
Artefact Profiling: Panomics Approaches for Understanding the Materiality of Written Artefacts


Abstract
:1. Introduction
2. Sampling Strategies
2.1. Sampling Strategies for High Resolution Omics Approaches That Can Be Performed on Site
2.2. Sampling Strategies for High Resolution Omics Approaches That Require No Sample Preparation but Must Be Performed in a Laboratory Environment
3. Paleogenomics and Metagenomics Analyses
3.1. Analytical Procedure for Genomics Analyses
3.2. Analysis of Endogenous DNA Sequences
3.3. Analysis of Exogenous DNA Sequences
4. Paleoproteomics and Metaproteomics Analyses
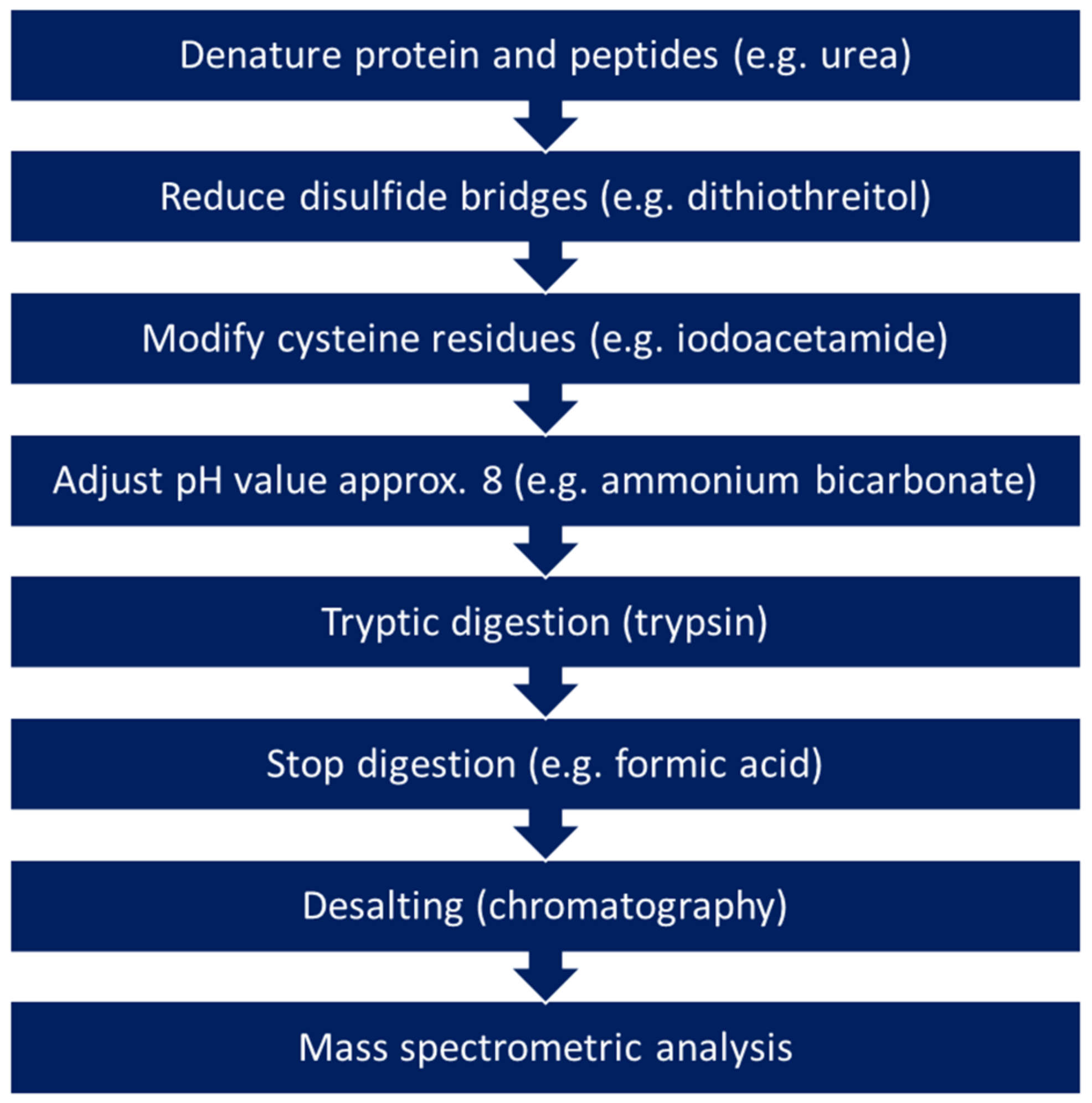
4.1. Analytical Procedure for Proteomics Analyses
4.2. Analysis of Endogenous Proteomics Sequences
4.3. Analysis of Exogenous Proteomics Sequences
5. Paleometabolomics and Metametabolomics Analyses
5.1. Analytical Procedure for Metabolomics Analyses

{kind=link}
{kind=link}
| Technology | Metabolite Coverage | Advantages | Disadvantages |
|---|---|---|---|
| GC-MS |
|
|
|
| LC-MS |
|
|
|
| NMR spectroscopy |
|
|
|
5.2. Analysis of Endogenous Metabolites
5.3. Analysis of Exogenous Metabolites
6. Metallomics and Isotopolomics Analyses
6.1. Analytical Procedure for Metallomics and Isotopolomics Analyses
6.2. Analysis of Endogenous Metals and Isotope Ratios
6.3. Analysis of Exogenous Metals and Isotope Ratios
7. Application of Panomics Strategies to Manuscript Research
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Piñar, G.; Sterflinger, K. Natural sciences at the service of art and cultural heritage: An interdisciplinary area in development and important challenges. Microb. Biotechnol. 2021, 14, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Fiddyment, S.; Teasdale, M.D.; Vnouček, J.; Lévêque, É.; Binois, A.; Collins, M.J. So you want to do biocodicology? A field guide to the biological analysis of parchment. Herit. Sci. 2019, 7, 35. [Google Scholar] [CrossRef] [Green Version]
- Creydt, M.; Fischer, M. Omics approaches for food authentication. Electrophoresis 2018, 39, 1569–1581. [Google Scholar] [CrossRef] [PubMed]
- D’Adamo, G.L.; Widdop, J.T.; Giles, E.M. The future is now? Clinical and translational aspects of “Omics” technologies. Immunol. Cell Biol. 2021, 99, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.; Braus, G.; Gemeinholzer, B.; Häuser, C.; Jahn, R.; Lohrmann, V.; Lüter, C.; Meyer, A.; Misof, B.; Raupach, M.; et al. Herausforderungen und Chancen der Integrativen Taxonomie für Forschung und Gesellschaft—Taxonomische Forschung im Zeitalter der OMICS-Technologien; Misof, B., Wägele, J.W., Eds.; Deutsche Akademie der Naturforscher Leopoldina e. V.: Halle, Germany, 2014; ISBN 978-3-8047-3281-0. [Google Scholar]
- Marvasi, M.; Cavalieri, D.; Mastromei, G.; Casaccia, A.; Perito, B. Omics technologies for an in-depth investigation of biodeterioration of cultural heritage. Int. Biodeterior. Biodegrad. 2019, 144, 104736. [Google Scholar] [CrossRef]
- Pyzik, A.; Ciuchcinski, K.; Dziurzynski, M.; Dziewit, L. The bad and the good—Microorganisms in cultural heritage environments—An update on biodeterioration and biotreatment approaches. Materials 2021, 14, 177. [Google Scholar] [CrossRef]
- Beata, G. The use of -omics tools for assessing biodeterioration of cultural heritage: A Review. J. Cult. Herit. 2020, 45, 351–361. [Google Scholar] [CrossRef]
- Vilanova, C.; Porcar, M. Art-omics: Multi-omics meet archaeology and art conservation. Microb. Biotechnol. 2020, 13, 435–441. [Google Scholar] [CrossRef] [Green Version]
- Sterflinger, K.; Pinar, G. Molecular-based techniques for the study of microbial communities in artworks. In Microorganisms in the Deterioration and Preservation of Cultural Heritage; Joseph, E., Ed.; Springer: Cham, Germany, 2021; pp. 59–77. ISBN 978-3-030-69410-4. [Google Scholar]
- Pinar, G.; Sterflinger, K. Two decades using molecular techniques to study biodeterioration of cultural heritage: An amazing biotechnological development. In Conserving Cultural Heritage; Mosquera, M.J., Almoraima Gil, M.L., Eds.; CRC Press/Balkema, Taylor & Francis Group: London, UK; Leiden, The Netherlands, 2018; pp. 299–302. ISBN 9781315158648. [Google Scholar]
- Zilberstein, G.; Zilberstein, R.; Zilberstein, S.; Maor, U.; Cohen-Ofri, I.; Shor, P.; Bitler, T.; Riestra, B.; Righetti, P.G. Proteomics and metabolomics composition of the ink of a letter in a fragment of a dead sea scroll from cave 11 (P1032-Fr0). J. Proteom. 2021, 249, 104370. [Google Scholar] [CrossRef]
- Creydt, M.; Fischer, M. Mass spectrometry-based proteomics and metaproteomics analysis of ancient manuscripts. In Exploring Written Artefacts, Objects, Methods, and Concepts; Quenzer, J.B., Ed.; De Gruyter: Hamburg, Germany; pp. 183–212. ISBN 9783110753301.
- Smith, O.; Gilbert, M.T.P. Ancient RNA. In Paleogenomics. Population Genomics; Lindqvist, C., Rajora, O.P., Eds.; Springer: Cham, Germany, 2019; pp. 53–74. ISBN 978-3-030-04753-5. [Google Scholar]
- Johnson, C.; Patterson, A.; Idle, J.; Gonzalez, F. Xenobiotic metabolomics: Major impact on the metabolome. Annu. Rev. Pharmacol. Toxicol. 2011, 52, 37–56. [Google Scholar] [CrossRef]
- Paul, A.; de Boves Harrington, P. Chemometric applications in metabolomic studies using chromatography-mass spectrometry. TrAC Trends Anal. Chem. 2021, 135, 116165. [Google Scholar] [CrossRef]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Burgio, L. Pigments, dyes and inks: Their analysis on manuscripts, scrolls and papyri. Archaeol. Anthropol. Sci. 2021, 13, 194. [Google Scholar] [CrossRef]
- Sharma, D.; Singh, M.R.; Dighe, B. Chromatographic study on traditional natural preservatives used for palm leaf manuscripts in india. Restaur. Int. J. Preserv. Libr. Arch. Mater. 2018, 39, 249–264. [Google Scholar] [CrossRef]
- Anava, S.; Neuhof, M.; Gingold, H.; Sagy, O.; Munters, A.; Svensson, E.M.; Afshinnekoo, E.; Danko, D.; Foox, J.; Shor, P.; et al. Illuminating genetic mysteries of the dead sea scrolls. Cell 2020, 181, 1218–1231. [Google Scholar] [CrossRef] [PubMed]
- Creydt, M.; Ludwig, L.; Köhl, M.; Fromm, J.; Fischer, M. Wood profiling by non-targeted high-resolution mass spectrometry: Part 1, Metabolite profiling in cedrela wood for the determination of the geographical origin. J. Chromatogr. A 2021, 1641, 461993. [Google Scholar] [CrossRef]
- Creydt, M.; Lautner, S.; Fromm, J.; Fischer, M. Wood Profiling by non-targeted liquid chromatography high-resolution mass spectrometry: Part 2, Detection of the geographical origin of spruce wood (Picea abies) by determination of metabolite pattern. J. Chromatogr. A 2022, 1663, 462737. [Google Scholar] [CrossRef]
- Migliore, L.; Thaller, M.C.; Vendittozzi, G.; Mejia, A.Y.; Mercuri, F.; Orlanducci, S.; Rubechini, A. Purple spot damage dynamics investigated by an integrated approach on a 1244 A.D. Parchment roll from the secret vatican archive. Sci. Rep. 2017, 7, 9521. [Google Scholar] [CrossRef]
- Perini, N.; Mercuri, F.; Orlanducci, S.; Thaller, M.C.; Migliore, L. The integration of metagenomics and chemical physical techniques biodecoded the buried traces of the biodeteriogens of parchment purple spots. Front. Microbiol. 2020, 11, 598945. [Google Scholar] [CrossRef]
- Raeisnia, N.; Arefian, E.; Amoozegar, M.A. Microbial community of an 11th century manuscript by both culture-dependent and -independent approaches. Microbiology 2022, 91, 313–323. [Google Scholar] [CrossRef]
- D’Amato, A.; Zilberstein, G.; Zilberstein, S.; Compagnoni, B.L.; Righetti, P.G. Of mice and men: Traces of life in the death registries of the 1630 plague in milano. J. Proteom. 2018, 180, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Zilberstein, G.; Maor, U.; Baskin, E.; D’Amato, A.; Righetti, P. Unearthing Bulgakov’s trace proteome from the master i margarita manuscript. J. Proteom. 2016, 152, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Zilberstein, G.; Maor, U.; Baskin, E.; Righetti, P.G. Maestro, marguerite, morphine: The last years in the life of Mikhail Bulgakov. J. Proteom. 2016, 131, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Zilberstein, G.; Zilberstein, S.; Maor, U.; Baskin, E.; D’Amato, A.; Righetti, P.G. De re metallica. Johannes Kepler and alchemy. Talanta 2019, 204, 82–88. [Google Scholar] [CrossRef]
- Saravayskaya, Y.; Zilberstein, G.; Zilberstein, R.; Zilberstein, S.; Maor, U.; D’Amato, A.; Righetti, P.G. “1984”: What Orwell could not predict. proteomic analysis of his scripts. Electrophoresis 2020, 41, 1931–1940. [Google Scholar] [CrossRef]
- Zilberstein, G.; Zilberstein, R.; Zilberstein, S.; Fau, G.; D’Amato, A.; Righetti, P.G. Il n’y a Pas d’amour heureux pour Casanova: Chemical- and bio-analysis of his memoirs. Electrophoresis 2019, 40, 3050–3056. [Google Scholar] [CrossRef]
- Zilberstein, G.; Zilberstein, S.; Righetti, P.G. Stalin’s “black dog”: A postmortem diagnosis. Anal. Bioanal. Chem. 2020, 412, 7701–7708. [Google Scholar] [CrossRef]
- Oliveira, F.M.; Araujo, C.A.R.; Macario, K.D.; Cid, A.S. Radiocarbon analysis of the torah scrolls from the national museum of brazil collection. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. Atoms 2015, 361, 531–534. [Google Scholar] [CrossRef]
- Jull, A.J.; Burr, G. Some interesting applications of radiocarbon dating to art and archaeology. Archeometriai Muh. 2014, 11, 139–148. [Google Scholar]
- Giussani, B.; Monticelli, D.; Rampazzi, L. Role of laser ablation–inductively coupled plasma–mass spectrometry in cultural heritage research: A review. Anal. Chim. Acta 2009, 635, 6–21. [Google Scholar] [CrossRef]
- Wagner, B.; Czajka, A. Non-invasive approximation of elemental composition of historic inks by LA-ICP-MS measurements of bathophenanthroline indicators. Talanta 2021, 222, 121520. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Jia, W. Multi-Omics Profiling: The way toward precision medicine in metabolic diseases. J. Mol. Cell Biol. 2021, 13, 576–593. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, C.; Qureshi, A.; Emili, A. Panomics for precision medicine. Trends Mol. Med. 2018, 24, 85–101. [Google Scholar] [CrossRef]
- Yılmaz, D. Archaeology as an interdisciplinary science at the cross-roads of physical, chemical, biological, and social sciences: New perspectives and research. In Transdisciplinarity; Rezaei, N., Ed.; Springer International Publishing: Cham, Germany, 2022; pp. 435–455. ISBN 978-3-030-94651-7. [Google Scholar]
- Roberts, J.; Robinson, P. Paleography and codicology. In A Companion to the History of the Book; Eliot, S., Rose, J., Eds.; Wiley: Chichester, UK, 2019; pp. 51–64. ISBN 9781119018193. [Google Scholar]
- Rehbein, M.; Sahle, P.; Schassan, T.; Assmann, B. Kodikologie nud Paläographie im Digitalen Zeitalter—Codicology and Palaeography in the Digital Age; Kodikologie und Paläographie im digitalen Zeitalter; BoD—Books on Demand: Norderstedt, Germany, 2009; ISBN 9783837098426. [Google Scholar]
- Online Etymology Dictionary, Definition: Paleo-. Available online: https://www.etymonline.com/word/paleo- (accessed on 29 May 2023).
- Online Etymology Dictionary, Definition: Pan-. Available online: https://www.etymonline.com/word/pan- (accessed on 29 May 2023).
- Creydt, M.; Fischer, M. Panomics—Fingerprinting approaches for food fraud detection. In Encyclopedia of Food Safety; Smithers, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2023; in press; ISBN 978-0-08-100596-5. [Google Scholar]
- Pinzari, F.; Montanari, M.; Michaelsen, A.; Pinar, G. Analytical protocols for the assessment of biological damage in historical documents. Coalit. Newsl. 2010, 19, 6–13. [Google Scholar]
- Piñar, G.; Sterflinger, K.; Pinzari, F. Unmasking the measles-like parchment discoloration: Molecular and microanalytical approach. Environ. Microbiol. 2015, 17, 427–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karakasidou, K.; Nikolouli, K.; Amoutzias, G.D.; Pournou, A.; Manassis, C.; Tsiamis, G.; Mossialos, D. Microbial diversity in biodeteriorated greek historical documents dating back to the 19th and 20th century: A Case Study. Microbiologyopen 2018, 7, e00596. [Google Scholar] [CrossRef]
- Piñar, G.; Tafer, H.; Schreiner, M.; Miklas, H.; Sterflinger, K. Decoding the biological information contained in two ancient slavonic parchment codices: An added historical value. Environ. Microbiol. 2020, 22, 3218–3233. [Google Scholar] [CrossRef]
- Rabin, I.; Hahn, O. Detection of fakes: The merits and limits of non-invasive materials analysis. In Fakes and Forgeries of Written Artefacts from Ancient Mesopotamia to Modern China; Michel, C., Friedrich, M., Eds.; De Gruyter: Berlin. Germany; Boston, MA, USA, 2020; pp. 281–290. ISBN 9783110714333. [Google Scholar]
- Paiva de Carvalho, H.; Sequeira, S.O.; Pinho, D.; Trovão, J.; da Costa, R.M.F.; Egas, C.; Macedo, M.F.; Portugal, A. Combining an innovative non-invasive sampling method and high-throughput sequencing to characterize fungal communities on a canvas painting. Int. Biodeterior. Biodegrad. 2019, 145, 104816. [Google Scholar] [CrossRef]
- Schulz, A.; Lautner, S.; Fromm, J.; Fischer, M. Not stealing from the treasure chest (or just a bit): Analyses on plant derived writing supports and non-invasive DNA sampling. PLoS ONE 2018, 13, e0198513. [Google Scholar] [CrossRef]
- Ding, X.; Lan, W.; Gu, J.-D. A review on sampling techniques and analytical methods for microbiota of cultural properties and historical architecture. Appl. Sci. 2020, 10, 8099. [Google Scholar] [CrossRef]
- Multari, D.H.; Ravishankar, P.; Sullivan, G.J.; Power, R.K.; Lord, C.; Fraser, J.A.; Haynes, P.A. Development of a novel minimally invasive sampling and analysis technique using skin sampling tape strips for bioarchaeological proteomics. J. Archaeol. Sci. 2022, 139, 105548. [Google Scholar] [CrossRef]
- Titubante, M.; Giannini, F.; Pasqualucci, A.; Romani, M.; Verona-Rinati, G.; Mazzuca, C.; Micheli, L. Towards a non-invasive approach for the characterization of arabic/christian manuscripts. Microchem. J. 2020, 155, 104684. [Google Scholar] [CrossRef]
- Fiddyment, S.; Holsinger, B.; Ruzzier, C.; Devine, A.; Binois, A.; Albarella, U.; Fischer, R.; Nichols, E.; Curtis, A.; Cheese, E.; et al. Animal origin of 13th-century uterine vellum revealed using noninvasive peptide fingerprinting. Proc. Natl. Acad. Sci. USA 2015, 112, 15066–15071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Righetti, P.G.; Zilberstein, G.; Zilberstein, S. New baits for fishing in cultural heritage’s Mare Magnum. J. Proteom. 2021, 235, 104113. [Google Scholar] [CrossRef] [PubMed]
- Righetti, P.G.; Zilberstein, G.; D’Amato, A. What Sherlock sorely missed: The EVA technology for cultural heritage exploration. Expert. Rev. Proteom. 2019, 16, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Calvano, C.D.; Rigante, E.; Picca, R.A.; Cataldi, T.R.I.; Sabbatini, L. An easily transferable protocol for in-situ quasi-non-invasive analysis of protein binders in works of art. Talanta 2020, 215, 120882. [Google Scholar] [CrossRef]
- Cicatiello, P.; Ntasi, G.; Rossi, M.; Marino, G.; Giardina, P.; Birolo, L. Minimally invasive and portable method for the identification of proteins in ancient paintings. Anal. Chem. 2018, 90, 10128–10133. [Google Scholar] [CrossRef]
- Ntasi, G.; Kirby, D.P.; Stanzione, I.; Carpentieri, A.; Somma, P.; Cicatiello, P.; Marino, G.; Giardina, P.; Birolo, L. A Versatile and user-friendly approach for the analysis of proteins in ancient and historical objects. J. Proteom. 2021, 231, 104039. [Google Scholar] [CrossRef]
- Kirby, D.P.; Manick, A.; Newman, R. Minimally invasive sampling of surface coatings for protein identification by peptide mass fingerprinting: A case study with photographs. J. Am. Inst. Conserv. 2020, 59, 235–245. [Google Scholar] [CrossRef]
- Newton, J.; Ramage, G.; Gadegaard, N.; Zachs, W.; Rogers, S.; Barrett, M.P.; Carruthers, G.; Burgess, K. Minimally-destructive atmospheric ionisation mass spectrometry authenticates authorship of historical manuscripts. Sci. Rep. 2018, 8, 10944. [Google Scholar] [CrossRef] [Green Version]
- McEwen, C.N.; McKay, R.G.; Larsen, B.S. Analysis of solids, liquids, and biological tissues using solids probe introduction at atmospheric pressure on commercial LC/MS instruments. Anal. Chem. 2005, 77, 7826–7831. [Google Scholar] [CrossRef] [PubMed]
- Creydt, M.; Fischer, M. Food Metabolomics: Latest hardware—Developments for nontargeted food authenticity and food safety testing. Electrophoresis 2022, 43, 2334–2350. [Google Scholar] [CrossRef]
- Hiraoka, K.; Ariyada, O.; Usmanov, D.T.; Chen, L.C.; Ninomiya, S.; Yoshimura, K.; Takeda, S.; Yu, Z.; Mandal, M.K.; Wada, H.; et al. Probe electrospray ionization (PESI) and its modified versions: Dipping PESI (DPESI), sheath-flow PESI (SfPESI) and adjustable SfPESI (Ad-SfPESI). Mass Spectrom. 2020, 9, A0092. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, K.; Nishidate, K.; Mori, K.; Asakawa, D.; Suzuki, S. Development of probe electrospray using a solid needle. Rapid Commun. Mass Spectrom. 2007, 21, 3139–3144. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, M.; Robotti, E.; Bearman, G.; France, F.; Barberis, E.; Shor, P.; Marengo, E. Direct analysis in real time mass spectrometry for the nondestructive investigation of conservation treatments of cultural heritage. J. Anal. Methods Chem. 2016, 2016, 6853591. [Google Scholar] [CrossRef] [Green Version]
- Adams, J. Analysis of printing and writing papers by using direct analysis in real time mass spectrometry. Int. J. Mass Spectrom. 2011, 301, 109–126. [Google Scholar] [CrossRef]
- Jones, R.W.; McClelland, J.F. Analysis of writing inks on paper using direct analysis in real time mass spectrometry. Forensic Sci. Int. 2013, 231, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Heaton, K.; Solazzo, C.; Collins, M.J.; Thomas-Oates, J.; Bergström, E.T. Towards the application of desorption electrospray ionisation mass spectrometry (DESI-MS) to the Analysis of ancient proteins from artefacts. J. Archaeol. Sci. 2009, 36, 2145–2154. [Google Scholar] [CrossRef]
- Sun, Q.; Luo, Y.; Wang, Y.; Zhang, Q.; Yang, X. Comparative analysis of aged documents by desorption electrospray ionization–mass spectrometry (DESI-MS) imaging. J. Forensic Sci. 2022, 67, 2062–2072. [Google Scholar] [CrossRef]
- Sun, Q.; Luo, Y.; Sun, N.; Zhang, Q.; Wang, Y.; Yang, X. Technical note: Analysis of biological substances in ink fingerprint by desorption electrospray ionization mass spectrometry. Forensic Sci. Int. 2022, 336, 111321. [Google Scholar] [CrossRef]
- Lee, G.; Cha, S. Depth-dependent chemical analysis of handwriting by nanospray desorption electrospray ionization mass spectrometry. J. Am. Soc. Mass Spectrom. 2021, 32, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, C.; Van Nuffel, S.; Walter, P.; Brunelle, A. Time-of-flight secondary ion mass spectrometry imaging in cultural heritage: A focus on old paintings. J. Mass Spectrom. 2022, 57, e4803. [Google Scholar] [CrossRef] [PubMed]
- Spoto, G. Secondary ion mass spectrometry in art and archaeology. Thermochim. Acta 2000, 365, 157–166. [Google Scholar] [CrossRef]
- He, A.; Karpuzov, D.; Xu, S. Ink identification by time-of-flight secondary ion mass spectroscopy. Surf. Interface Anal. 2006, 38, 854–858. [Google Scholar] [CrossRef]
- Attard-Montalto, N.; Ojeda, J.J.; Reynolds, A.; Ismail, M.; Bailey, M.; Doodkorte, L.; de Puit, M.; Jones, B.J. Determining the chronology of deposition of natural fingermarks and inks on paper using secondary ion mass spectrometry. Analyst 2014, 139, 4641–4653. [Google Scholar] [CrossRef] [Green Version]
- Coumbaros, J.; Kirkbride, K.P.; Klass, G.; Skinner, W. Application of time of flight secondary ion mass spectrometry to the in situ analysis of ballpoint pen inks on paper. Forensic Sci. Int. 2009, 193, 42–46. [Google Scholar] [CrossRef]
- Moore, K.L.; Barac, M.; Brajković, M.; Bailey, M.J.; Siketić, Z.; Bogdanović Radović, I. Determination of deposition order of toners, inkjet inks, and blue ballpoint pen combining mev-secondary ion mass spectrometry and particle induced X-ray emission. Anal. Chem. 2019, 91, 12997–13005. [Google Scholar] [CrossRef]
- Goacher, R.E.; DiFonzo, L.G.; Lesko, K.C. Challenges determining the correct deposition order of different intersecting black inks by time-of-flight secondary ion mass spectrometry. Anal. Chem. 2017, 89, 759–766. [Google Scholar] [CrossRef]
- Vilde, V.; Abel, M.-L.; Watts, J.F. A surface investigation of parchments using ToF-SIMS and PCA. Surf. Interface Anal. 2016, 48, 393–397. [Google Scholar] [CrossRef]
- Wagner, B.; Syta, O.; Sawicki, M. A moderate microsampling in laser ablation inductively coupled plasma mass spectrometry analysis of cultural heritage objects: A review. In Lasers in the Conservation of Artworks XI, Proceedings of Lacona XI; Targowski, P., Ed.; NCU Press: Toruń, Poland, 2017; pp. 155–178. [Google Scholar]
- Gonzalez, J.; Mao, X.L.; Roy, J.; Mao, S.S.; Russo, R.E. Comparison of 193, 213 and 266 nm laser ablation ICP-MS. J. Anal. At. Spectrom. 2002, 17, 1108–1113. [Google Scholar] [CrossRef]
- Horn, I.; von Blanckenburg, F. Investigation on elemental and isotopic fractionation during 196 nm femtosecond laser ablation multiple collector inductively coupled plasma mass spectrometry. Spectrochim. Acta Part B At. Spectrosc. 2007, 62, 410–422. [Google Scholar] [CrossRef]
- Burger, M.; Glaus, R.; Hubert, V.; van Willigen, S.; Wörle-Soares, M.; Convertini, F.; Lefranc, P.; Nielsen, E.; Günther, D. Novel sampling techniques for trace element quantification in ancient copper artifacts using laser ablation inductively coupled plasma mass spectrometry. J. Archaeol. Sci. 2017, 82, 62–71. [Google Scholar] [CrossRef]
- Knaf, A.; Londero, P.; Nikkel, J.; Hark, R.; Bezur, A. Novel portable laser ablation micro-sampling in cultural Heritage. Microsc. Microanal. 2021, 27, 3014–3016. [Google Scholar] [CrossRef]
- Wagner, B.; Bulska, E. On the use of laser ablation inductively coupled plasma mass spectrometry for the investigation of the written heritage. J. Anal. At. Spectrom. 2004, 19, 1325–1329. [Google Scholar] [CrossRef]
- Glaus, R.; Koch, J.; Günther, D. Portable laser ablation sampling device for elemental fingerprinting of objects outside the laboratory with laser ablation inductively coupled plasma mass spectrometry. Anal. Chem. 2012, 84, 5358–5364. [Google Scholar] [CrossRef] [PubMed]
- Glaus, R.; Dorta, L.; Zhang, Z.; Ma, Q.; Berke, H.; Günther, D. Isotope ratio determination of objects in the field by portable laser ablation sampling and subsequent multicollector ICPMS. J. Anal. At. Spectrom. 2013, 28, 801–809. [Google Scholar] [CrossRef]
- Kradolfer, S.; Heutschi, K.; Koch, J.; Günther, D. Tracking mass removal of portable laser ablation sampling by its acoustic response. Spectrochim. Acta Part B At. Spectrosc. 2021, 179, 106118. [Google Scholar] [CrossRef]
- Merkel, S.W.; D’Imporzano, P.; van Zuilen, K.; Kershaw, J.; Davies, G.R. “Non-invasive” portable laser ablation sampling for lead isotope analysis of archaeological silver: A comparison with bulk and in situ laser ablation techniques. J. Anal. At. Spectrom. 2022, 37, 148–156. [Google Scholar] [CrossRef]
- Seman, S.; Dussubieux, L.; Cloquet, C.; Pryce, T.O. Strontium isotope analysis in ancient glass from south asia using portable laser ablation sampling. Archaeometry 2021, 63, 88–104. [Google Scholar] [CrossRef]
- Numrich, M.; Schwall, C.; Lockhoff, N.; Nikolentzos, K.; Konstantinidi-Syvridi, E.; Cultraro, M.; Horejs, B.; Pernicka, E. Portable laser ablation sheds light on early bronze age gold treasures in the old world: New insights from troy, poliochni, and related finds. J. Archaeol. Sci. 2023, 149, 105694. [Google Scholar] [CrossRef]
- Venter, A.; Nefliu, M.; Graham Cooks, R. Ambient desorption ionization mass spectrometry. TrAC Trends Anal. Chem. 2008, 27, 284–290. [Google Scholar] [CrossRef]
- Lawal, R.O.; Donnarumma, F.; Murray, K.K. Deep-ultraviolet laser ablation electrospray ionization mass spectrometry. J. Mass Spectrom. 2019, 54, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Voß, H.; Moritz, M.; Pelczar, P.; Gagliani, N.; Huber, S.; Nippert, V.; Schlüter, H.; Hahn, J. Tissue sampling and homogenization with NIRL enables spatially resolved cell layer specific proteomic analysis of the murine intestine. Int. J. Mol. Sci. 2022, 23, 6132. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-C.; Chang, T.-L.; Li, C.-H.; Su, K.-W.; Liu, C.-C. Thermally stable and uniform DNA amplification with picosecond laser ablated graphene rapid thermal cycling device. Biosens. Bioelectron. 2019, 146, 111581. [Google Scholar] [CrossRef] [PubMed]
- Moretti, P.; Iwanicka, M.; Melessanaki, K.; Dimitroulaki, E.; Kokkinaki, O.; Daugherty, M.; Sylwestrzak, M.; Pouli, P.; Targowski, P.; van den Berg, K.J.; et al. Laser cleaning of paintings: In situ optimization of operative parameters through non-invasive assessment by optical coherence tomography (OCT), Reflection FT-IR spectroscopy and laser induced fluorescence spectroscopy (LIF). Herit. Sci. 2019, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Lan, T.; Lindqvist, C. Paleogenomics: Genome-scale analysis of ancient DNA and population and evolutionary genomic inferences. In Population Genomics: Concepts, Approaches and Applications; Rajora, O.P., Ed.; Springer: Cham, Germany, 2019; pp. 323–360. ISBN 978-3-030-04589-0. [Google Scholar]
- Heintzman, P.; Soares, A.; Chang, D.; Shapiro, B. Paleogenomics. Rev. Cell Biol. Mol. Med. 2015, 1, 243–267. [Google Scholar] [CrossRef]
- Dabney, J.; Meyer, M.; Pääbo, S. Ancient DNA damage. Cold Spring Harb. Perspect. Biol. 2013, 5, a012567. [Google Scholar] [CrossRef]
- Orlando, L.; Allaby, R.; Skoglund, P.; Der Sarkissian, C.; Stockhammer, P.W.; Ávila-Arcos, M.C.; Fu, Q.; Krause, J.; Willerslev, E.; Stone, A.C.; et al. Ancient DNA analysis. Nat. Rev. Methods Prim. 2021, 1, 14. [Google Scholar] [CrossRef]
- Andreeva, T.V.; Malyarchuk, A.B.; Soshkina, A.D.; Dudko, N.A.; Plotnikova, M.Y.; Rogaev, E.I. Methodologies for ancient DNA extraction from bones for genomic analysis: Approaches and guidelines. Russ. J. Genet. 2022, 58, 1017–1035. [Google Scholar] [CrossRef]
- Renaud, G.; Schubert, M.; Sawyer, S.; Orlando, L. Authentication and assessment of contamination in ancient DNA. Methods Mol. Biol. 2019, 1963, 163–194. [Google Scholar]
- Jónsson, H.; Ginolhac, A.; Schubert, M.; Johnson, P.L.F.; Orlando, L. MapDamage2.0: Fast approximate bayesian estimates of ancient DNA damage parameters. Bioinformatics 2013, 29, 1682–1684. [Google Scholar] [CrossRef] [Green Version]
- Fulton, T.; Shapiro, B. Setting up an ancient DNA laboratory. In Ancient DNA. Methods in Molecular Biology, Vol 1963; Shapiro, B., Barlow, A., Heintzman, P., Hofreiter, M., Paijmans, J., Soares, A., Eds.; Humana Press: New York, NY, USA, 2019; pp. 1–13. ISBN 978-1-61779-516-9. [Google Scholar]
- Matsvay, A.D.; Alborova, I.E.; Pimkina, E.V.; Markelov, M.L.; Khafizov, K.; Mustafin, K.K. Experimental approaches for ancient DNA extraction and sample preparation for next generation sequencing in ultra-clean conditions. Conserv. Genet. Resour. 2019, 11, 345–353. [Google Scholar] [CrossRef]
- Peyrégne, S.; Peter, B.M. AuthentiCT: A model of ancient DNA damage to estimate the proportion of present-day DNA contamination. Genome Biol. 2020, 21, 246. [Google Scholar] [CrossRef] [PubMed]
- Neukamm, J.; Peltzer, A.; Nieselt, K. DamageProfiler: Fast damage pattern calculation for ancient DNA. Bioinformatics 2021, 37, 3652–3653. [Google Scholar] [CrossRef]
- Peyrégne, S.; Prüfer, K. Present-Day DNA Contamination in ancient DNA datasets. BioEssays 2020, 42, 2000081. [Google Scholar] [CrossRef]
- Lech, T. Ancient DNA in historical parchments—Identifying a procedure for extraction and amplification of genetic material. Genet. Mol. Res. 2016, 15, gmr8661. [Google Scholar] [CrossRef] [PubMed]
- Marota, I.; Basile, C.; Ubaldi, M.; Rollo, F. DNA decay rate in papyri and human remains from egyptian archaeological sites. Am. J. Phys. Anthropol. 2002, 117, 310–318. [Google Scholar] [CrossRef]
- Barta, J.L.; Monroe, C.; Teisberg, J.E.; Winters, M.; Flanigan, K.; Kemp, B.M. One of the key characteristics of ancient DNA, low copy number, may be a product of its extraction. J. Archaeol. Sci. 2014, 46, 281–289. [Google Scholar] [CrossRef]
- Rohland, N.; Hofreiter, M. Comparison and optimization of ancient DNA extraction. Biotechniques 2007, 42, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Gamba, C.; Hanghøj, K.; Gaunitz, C.; Alfarhan, A.H.; Alquraishi, S.A.; Al-Rasheid, K.A.S.; Bradley, D.G.; Orlando, L. Comparing the performance of three ancient DNA extraction methods for high-throughput sequencing. Mol. Ecol. Resour. 2016, 16, 459–469. [Google Scholar] [CrossRef]
- Shapiro, B.; Hofreiter, M. Ancient DNA: Methods and Protocols; Methods in Molecular Biology; Humana Press Incorporated: Totowa, NJ, USA, 2012. [Google Scholar]
- Briggs, A.W.; Stenzel, U.; Meyer, M.; Krause, J.; Kircher, M.; Pääbo, S. Removal of deaminated cytosines and detection of in vivo methylation in ancient DNA. Nucleic Acids Res. 2010, 38, e87. [Google Scholar] [CrossRef] [Green Version]
- Pervez, M.T.; Hasnain, M.J.U.; Abbas, S.H.; Moustafa, M.F.; Aslam, N.; Shah, S.S.M. A comprehensive review of performance of next-generation sequencing platforms. Biomed Res. Int. 2022, 2022, 3457806. [Google Scholar] [CrossRef] [PubMed]
- Elizabeth Pennisi, A $100 Genome? New DNA Sequencers Could Be a ‘Game Changer’ for Biology, Medicine. Available online: https://www.science.org/content/article/100-genome-new-dna-sequencers-could-be-game-changer-biology-medicine (accessed on 21 December 2022).
- Schlecht, U.; Mok, J.; Dallett, C.; Berka, J. ConcatSeq: A method for increasing throughput of single molecule sequencing by concatenating short DNA fragments. Sci. Rep. 2017, 7, 5252. [Google Scholar] [CrossRef] [Green Version]
- Pavlovic, J.; Cavalieri, D.; Mastromei, G.; Pangallo, D.; Perito, B.; Marvasi, M. MinION Technology for microbiome sequencing applications for the conservation of cultural heritage. Microbiol. Res. 2021, 247, 126727. [Google Scholar] [CrossRef] [PubMed]
- Piñar, G.; Poyntner, C.; Lopandic, K.; Tafer, H.; Sterflinger, K. Rapid diagnosis of biological colonization in cultural artefacts using the MinION Nanopore sequencing technology. Int. Biodeterior. Biodegrad. 2020, 148, 104908. [Google Scholar] [CrossRef]
- Knapp, M.; Hofreiter, M. Next generation sequencing of ancient DNA: Requirements, strategies and perspectives. Genes 2010, 1, 227–243. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.; Kircher, M. Illumina sequencing library preparation for highly multiplexed target capture and sequencing. Cold Spring Harb. Protoc. 2010, 2010, pdb-prot5448. [Google Scholar] [CrossRef]
- Kircher, M.; Sawyer, S.; Meyer, M. Double indexing overcomes inaccuracies in multiplex sequencing on the illumina platform. Nucleic Acids Res. 2012, 40, e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gansauge, M.-T.; Meyer, M. Single-stranded DNA library preparation for the sequencing of ancient or damaged DNA. Nat. Protoc. 2013, 8, 737–748. [Google Scholar] [CrossRef]
- Gansauge, M.-T.; Gerber, T.; Glocke, I.; Korlevic, P.; Lippik, L.; Nagel, S.; Riehl, L.M.; Schmidt, A.; Meyer, M. Single-stranded DNA library preparation from highly degraded DNA using T4 DNA Ligase. Nucleic Acids Res. 2017, 45, e79. [Google Scholar] [CrossRef] [Green Version]
- Gansauge, M.-T.; Aximu-Petri, A.; Nagel, S.; Meyer, M. Manual and automated preparation of single-stranded DNA libraries for the sequencing of DNA from ancient biological remains and other sources of highly degraded DNA. Nat. Protoc. 2020, 15, 2279–2300. [Google Scholar] [CrossRef] [PubMed]
- Kapp, J.D.; Green, R.E.; Shapiro, B. A fast and efficient single-stranded genomic library preparation method optimized for ancient DNA. J. Hered. 2021, 112, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Carøe, C.; Gopalakrishnan, S.; Vinner, L.; Mak, S.S.T.; Sinding, M.H.S.; Samaniego, J.A.; Wales, N.; Sicheritz-Pontén, T.; Gilbert, M.T.P. Single-tube library preparation for degraded DNA. Methods Ecol. Evol. 2018, 9, 410–419. [Google Scholar] [CrossRef] [Green Version]
- Suchan, T.; Kusliy, M.A.; Khan, N.; Chauvey, L.; Tonasso-Calvière, L.; Schiavinato, S.; Southon, J.; Keller, M.; Kitagawa, K.; Krause, J.; et al. Performance and automation of ancient DNA Capture with RNA HyRAD probes. Mol. Ecol. Resour. 2022, 22, 891–907. [Google Scholar] [CrossRef] [PubMed]
- Vai, S.; Lari, M.; Caramelli, D. DNA sequencing in cultural heritage. In Analytical Chemistry for Cultural Heritage; Mazzeo, R., Ed.; Springer: Cham, Germany, 2017; pp. 329–346. ISBN 978-3-319-52804-5. [Google Scholar]
- Teasdale, M.D.; Fiddyment, S.; Vnouček, J.; Mattiangeli, V.; Speller, C.; Binois, A.; Carver, M.; Dand, C.; Newfield, T.P.; Webb, C.C.; et al. The york gospels: A 1000-year biological palimpsest. R. Soc. Open Sci. 2021, 4, 170988. [Google Scholar] [CrossRef] [Green Version]
- Teasdale, M.D.; van Doorn, N.L.; Fiddyment, S.; Webb, C.C.; O’Connor, T.; Hofreiter, M.; Collins, M.J.; Bradley, D.G. Paging through history: Parchment as a reservoir of ancient DNA for next generation sequencing. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2015, 370, 20130379. [Google Scholar] [CrossRef]
- Shepherd, L.D.; Whitehead, P.; Whitehead, A. Genetic analysis identifies the missing parchment of New Zealand’s founding document, the treaty of Waitangi. PLoS ONE 2019, 14, e0210528. [Google Scholar] [CrossRef]
- Cappa, F.; Piñar, G.; Brenner, S.; Frühmann, B.; Wetter, W.; Schreiner, M.; Engel, P.; Miklas, H.; Sterflinger, K. The Kiev Folia: An interdisciplinary approach to unravelling the past of an ancient Slavonic manuscript. Int. Biodeterior. Biodegrad. 2022, 167, 105342. [Google Scholar] [CrossRef]
- Wagner, S.; Lagane, F.; Seguin-Orlando, A.; Schubert, M.; Leroy, T.; Guichoux, E.; Chancerel, E.; Bech-Hebelstrup, I.; Bernard, V.; Billard, C.; et al. High-throughput DNA sequencing of ancient wood. Mol. Ecol. 2018, 27, 1138–1154. [Google Scholar] [CrossRef] [Green Version]
- Lendvay, B.; Hartmann, M.; Brodbeck, S.; Nievergelt, D.; Reinig, F.; Zoller, S.; Parducci, L.; Gugerli, F.; Büntgen, U.; Sperisen, C. Improved recovery of ancient DNA from subfossil wood—Application to the world’s oldest late glacial pine forest. New Phytol. 2018, 217, 1737–1748. [Google Scholar] [CrossRef] [Green Version]
- Boesi, A. Paper plants in the Tibetan world: A Preliminary study. In Tibetan Printing: Comparison, Continuities, and Change; Diemberger, H., Ehrhard, K., Kornicki, P.F., Eds.; Brill: Leiden, The Netherlands; Boston, MA, USA, 2016; pp. 501–530. ISBN 978-90-04-31625-6. [Google Scholar]
- Cappitelli, F.; Pasquariello, G.; Tarsitani, G.; Sorlini, C. Scripta manent? Assessing microbial risk to paper heritage. Trends Microbiol. 2010, 18, 538–542. [Google Scholar] [CrossRef]
- Pinheiro, A.C.; Sequeira, S.O.; Macedo, M.F. Fungi in archives, libraries, and museums: A Review on paper conservation and human health. Crit. Rev. Microbiol. 2019, 45, 686–700. [Google Scholar] [CrossRef] [PubMed]
- Okpalanozie, O.E.; Adebusoye, S.A.; Troiano, F.; Cattò, C.; Ilori, M.O.; Cappitelli, F. Assessment of indoor air environment of a Nigerian museum library and its biodeteriorated books using culture-dependent and –independent techniques. Int. Biodeterior. Biodegrad. 2018, 132, 139–149. [Google Scholar] [CrossRef]
- Lech, T. Evaluation of a parchment document, the 13th century incorporation charter for the city of Krakow, Poland, for microbial hazards. Appl. Environ. Microbiol. 2016, 82, 2620–2631. [Google Scholar] [CrossRef] [Green Version]
- Ruppert, K.M.; Kline, R.J.; Rahman, M.S. Past, present, and future perspectives of environmental DNA (eDNA) metabarcoding: A systematic review in methods, monitoring, and applications of global eDNA. Glob. Ecol. Conserv. 2019, 17, e00547. [Google Scholar] [CrossRef]
- Pérez-Cobas, A.E.; Gomez-Valero, L.; Buchrieser, C. Metagenomic approaches in microbial ecology: An update on whole-genome and marker gene sequencing analyses. Microb. Genom. 2020, 6, mgen000409. [Google Scholar] [CrossRef]
- Piñar, G.; Cappa, F.; Vetter, W.; Schreiner, M.; Miklas, H.; Sterflinger, K. Complementary strategies for deciphering the information contained in ancient parchment documentary materials. Appl. Sci. 2022, 12, 10479. [Google Scholar] [CrossRef]
- Pinheiro, C.; Miller, A.Z.; Vaz, P.; Caldeira, A.T.; Casanova, C. Underneath the purple stain. Heritage 2022, 5, 4100–4113. [Google Scholar] [CrossRef]
- Melo, D.; Sequeira, S.O.; Lopes, J.A.; Macedo, M.F. Stains versus colourants produced by fungi colonising paper cultural heritage: A review. J. Cult. Herit. 2019, 35, 161–182. [Google Scholar] [CrossRef]
- Mazzoli, R.; Giuffrida, M.G.; Pessione, E. Back to the Past: “Find the guilty bug-microorganisms involved in the biodeterioration of archeological and historical artifacts. ” Appl. Microbiol. Biotechnol. 2018, 102, 6393–6407. [Google Scholar] [CrossRef]
- Oetari, A.; Susetyo-Salim, T.; Sjamsuridzal, W.; Suherman, E.A.; Monica, M.; Wongso, R.; Fitri, R.; Nurlaili, D.G.; Ayu, D.C.; Teja, T.P. Occurrence of fungi on deteriorated old dluwang manuscripts from Indonesia. Int. Biodeterior. Biodegrad. 2016, 114, 94–103. [Google Scholar] [CrossRef]
- Szulc, J.; Otlewska, A.; Ruman, T.; Kubiak, K.; Karbowska-Berent, J.; Kozielec, T.; Gutarowska, B. Analysis of paper foxing by newly available omics techniques. Int. Biodeterior. Biodegrad. 2018, 132, 157–165. [Google Scholar] [CrossRef]
- Kraková, L.; Šoltys, K.; Otlewska, A.; Pietrzak, K.; Purkrtová, S.; Savická, D.; Puškárová, A.; Bučková, M.; Szemes, T.; Budiš, J.; et al. Comparison of methods for identification of microbial communities in book collections: Culture-dependent (sequencing and MALDI-TOF MS) and culture-independent (Illumina MiSeq). Int. Biodeterior. Biodegrad. 2018, 131, 51–59. [Google Scholar] [CrossRef]
- Taha, A.S.; Salem, M.Z.M.; Abo Elgat, W.A.A.; Ali, H.M.; Hatamleh, A.A.; Abdel-Salam, E.M. Assessment of the impact of different treatments on the technological and antifungal properties of papyrus (Cyperus papyrus L.) sheets. Materials 2019, 12, 620. [Google Scholar] [CrossRef] [Green Version]
- Saada, H.; Othman, M.; Khaleil, M. Mold-deteriorated archaeological Egyptian papyri: Biodeteriogens, monitoring the deterioration, and treatment approach. Archaeometry 2022, 65, 335–353. [Google Scholar] [CrossRef]
- Šoltys, K.; Planý, M.; Biocca, P.; Vianello, V.; Bučková, M.; Puškárová, A.; Sclocchi, M.C.; Colaizzi, P.; Bicchieri, M.; Pangallo, D.; et al. Lead soaps formation and biodiversity in a XVIII century wax seal coloured with minium. Environ. Microbiol. 2020, 22, 1517–1534. [Google Scholar] [CrossRef]
- Szulc, J.; Jablonskaja, I.; Jabłońska, E.; Ruman, T.; Karbowska-Berent, J.; Gutarowska, B. Metabolomics and metagenomics characteristic of historic beeswax seals. Int. Biodeterior. Biodegradation 2020, 152, 105012. [Google Scholar] [CrossRef]
- Pavlović, J.; Sclocchi, M.C.; Planý, M.; Ruggiero, D.; Puškárová, A.; Bučková, M.; Šoltys, K.; Colaizzi, P.; Riccardi, M.L.; Pangallo, D.; et al. The microbiome of candle beeswax drops on ancient manuscripts. Int. Biodeterior. Biodegrad. 2022, 174, 105482. [Google Scholar] [CrossRef]
- Ostrom, P.H.; Schall, M.; Gandhi, H.; Shen, T.-L.; Hauschka, P.V.; Strahler, J.R.; Gage, D.A. New strategies for characterizing ancient proteins using matrix-assisted laser desorption ionization mass spectrometry. Geochim. Cosmochim. Acta 2000, 64, 1043–1050. [Google Scholar] [CrossRef]
- Warinner, C.; Korzow Richter, K.; Collins, M.J. Paleoproteomics. Chem. Rev. 2022, 122, 13401–13446. [Google Scholar] [CrossRef]
- Hendy, J. Ancient protein analysis in archaeology. Sci. Adv. 2021, 7, eabb9314. [Google Scholar] [CrossRef] [PubMed]
- Demarchi, B. Mechanisms of degradation and survival. In Amino Acids and Proteins in Fossil Biominerals; Demarchi, B., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 23–42. ISBN 9781119089537. [Google Scholar]
- Ramsøe, A.; van Heekeren, V.; Ponce, P.; Fischer, R.; Barnes, I.; Speller, C.; Collins, M.J. DeamiDATE 1.0: Site-specific deamidation as a tool to assess authenticity of members of ancient proteomes. J. Archaeol. Sci. 2020, 115, 105080. [Google Scholar] [CrossRef]
- Leo, G.; Bonaduce, I.; Andreotti, A.; Marino, G.; Pucci, P.; Colombini, M.P.; Birolo, L. Deamidation at asparagine and glutamine as a major modification upon deterioration/aging of proteinaceous binders in mural paintings. Anal. Chem. 2011, 83, 2056–2064. [Google Scholar] [CrossRef]
- Schroeter, E.; Cleland, T. Glutamine deamidation: An indicator of antiquity, or preservational quality? Rapid Commun. Mass Spectrom. 2015, 30, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Ying, Y.; Li, H. Recent progress in the analysis of protein deamidation using mass spectrometry. Methods 2022, 200, 42–57. [Google Scholar] [CrossRef]
- Nair, B.; Palomo, I.R.; Markussen, B.; Wiuf, C.; Fiddyment, S.; Collins, M.J. Parchment glutamine index (PQI): A novel method to estimate glutamine deamidation levels in parchment collagen obtained from low-quality MALDI-TOF data. bioRxiv 2022. [Google Scholar] [CrossRef]
- Toniolo, L.; D’Amato, A.; Saccenti, R.; Gulotta, D.; Righetti, P.G. The Silk Road, Marco Polo, a Bible and its proteome: A detective story. J. Proteom. 2012, 75, 3365–3373. [Google Scholar] [CrossRef] [PubMed]
- Demarchi, B. Ancient protein sequences. In Amino Acids and Proteins in Fossil Biominerals; Demarchi, B., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 113–126. ISBN 9781119089537. [Google Scholar]
- Haugg, S.; Creydt, M.; Zierold, R.; Fischer, M.; Blick, R.H. Booster-microchannel plate (BMCP) detector for signal amplification in MALDI-TOF mass spectrometry for ions beyond m/z 50,000. Phys. Chem. Chem. Phys. 2023, 25, 7312–7322. [Google Scholar] [CrossRef]
- Miller, R.M.; Smith, L.M. Overview and considerations in bottom-up proteomics. Analyst 2023, 148, 475–486. [Google Scholar] [CrossRef]
- Pandeswari, P.B.; Sabareesh, V. Middle-down approach: A choice to sequence and characterize proteins/proteomes by mass spectrometry. RSC Adv. 2019, 9, 313–344. [Google Scholar] [CrossRef] [Green Version]
- Soltwisch, J.; Kettling, H.; Vens-Cappell, S.; Niehaus, M.; Müthing, J.; Dreisewerd, K. Mass spectrometry imaging with laser-induced postionization. Science 2015, 348, 211. [Google Scholar] [CrossRef]
- Potthoff, A.; Dreisewerd, K.; Soltwisch, J. Detailed characterization of the postionization efficiencies in MALDI-2 as a function of relevant input parameters. J. Am. Soc. Mass Spectrom. 2020, 31, 1844–1853. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Jones, B.R. Nano-LC: An updated review. Biomed. Chromatogr. 2022, 36, e5317. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S. Developments, advancements, and contributions of mass spectrometry in omics technologies. In Advances in Protein Molecular and Structural Biology Methods; Tripathi, T., Dubey, V., Eds.; Academic Press: London, UK, 2022; pp. 327–356. ISBN 978-0-323-90264-9. [Google Scholar]
- Nys, G.; Nix, C.; Cobraiville, G.; Servais, A.-C.; Fillet, M. Enhancing protein discoverability by data independent acquisition assisted by ion mobility mass spectrometry. Talanta 2020, 213, 120812. [Google Scholar] [CrossRef]
- Zarrouk, E.; Lenski, M.; Bruno, C.; Thibert, V.; Contreras, P.; Privat, K.; Ameline, A.; Fabresse, N. High-resolution mass spectrometry: Theoretical and technological aspects. Toxicol. Anal. Clin. 2022, 34, 3–18. [Google Scholar] [CrossRef]
- Creydt, M.; Fischer, M. Food phenotyping: Recording and processing of non-targeted liquid chromatography mass spectrometry data for verifying food authenticity. Molecules 2020, 25, 3972. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.-L.; Hu, Z.-L.; Zhang, S.; Qing, Y.; Fragasso, A.; Maglia, G.; Meller, A.; Bayley, H.; Dekker, C.; Long, Y.-T. Nanopore-based technologies beyond DNA sequencing. Nat. Nanotechnol. 2022, 17, 1136–1146. [Google Scholar] [CrossRef]
- Kirby, D.P.; Buckley, M.; Promise, E.; Trauger, S.A.; Holdcraft, T.R. Identification of collagen-based materials in cultural heritage. Analyst 2013, 138, 4849–4858. [Google Scholar] [CrossRef]
- Buckley, M.; Whitcher Kansa, S.; Howard, S.; Campbell, S.; Thomas-Oates, J.; Collins, M. Distinguishing between archaeological sheep and goat bones using a single collagen peptide. J. Archaeol. Sci. 2010, 37, 13–20. [Google Scholar] [CrossRef]
- Buckley, M.; Fraser, S.; Herman, J.; Melton, N.D.; Mulville, J.; Pálsdóttir, A.H. Species identification of archaeological marine mammals using collagen fingerprinting. J. Archaeol. Sci. 2014, 41, 631–641. [Google Scholar] [CrossRef]
- Hickinbotham, S.; Fiddyment, S.; Stinson, T.L.; Collins, M.J. How to get your goat: Automated identification of species from MALDI-ToF spectra. Bioinformatics 2020, 36, 3719–3725. [Google Scholar] [CrossRef] [Green Version]
- Ruffini-Ronzani, N.; Nieus, J.-F.; Soncin, S.; Hickinbotham, S.; Dieu, M.; Bouhy, J.; Charles, C.; Ruzzier, C.; Falmagne, T.; Hermand, X.; et al. A biocodicological analysis of the medieval library and archive from orval abbey, Belgium. R. Soc. Open Sci. 2021, 8, 210210. [Google Scholar] [CrossRef]
- Sommer, D.; Mühlen Axelsson, K.; Collins, M.; Fiddyment, S.; Bredal-Jørgensen, J.; Simonsen, K.; Lauridsen, C.; Larsen, R. Multiple microanalyses of a sample from the Vinland map. Archaeometry 2017, 59, 287–301. [Google Scholar] [CrossRef]
- Calà, E.; Agostino, A.; Fenoglio, G.; Capra, V.; Porticelli, F.; Manzari, F.; Fiddyment, S.; Aceto, M. The Messale Rosselli: Scientific investigation on an outstanding 14th century illuminated manuscript from Avignon. J. Archaeol. Sci. Rep. 2019, 23, 721–730. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, C.; Rabitsch, S.; Vnoucek, J.; Fiddyment, S.; Quandt, A.; Collins, M.; Kanold, I.; Aceto, M.; Melo, M.; Calà, E.; et al. The Vienna Genesis: Material Analysis and Conservation of a Late Antique Illuminated Manuscript on Purple Parchment; Hofmann, C., Ed.; Böhlau: Wien, Austria, 2020; ISBN 978-320521058-0. [Google Scholar]
- Fiddyment, S.; Goodison, N.J.; Brenner, E.; Signorello, S.; Price, K.; Collins, M.J. Girding the loins? Direct evidence of the use of a medieval parchment birthing girdle from biomolecular analysis. R. Soc. Open Sci. 2021, 8, 202055. [Google Scholar] [CrossRef]
- Doherty, S.P.; Henderson, S.; Fiddyment, S.; Finch, J.; Collins, M.J. Scratching the surface: The use of sheepskin parchment to deter textual erasure in early modern legal deeds. Herit. Sci. 2021, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- van der Werf, I.D.; Calvano, C.D.; Germinario, G.; Cataldi, T.R.I.; Sabbatini, L. Chemical characterization of medieval illuminated parchment scrolls. Microchem. J. 2017, 134, 146–153. [Google Scholar] [CrossRef]
- Han, B.; Niang, J.; Rao, H.; Lyu, N.; Oda, H.; Sakamoto, S.; Yang, Y.; Sablier, M. Paper fragments from the Tibetan Samye Monastery: Clues for an unusual sizing recipe implying wheat starch and milk in early Tibetan papermaking. J. Archaeol. Sci. Reports 2021, 36, 102793. [Google Scholar] [CrossRef]
- Zilberstein, G.; Zilberstein, S.; Maor, U.; Righetti, P.G. Surface analysis of ancient parchments via the EVA film: The Aleppo Codex. Anal. Biochem. 2020, 604, 113824. [Google Scholar] [CrossRef] [PubMed]
- Ashfaq, M.Y.; Da’na, D.A.; Al-Ghouti, M.A. Application of MALDI-TOF MS for identification of environmental bacteria: A review. J. Environ. Manag. 2022, 305, 114359. [Google Scholar] [CrossRef] [PubMed]
- Avataneo, C.; Sablier, M. New Criteria for the characterization of traditional East Asian papers. Environ. Sci. Pollut. Res. Int. 2017, 24, 2166–2181. [Google Scholar] [CrossRef]
- Han, B.; Vial, J.; Sakamoto, S.; Sablier, M. Identification of traditional East Asian handmade papers through the multivariate data analysis of pyrolysis-GC/MS Data. Analyst 2019, 144, 1230–1244. [Google Scholar] [CrossRef] [PubMed]
- Corsaro, C.; Mallamace, D.; Łojewska, J.; Mallamace, F.; Pietronero, L.; Missori, M. Molecular degradation of ancient documents revealed by 1H HR-MAS NMR spectroscopy. Sci. Rep. 2013, 3, 2896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, I.K.; Neme Tauil, R.; Valacco, M.P.; Moreno, S.; Siracusano, G.; Maier, M.S. Investigation of proteins in samples of a mid-18th century colonial mural painting by MALDI-TOF/MS and LC-ESI/MS (Orbitrap). Microchem. J. 2018, 143, 457–466. [Google Scholar] [CrossRef]
- Lattuati-Derieux, A.; Bonnassies-Termes, S.; Lavédrine, B. Characterisation of compounds emitted during natural and artificial ageing of a book. Use of headspace-solid-phase microextraction/gas chromatography/mass spectrometry. J. Cult. Herit. 2006, 7, 123–133. [Google Scholar] [CrossRef]
- Stringer, K.A.; McKay, R.T.; Karnovsky, A.; Quémerais, B.; Lacy, P. Metabolomics and its application to acute lung diseases. Front. Immunol. 2016, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S. NMR metabolomics: A look ahead. J. Magn. Reson. 2019, 306, 155–161. [Google Scholar] [CrossRef]
- Letertre, M.P.M.; Dervilly, G.; Giraudeau, P. Combined nuclear magnetic resonance spectroscopy and mass spectrometry approaches for metabolomics. Anal. Chem. 2021, 93, 500–518. [Google Scholar] [CrossRef]
- Blümich, B.; Singh, K. Desktop NMR and its applications from materials science to organic chemistry. Angew. Chem. Int. Ed. 2018, 57, 6996–7010. [Google Scholar] [CrossRef]
- Rehorn, C.; Bluemich, B. Cultural heritage studies with mobile NMR. Angew. Chem. Int. Ed. 2018, 57, 7304–7312. [Google Scholar] [CrossRef]
- Manso, M.; Carvalho, M.L. Application of spectroscopic techniques for the study of paper documents: A survey. Spectrochim. Acta Part B At. Spectrosc. 2009, 64, 482–490. [Google Scholar] [CrossRef]
- Mirsaeidi, M.; Banoei, M.; Winston, B.; Schraufnagel, D. Metabolomics: Applications and promise in mycobacterial disease. Ann. Am. Thorac. Soc. 2015, 12, 1278–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, D.; Lee, M.; Kim, S.; Kim, K. Applications of NMR spectroscopy based metabolomics: A review. J. Korean Magn. Reson. Soc. 2013, 17, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Emwas, A.-H.; Roy, R.; McKay, R.T.; Tenori, L.; Saccenti, E.; Gowda, G.A.N.; Raftery, D.; Alahmari, F.; Jaremko, L.; Jaremko, M.; et al. NMR spectroscopy for metabolomics research. Metabolites 2019, 9, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogolitsyna, A.; Becker, M.; Dupont, A.-L.; Borgards, A.; Rosenau, T.; Potthast, A. Determination of carbohydrate- and lignin-derived components in complex effluents from cellulose processing by capillary electrophoresis with electrospray ionization-mass spectrometric detection. J. Chromatogr. A 2011, 1218, 8561–8566. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Yang, Q.; Liu, J.; Lee, Z. Evaluating volatile organic compounds from chinese traditional handmade paper by SPME-GC/MS. Herit. Sci. 2021, 9, 153. [Google Scholar] [CrossRef]
- Mallamace, D.; Vasi, S.; Missori, M.; Mallamace, F.; Corsaro, C. NMR Investigation of degradation processes of ancient and modern paper at different hydration levels. Front. Phys. 2017, 13, 138202. [Google Scholar] [CrossRef]
- Kaszonyi, A.; Izsák, L.; Králik, M.; Jablonsky, M. Accelerated and natural aging of cellulose-based paper: Py-GC/MS method. Molecules 2022, 27, 2855. [Google Scholar] [CrossRef]
- Ortiz-Herrero, L.; Blanco, M.E.; García-Ruiz, C.; Bartolomé, L. Direct and indirect approaches based on paper analysis by Py-GC/MS for estimating the age of documents. J. Anal. Appl. Pyrolysis 2018, 131, 9–16. [Google Scholar] [CrossRef]
- Lucejko, J.J.; Colombini, M.P.; Ribechini, E. Chemical alteration patterns of ancient Egyptian papyri studied by pyrolysis-GC/MS with in Situ Silylation. J. Anal. Appl. Pyrolysis 2020, 152, 104967. [Google Scholar] [CrossRef]
- Sebestyén, Z.; Badea, E.; Carsote, C.; Czégény, Z.; Szabó, T.; Babinszki, B.; Bozi, J.; Jakab, E. Characterization of historical leather bookbindings by various thermal methods (TG/MS, Py-GC/MS, and micro-DSC) and FTIR-ATR spectroscopy. J. Anal. Appl. Pyrolysis 2022, 162, 105428. [Google Scholar] [CrossRef]
- Sebestyén, Z.; Czégény, Z.; Badea, E.; Carsote, C.; Şendrea, C.; Barta-Rajnai, E.; Bozi, J.; Miu, L.; Jakab, E. Thermal characterization of new, artificially aged and historical leather and parchment. J. Anal. Appl. Pyrolysis 2015, 115, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Vilmont, L.-B.; Kim, H.-J.; Lavédrine, B.; Sakamoto, S.; Sablier, M. Characterization of Korean handmade papers collected in a hanji reference book. Herit. Sci. 2021, 9, 96. [Google Scholar] [CrossRef]
- Degano, I.; La Nasa, J. Trends in high performance liquid chromatography for cultural heritage. Top. Curr. Chem. 2016, 374, 20. [Google Scholar] [CrossRef] [PubMed]
- Calà, E.; Benzi, M.; Gosetti, F.; Zanin, A.; Gulmini, M.; Idone, A.; Serafini, I.; Ciccola, A.; Curini, R.; Whitworth, I.; et al. Towards the identification of the lichen species in historical orchil dyes by HPLC-MS/MS. Microchem. J. 2019, 150, 104140. [Google Scholar] [CrossRef]
- Barberis, E.; Manfredi, M.; Zilberstein, G.; Zilberstein, S.; Righetti, P. fiat lux... How Alessandro Volta illuminated his scripts. Comptes Rendus. Chim. 2021, 24, 361–371. [Google Scholar] [CrossRef]
- Sawoszczuk, T.; Syguła-Cholewińska, J.; del Hoyo-Meléndez, J.M. Application of HS-SPME-GC-MS method for the detection of active moulds on historical parchment. Anal. Bioanal. Chem. 2017, 409, 2297–2307. [Google Scholar] [CrossRef]
- Zilberstein, G.; Zilberstein, S.; Rocco, R.M.; Righetti, P.G. Jack London and White Fang: A lost struggle. Comptes Rendus. Chim. 2022, 25, 115–123. [Google Scholar] [CrossRef]
- Lazic, V.; Vadrucci, M.; Fantoni, R.; Chiari, M.; Mazzinghi, A.; Gorghinian, A. Applications of laser induced breakdown spectroscopy for cultural heritage: A comparison with XRF and PIXE techniques. Spectrochim. Acta Part B At. Spectrosc. 2018, 149, 1–14. [Google Scholar] [CrossRef]
- Ishii, K. PIXE and its applications to elemental analysis. Quantum Beam Sci. 2019, 3, 12. [Google Scholar] [CrossRef] [Green Version]
- Rawat, K.; Sharma, N.; Singh, V.K. X-ray fluorescence and comparison with other analytical methods (AAS, ICP-AES, LA-ICP-MS, IC, LIBS, SEM-EDS, and XRD). In X-ray Fluorescence in Biological Sciences: Principles, Instrumentation, and Applications; Singh, V.K., Kawai, J., Tripathi, D.K., Eds.; Wiley: Hoboken, NJ, USA, 2022; pp. 1–20. ISBN 9781119645719. [Google Scholar]
- Watteeuw, L.; van Bos, M.; Gersten, T.; Vandermeulen, B.; Hameeuw, H. An applied complementary use of macro X-ray fluorescence scanning and multi-light reflectance imaging to study medieval illuminated manuscripts. The Rijmbijbel of Jacob van Maerlant. Microchem. J. 2020, 155, 104582. [Google Scholar] [CrossRef]
- Mazzinghi, A.; Ruberto, C.; Castelli, L.; Ricciardi, P.; Czelusniak, C.; Giuntini, L.; Mandò, P.A.; Manetti, M.; Palla, L.; Taccetti, F. The importance of being little: MA-XRF on manuscripts on a venetian island. X-ray Spectrom. 2021, 50, 272–278. [Google Scholar] [CrossRef]
- Melo, M.J.; Otero, V.; Nabais, P.; Teixeira, N.; Pina, F.; Casanova, C.; Fragoso, S.; Sequeira, S.O. Iron-gall inks: A review of their degradation mechanisms and conservation treatments. Herit. Sci. 2022, 10, 145. [Google Scholar] [CrossRef]
- Cappa, F.; Sterflinger, K. Non-invasive physico-chemical and biological analysis of parchment manuscripts—An overview. Int. J. Preserv. Libr. Arch. Mater. 2022, 43, 127–142. [Google Scholar] [CrossRef]
- Brill, R.H.; Felker-Dennis, C.; Shirahata, H.; Joel, E.C. Lead Isotope Analyses of Some Chinese and Central Asian Pigments; Conservation of Ancient Sites on the Silk Road; The Getty Conservation Institute: Los Angeles, CA, USA, 1997; pp. 369–378. [Google Scholar]
- Nord, A.G.; Billström, K. Isotopes in cultural heritage: Present and future possibilities. Herit. Sci. 2018, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Dolgin, B.; Chen, Y.; Bulatov, V.; Schechter, I. Use of LIBS for rapid characterization of parchment. Anal. Bioanal. Chem. 2006, 386, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Manso, M.; Costa, M.; Carvalho, M.L. X-ray fluorescence spectrometry on paper characterization: A case study on XVIII and XIX century documents. Spectrochim. Acta Part B At. Spectrosc. 2008, 63, 1320–1323. [Google Scholar] [CrossRef]
- Romano, F.P.; Puglia, E.; Caliri, C.; Pavone, D.P.; Alessandrelli, M.; Busacca, A.; Fatuzzo, C.G.; Fleischer, K.J.; Pernigotti, C.; Preisler, Z.; et al. Layout of ancient Greek papyri through lead-drawn ruling lines revealed by macro X-ray fluorescence imaging. Sci. Rep. 2023, 13, 6582. [Google Scholar] [CrossRef]


| Term | Explanation |
|---|---|
| Archaeometry | Application of physics, chemistry, geology, and engineering sciences to analyse various archaeological materials [39]. |
| Biocodicology | Application of genomics and proteomics approaches to ancient manuscripts [2]. |
| Codicology | Analysis of the technical craft aspects and physical properties of a manuscript [40,41]. |
| Omics/Paleo-omics | Comparative analysis of different elemental or molecular entireties [3]. In the context of studies on ancient manuscripts or artefacts, the focus is on the analysis of the actual object, e.g., the writing surfaces or the inks used. The prefix “paleo” means old or ancient [5,42]. |
| Meta-omics | The focus of meta-omics studies is not the object itself, but the residues of other organism, e.g., microorganisms or the authors and readers [11]. Depending on the omics strategy chosen, the terms meta-genomics, meta-proteomics or meta-metabolomics are used, for example. |
| Paleography | The term refers to the study of ancient writings to be able to make spatial or temporal classifications. The focus is, for example, on the forms of the letters, spelling, or the use of typical abbreviations [40,41]. |
| Panomics | The prefix “pan” comes from the Greek and means all, every, or whole [43]. The term panomics is used for personalised medical questions or when analyzing food. It is a networked, symbiotic approach based on genomics, transcriptomics, proteomics, and metabolomics-based data [38,44]. |
| Scientific Issue | Omics Technology | Analytical Method | Reference |
|---|---|---|---|
| Characterization of Parchment | Genomics | NGS | [55] |
| Proteomics | LC-MS and MALDI-TOF | ||
| Characterization of Parchment | Genomics | NGS | [133] |
| Proteomics | MALDI-TOF | ||
| Characterization of Parchment | Genomics | NGS | [134] |
| Proteomics | MALDI-TOF | ||
| Characterization of Parchment | Genomics | NGS | [136] |
| Metallomics | FTIR, XRF, MSI | ||
| Characterization of Parchment | Proteomics | MALDI-TOF, FTIR | [185] |
| Metabolomics | Amino acid analysis, FTIR, GC-MS | ||
| Metallomics | FTIR, Raman, Energy dispersive X-ray spectroscopy | ||
| Organic composition of parchment and paint binders | Proteomicse | MADLI-TOF | [190] |
| Metabolomics | GC-MS | ||
| Studying the manufacturing process of Tibetan paper | Proteomicse | LC-MS | [191] |
| Metabolomics | GC-MS | ||
| Isotopolomics | Radiocarbon Dating | ||
| Composition of ink binders | Proteomics | MALDI-TOF | [12] |
| Metabolomics | GC-MS and LC-MS | ||
| Analysis of paper foxing | Genomics | NGS | [151] |
| Metabolomics | SALDI-TOF | ||
| Identification of microbial communities in book collections | Genomics | Culture-dependent analysis and NGS | [152] |
| Proteomics | MALDI-TOF | ||
| Disease and treatment of nephrotic syndrome by the author Mikhail Bulgakov | Proteomics | LC-MS | [27,28,56] |
| Metabolomics | GC-MS | ||
| Infection of Casanova with gonorrhea bacteria | Proteomics | LC-MS | [31] |
| Metallomics | Special mini-Hg-sensor | ||
| Investigating the microbiological and metabolic diversity of beeswax seals | Genomics | NGS | [156] |
| Metabolomics | SALDI-TOF | ||
| Characterization and study of the biodeterioration as well as the associated microbiome of a wax seal | Genomics | Culture-dependent analysis and NGS | [155] |
| Metabolomics | Raman, FTIR | ||
| Metallomics | Energy dispersive X-ray spectroscopy |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Creydt, M.; Fischer, M. Artefact Profiling: Panomics Approaches for Understanding the Materiality of Written Artefacts. Molecules 2023, 28, 4872. https://doi.org/10.3390/molecules28124872
Creydt M, Fischer M. Artefact Profiling: Panomics Approaches for Understanding the Materiality of Written Artefacts. Molecules. 2023; 28(12):4872. https://doi.org/10.3390/molecules28124872
Chicago/Turabian StyleCreydt, Marina, and Markus Fischer. 2023. "Artefact Profiling: Panomics Approaches for Understanding the Materiality of Written Artefacts" Molecules 28, no. 12: 4872. https://doi.org/10.3390/molecules28124872
APA StyleCreydt, M., & Fischer, M. (2023). Artefact Profiling: Panomics Approaches for Understanding the Materiality of Written Artefacts. Molecules, 28(12), 4872. https://doi.org/10.3390/molecules28124872