbacher, H.; Boehm, B.O.; Burster, T. Cathepsin A is expressed in primary human antigen-presenting cells. Immunol. Lett. 2010, 128, 143–147. [Google Scholar] [CrossRef]
- Hassan, M.I.; Shajee, B.; Waheed, A.; Ahmad, F.; Sly, W.S. Structure, function and applications of carbonic anhydrase isozymes. Bioorg. Med. Chem. 2013, 21, 1570–1582. [Google Scholar] [CrossRef]
- Gabirot, M.; López, P.; Martín, J.; De Fraipont, M.; Heulin, B.; Sinervo, B.; Clobert, J. Chemical composition of femoral secretions of oviparous and viviparous types of male common lizards Lacerta vivipara. Biochem. Syst. Ecol. 2008, 36, 539–544. [Google Scholar] [CrossRef]
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular organelles important in intercellular communication. J. Proteom. 2010, 73, 1907–1920. [Google Scholar] [CrossRef]
- Imparato, B.A.; Antoniazzi, M.M.; Rodrigues, M.T.; Jared, C. Morphology of the femoral glands in the lizard Ameiva ameiva (Teiidae) and their possible role in semiochemical dispersion. J. Morphol. 2007, 268, 636–648. [Google Scholar] [CrossRef]
- Khannoon, E.R.; Dollahon, N.R.; Bauer, A.M. Comparative study of the pheromone-manufacturing femoral glands in two sympatric species of lacertid lizards (Acanthodactylus). Zool. Sci. 2013, 30, 110–117. [Google Scholar] [CrossRef]
- Dietrich, M.A.; Irnazarow, I.; Adamek, M.; Jurecka, P.; Teich, L.; Rakus, K.; Kodzik, N.; Chadzińska, M.; Steinhagen, D.; Ciereszko, A. 2D-DIGE proteomic analysis of blood plasma reveals changes in immune-and stress-associated proteins following hormonal stimulation of carp males. Fish Shellfish Immunol. 2021, 118, 354–368. [Google Scholar] [CrossRef]
- Nynca, J.; Żarski, D.; Fröhlich, T.; Köster, M.; Bobe, J.; Ciereszko, A. Comparative proteomic analysis reveals the importance of the protective role of ovarian fluid over eggs during the reproduction of pikeperch. Aquaculture 2022, 548, 737656. [Google Scholar] [CrossRef]
- Rast, J.P.; D’Alessio, S.; Kraev, I.; Lange, S. Post-translational protein deimination signatures in sea lamprey (Petromyzon marinus) plasma and plasma-extracellular vesicles. Dev. Comp. Immunol. 2021, 125, 104225. [Google Scholar] [CrossRef]
- Tietgen, L.; Hagen, I.J.; Kleven, O.; Bernardi, C.D.; Kvalnes, T.; Norén, K.; Hasselgren, M.; Wallén, J.F.; Angerbjörn, A.; Landa, A.; et al. Fur colour in the Arctic fox: Genetic architecture and consequences for fitness. Proc. R. Soc. B 2021, 288, 20211452. [Google Scholar] [CrossRef]
- Penning, T.M.; Wangtrakuldee, P.; Auchus, R.J. Structural and functional biology of aldo-keto reductase steroid-transforming enzymes. Endocr. Rev. 2019, 40, 447–475. [Google Scholar] [CrossRef]
- Olsson, M.; Madsen, T.; Wapstra, E.; Silverin, B.; Ujvari, B.; Wittzell, H. MHC, health, color, and reproductive success in sand lizards. Behav. Ecol. Sociobiol. 2005, 58, 289–294. [Google Scholar] [CrossRef]
- Ruff, J.S.; Nelson, A.C.; Kubinak, J.L.; Potts, W.K. MHC signaling during social communication. In Self and Nonself; Springer: New York, NY, USA, 2012; pp. 290–313. [Google Scholar]
- Baeckens, S.; Martín, J.; García-Roa, R.; Van Damme, R. Sexual selection and the chemical signal design of lacertid lizards. Zool. J. Linn. Soc. 2018, 183, 445–457. [Google Scholar] [CrossRef] [Green Version]
- Kopena, R.; López, P.; Martín, J. Relative contribution of dietary carotenoids and vitamin E to visual and chemical sexual signals of male Iberian green lizards: An experimental test. Behav. Ecol. Sociobiol. 2014, 68, 571–581. [Google Scholar] [CrossRef]
- Chiva, C.; Olivella, R.; Borras, E.; Espadas, G.; Pastor, O.; Sole, A.; Sabido, E. QCloud: A cloud-based quality control system for mass spectrometry-based proteomics laboratories. PLoS ONE 2018, 13, e0189209. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote) omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar]
- STRING. Functional Protein Association Networks. Available online: https://string-db.org/ (accessed on 11 March 2021).
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- DAVID. Bioinformatics Resources (2021 Update). Available online: https://david.ncifcrf.gov/home.jsp (accessed on 18 March 2022).
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Protein ID | Protein Name | Gene Name | Length | Organism | IR * | IR in Tellkamp et al. ** | Comment |
|---|---|---|---|---|---|---|---|
| Cathepsins | |||||||
| D3JHM6_9SAUR | Cathepsin D (Fragment) | CTSD | 24 | Lacerta schreiberi | 1 | 10 | |
| A0A670IA46_PODMU | Peptidase A1 domain-containing protein | CTSD | 399 | Podarcis muralis | 3 | 10 | |
| A0A670IH97_PODMU | Carboxypeptidase (EC 3.4.16.-) | CTSA | 530 | Podarcis muralis | 14 | 112 | |
| Chaperones | |||||||
| A0A670J0X0_PODMU | Uncharacterized protein | PDIA6 | 449 | Podarcis muralis | 4 | not FG-specific *** | |
| A0A670KPF3_PODMU | 78 kDa glucose-regulated protein (Binding-immunoglobulin protein) (Heat shock protein 70 family protein 5) (Heat shock protein family A member 5) (Immunoglobulin heavy chain-binding protein) | HSPA5 | 654 | Podarcis muralis | 5 | 17 | |
| A0A670I8P5_PODMU | SHSP domain-containing protein | 169 | Podarcis muralis | 8 | not FG-specific *** | belongs to the small heat shock protein (HSP20) family | |
| A0A670K8T4_PODMU | Uncharacterized protein | HSPA8 | 646 | Podarcis muralis | 11 | not FG-specific *** | |
| Lipid related | |||||||
| A0A670JAW3_PODMU | GP-PDE domain-containing protein | GDPD3 | 318 | Podarcis muralis | 6 | not FG-specific *** | |
| A0A670JD81_PODMU | Beta-hexosaminidase (EC 3.2.1.52) | HEXA | 522 | Podarcis muralis | 10 | 105 | |
| A0A670HXK7_PODMU | Phospholipase A2 (EC 3.1.1.4) | 731 | Podarcis muralis | 24 | 158 | ||
| A0A670KKI2_PODMU | Aldo_ket_red domain-containing protein | 278 | Podarcis muralis | 28 | not FG-specific *** | members of aldo-keto reductase family are involved in steroid metabolic processes | |
| A0A670IG25_PODMU | Inositol-1-monophosphatase (EC 3.1.3.25) | IMPA1 | 347 | Podarcis muralis | 29 | not FG-specific *** | |
| Cytoskeletal, Keratins | |||||||
| A0A1D9CFN7_9SAUR | Beta-actin | 375 | Eremias argus | 7 | 261 | ||
| A0A670KC37_PODMU | IF rod domain-containing protein | LOC114593000 | 611 | Podarcis muralis | 9 | 4, 7, 11, 12, 25, 46, 109, 764 | keratin |
| A0A670JPK3_PODMU | IF rod domain-containing protein | LOC114581896 | 472 | Podarcis muralis | 27 | 4, 7, 11, 12, 25, 46, 109, 764 | keratin |
| A0A670JN74_PODMU | IF rod domain-containing protein | LOC114582343 | 462 | Podarcis muralis | 30 | 4, 7, 11, 12, 25, 46, 109, 764 | keratin |
| Histones | |||||||
| A0A670K9E9_PODMU | Histone H2A | LOC114582712 | 129 | Podarcis muralis | 15 | not FG-specific *** | |
| A0A670JD32_PODMU | Histone H3 | LOC114581771 | 136 | Podarcis muralis | 16 | not FG-specific *** | |
| A0A670K7U8_PODMU | Histone H4 | 101 | Podarcis muralis | 21 | not FG-specific *** | ||
| A0A670K3F4_PODMU | Histone H2B | LOC114581794 | 126 | Podarcis muralis | 22 | not FG-specific *** | |
| Antioxidant defense | |||||||
| A0A670I2N3_PODMU | Superoxide dismutase [Cu-Zn] (EC 1.15.1.1) | SOD1 | 155 | Podarcis muralis | 17 | not FG-specific *** | |
| A0A670I4X7_PODMU | Thioredoxin domain-containing protein | PRDX4 | 276 | Podarcis muralis | 25 | 113 | |
| Others | |||||||
| A0A670JGA4_PODMU | Carbonic anhydrase (EC 4.2.1.1) | LOC114583714 | 316 | Podarcis muralis | 2 | 2 | pH regulation |
| A0A670JY74_PODMU | Glyceraldehyde-3-phosphate dehydrogenase (EC 1.2.1.12) | GAPDH | 333 | Podarcis muralis | 12 | not FG-specific *** | |
| A0A670HW54_PODMU | 40S ribosomal protein S27a (Ubiquitin carboxyl extension protein 80) (Ubiquitin-40S ribosomal protein S27a) | RPS27A | 156 | Podarcis muralis | 13 | 204 | |
| A0A670ICE5_PODMU | Elongation factor 1-alpha | EEF1A1 | 462 | Podarcis muralis | 18 | 398 | |
| A0A670HUL4_PODMU | Voltage-dependent anion-selective channel protein 2 | VDAC2 | 283 | Podarcis muralis | 19 | not FG-specific *** | |
| A0A670HU92_PODMU | Mannosyl-glycoprotein endo-beta-N-acetylglucosaminidase (EC 3.2.1.96) | ENGASE | 811 | Podarcis muralis | 20 | 94 | glycoprotein modification |
| A0A670K0C8_PODMU | S_100 domain-containing protein | S100A14 | 102 | Podarcis muralis | 23 | 47 | cell survival and apoptosis/epidermal identity |
| A0A670JM37_PODMU | SERPIN domain-containing protein | SERPINE2 | 386 | Podarcis muralis | 26 | 416 | protease inhibitor |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibáñez, A.; Skupien-Rabian, B.; Jankowska, U.; Kędracka-Krok, S.; Zając, B.; Pabijan, M. Functional Protein Composition in Femoral Glands of Sand Lizards (Lacerta agilis). Molecules 2022, 27, 2371. https://doi.org/10.3390/molecules27072371
Ibáñez A, Skupien-Rabian B, Jankowska U, Kędracka-Krok S, Zając B, Pabijan M. Functional Protein Composition in Femoral Glands of Sand Lizards (Lacerta agilis). Molecules. 2022; 27(7):2371. https://doi.org/10.3390/molecules27072371
Chicago/Turabian StyleIbáñez, Alejandro, Bozena Skupien-Rabian, Urszula Jankowska, Sylwia Kędracka-Krok, Bartłomiej Zając, and Maciej Pabijan. 2022. "Functional Protein Composition in Femoral Glands of Sand Lizards (Lacerta agilis)" Molecules 27, no. 7: 2371. https://doi.org/10.3390/molecules27072371
APA StyleIbáñez, A., Skupien-Rabian, B., Jankowska, U., Kędracka-Krok, S., Zając, B., & Pabijan, M. (2022). Functional Protein Composition in Femoral Glands of Sand Lizards (Lacerta agilis). Molecules, 27(7), 2371. https://doi.org/10.3390/molecules27072371