
The Protective Effects of Water-Soluble Alginic Acid on the N-Terminal of Thymopentin

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Preliminary Structural Analysis of WSAA
2.2. Schematic Diagrams of WSAA–TP5 Complex
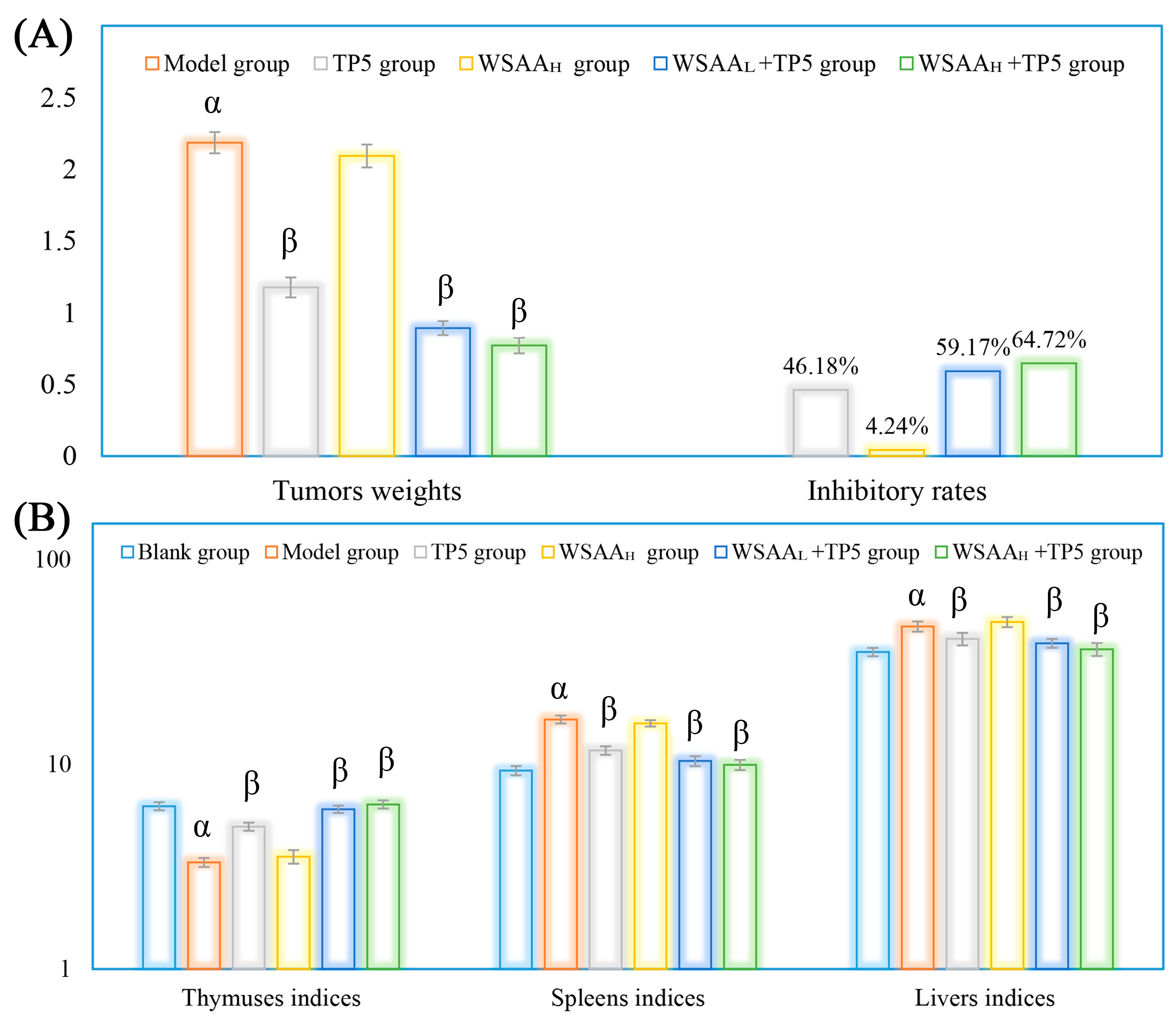
2.3. Organs Indices and Tumors Inhibitory Rates
2.4. Routine Analysis of Blood
2.5. Lymphocyte Subsets Detection
2.6. Lymphocytes Proliferation Activities and Antibodies Levels
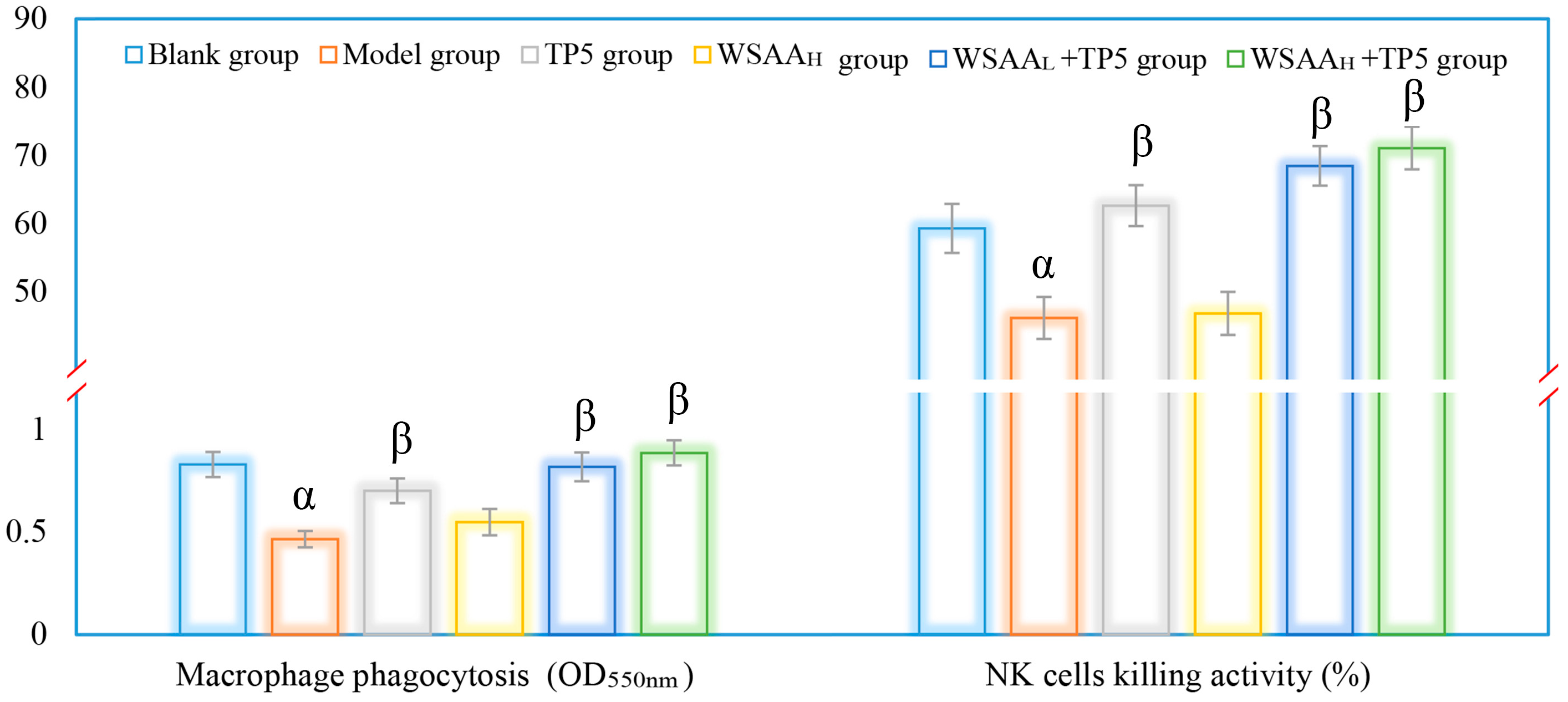
2.7. Macrophages and NK Cells Activities
2.8. Cytokines Levels in Sera
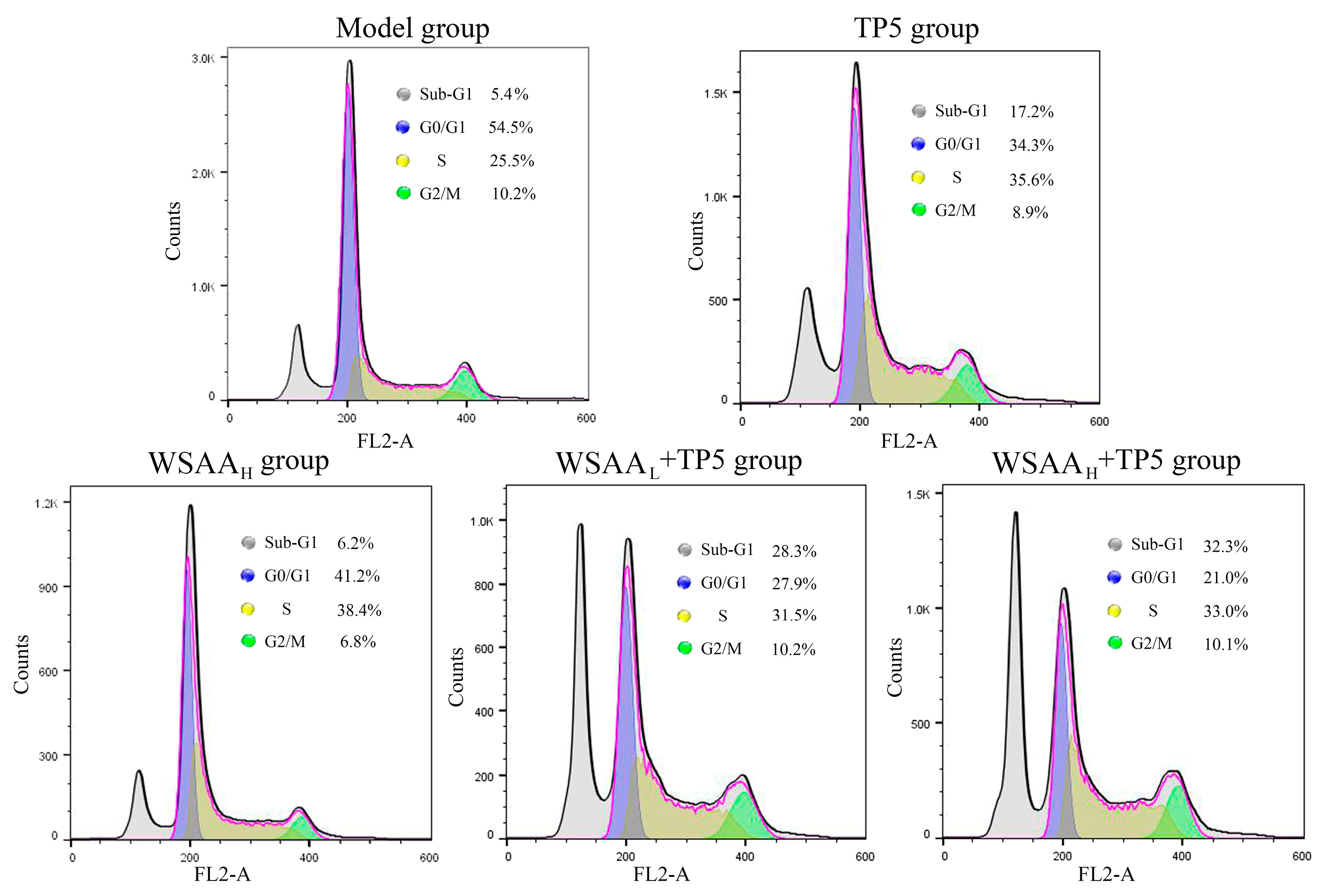
2.9. Cell Cycle Determination
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Preparation of WSAA
4.3. Molecular Weight Determination of WSAA
4.4. Design of Animal Experiment Program
4.5. Analysis of Physiological Indices
4.6. Blood Routine Examination
4.7. Lymphocytes Subsets Determination
4.8. Immune Cells Activities and IgG/IgM Expressions
4.9. Cytokines Levels Evaluation and Cell Cycle Detection
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Ballester, N.M.; Bataille, B.; Soulairol, I. Sodium alginate and alginic acid as pharmaceutical excipients for tablet formulation: Structure-function relationship. Carbohydr. Polym. 2021, 270, 118399. [Google Scholar] [CrossRef] [PubMed]
- Połomska, M.; Pogorzelec-Glaser, K.; Pawlaczyk, C.; Pietraszko, A. FT NIR Raman studies of alginic acid–benzimidazole polymer composite. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2011, 79, 797–800. [Google Scholar] [CrossRef] [PubMed]
- Boughrara, L.; Sebba, F.Z.; Sebti, H.; Choukchou-Braham, E.; Bounaceur, B.; Kada, S.O.; Zaoui, F. Removal of Zn(II) and Ni(II) heavy metal ions by new alginic acid-ester derivatives materials. Carbohydr. Polym. 2021, 272, 118439. [Google Scholar] [CrossRef] [PubMed]
- Ojerio, V.T.; Corre, V.L.; Toledo, N.A.; Andrino-Felarca, K.G.S.; Nievales, L.M.; Traifalgar, R.F.M. Alginic acid as immunostimulant: Effects of dose and frequency on growth performance, immune responses, and white spot syndrome virus resistance in tiger shrimp Penaeus monodon (Fabricius, 1798). Aquac. Int. 2018, 26, 267–278. [Google Scholar] [CrossRef]
- Wan, J.; Jiang, F.; Xu, Q.; Chen, D.; He, J. Alginic acid oligosaccharide accelerates weaned pig growth through regulating antioxidant capacity, immunity and intestinal development. RSC Adv. 2016, 6, 87026–87035. [Google Scholar] [CrossRef]
- Fernando, I.P.S.; Jayawardena, T.U.; Sanjeewa, K.K.A.; Wang, L.; Jeon, Y.-J.; Lee, W.W. Anti-inflammatory potential of alginic acid from Sargassum horneri against urban aerosol-induced inflammatory responses in keratinocytes and macrophages. Ecotoxicol. Environ. Saf. 2018, 160, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, Y.; Qin, Y.; Shen, P.; Peng, Q. Structures, properties and application of alginic acid: A review. Int. J. Biol. Macromol. 2020, 162, 618–628. [Google Scholar] [CrossRef]
- Kong, H.-J.; Lee, K.Y.; Mooney, D.J. Decoupling the dependence of rheological/mechanical properties of hydrogels from solids concentration. Polymer 2002, 43, 6239–6246. [Google Scholar] [CrossRef]
- Yu, R.; Petit, E.; Barboiu, M.; Li, S.; Sun, W.; Chen, C. Biobased dynamic hydrogels by reversible imine bonding for controlled release of thymopentin. Mater. Sci. Eng. C 2021, 127, 112210. [Google Scholar] [CrossRef]
- Schlesinger, D.H.; Goldstein, G. The amino acid sequence of thymopoietin II. Cell 1975, 5, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Bernengo, M.G.; Doveil, G.C.; Meregalli, M.; Appino, A.; Massobrio, R. Immunomodulation and Sézary syndrome: Experience with thymopentin (TP-5). Br. J. Dermatol. 1988, 119, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cheng, Y.; Zhang, X.; Zheng, L.; Han, Z.; Li, P.; Xiao, Y.; Zhang, Q.; Wang, F. The in vivo immunomodulatory and synergistic anti-tumor activity of thymosin α1–thymopentin fusion peptide and its binding to TLR2. Cancer Lett. 2013, 337, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Wang, W.; Wu, C.; Pan, Z.; Yao, G.; Fang, L.; Su, W. Myristic acid-modified thymopentin for enhanced plasma stability and immune-modulating activity. Int. Immunopharmacol. 2017, 47, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zhang, M.; Zhang, Z.; Wan, K.-W.; Ahmed, W.; Phoenix, D.A.; Elhissi, A.M.A.; Sun, X. Thymopentin Nanoparticles Engineered with High Loading Efficiency, Improved Pharmacokinetic Properties, and Enhanced Immunostimulating Effect Using Soybean Phospholipid and PHBHHx Polymer. Mol. Pharm. 2014, 11, 3371–3377. [Google Scholar] [CrossRef]
- Tischio, J.P.; Patrick, J.E.; Weintraub, H.S.; Chasin, M.; Goldstein, G. Short in vitro half-life of thymopoietin 32–36 pentapeptide in human plasma. Int. J. Pept. Protein Res. 1979, 14, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Qin, X.-Y.; Cao, X.; Li, W.-H.; Gong, T.; Zhang, Z.-R. Thymopentin-loaded phospholipid-based phase separation gel with long-lasting immunomodulatory effects: In vitro and in vivo studies. Acta Pharmacol. Sin. 2019, 40, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Chen, J.; Liu, L.; Zhang, J.; Wang, D.; Ma, L.; He, Y.; Liu, Y.; Liu, Z.; Wu, J. The X Protein of Hepatitis B Virus Inhibits Apoptosis in Hepatoma Cells through Enhancing the Methionine Adenosyltransferase 2A Gene Expression and Reducing S-Adenosylmethionine Production*. J. Biol. Chem. 2011, 286, 17168–17180. [Google Scholar] [CrossRef]
- Du, Y.; Kong, G.; You, X.; Zhang, S.; Zhang, T.; Gao, Y.; Ye, L.; Zhang, X. Elevation of Highly Up-regulated in Liver Cancer (HULC) by Hepatitis B Virus X Protein Promotes Hepatoma Cell Proliferation via Down-regulating p18*. J. Biol. Chem. 2012, 287, 26302–26311. [Google Scholar] [CrossRef]
- El–Serag, H.B.; Rudolph, K.L. Hepatocellular Carcinoma: Epidemiology and Molecular Carcinogenesis. Gastroenterology 2007, 132, 2557–2576. [Google Scholar] [CrossRef]
- Wang, J.; Liu, W.; Chen, Z.; Chen, H. Physicochemical characterization of the oolong tea polysaccharides with high molecular weight and their synergistic effects in combination with polyphenols on hepatocellular carcinoma. Biomed. Pharmacother. 2017, 90, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Xiong, C.; Xu, P.; Zhu, J.; Yang, Z.; Ren, H.; Luo, Q. Structural characterization and in vitro antitumor activity of A polysaccharide from Artemisia annua L. (Huang Huahao). Carbohydr. Polym. 2019, 213, 361–369. [Google Scholar] [CrossRef]
- Guo, H.; Zhang, T.; Yu, Y.; Xu, F. Cancer Physical Hallmarks as New Targets for Improved Immunotherapy. Trends Cell Biol. 2021, 31, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.D.; Cabral, H.; Stylianopoulos, T.; Jain, R.K. Improving cancer immunotherapy using nanomedicines: Progress, opportunities and challenges. Nat. Rev. Clin. Oncol. 2020, 17, 251–266. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, E.; Shiratsuchi, Y.; Kobayashi, Y.; Nagata, K. The importance of infiltrating neutrophils in SDF-1 production leading to regeneration of the thymus after whole-body X-irradiation. Cell Immunol. 2011, 268, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.-J.; Feng, Y.; Li, X.; Wang, X.-X.; Wang, Y.; Zhou, B.-T.; Liu, Y.; Liu, T.; Wu, Y.-C. Thymopentin (TP-5) prevents lipopolysaccharide-induced neuroinflammation and dopaminergic neuron injury by inhibiting the NF-κB/NLRP3 signaling pathway. Int. Immunopharmacol. 2023, 119, 110109. [Google Scholar] [CrossRef] [PubMed]
- Billerhart, M.; Schönhofer, M.; Schueffl, H.; Polzer, W.; Pichler, J.; Decker, S.; Taschauer, A.; Maier, J.; Anton, M.; Eckmann, S.; et al. CD47-targeted cancer immunogene therapy: Secreted SIRPα-Fc fusion protein eradicates tumors by macrophage and NK cell activation. Mol. Ther.-Oncolytics 2021, 23, 192–204. [Google Scholar] [CrossRef]
- Gungabeesoon, J.; Gort-Freitas, N.A.; Kiss, M.; Bolli, E.; Messemaker, M.; Siwicki, M.; Hicham, M.; Bill, R.; Koch, P.; Cianciaruso, C.; et al. A neutrophil response linked to tumor control in immunotherapy. Cell 2023, 186, 1448–1464.e1420. [Google Scholar] [CrossRef]
- Grządka, E.; Matusiak, J.; Godek, E. Alginic acid as a stabilizer of zirconia suspensions in the presence of cationic surfactants. Carbohydr. Polym. 2020, 246, 116634. [Google Scholar] [CrossRef]
- Fan, Y.-Z.; Chang, H.; Yu, Y.; Liu, J.; Zhao, L.; Yang, D.-J.; Wang, R. Thymopentin (TP5), an immunomodulatory peptide, suppresses proliferation and induces differentiation in HL-60 cells. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2006, 1763, 1059–1066. [Google Scholar] [CrossRef]
- Ji, H.Y.; Liu, C.; Dai, K.Y.; Yu, J.; Liu, A.J.; Chen, Y.F. The immunosuppressive effects of low molecular weight chitosan on thymopentin-activated mice bearing H22 solid tumors. Int. Immunopharmacol. 2021, 99, 108008. [Google Scholar] [CrossRef]
- Sadhukhan, R.; Majumdar, D.; Garg, S.; Landes, R.D.; McHargue, V.; Pawar, S.A.; Chowdhury, P.; Griffin, R.J.; Narayanasamy, G.; Boerma, M.; et al. Simultaneous exposure to chronic irradiation and simulated microgravity differentially alters immune cell phenotype in mouse thymus and spleen. Life Sci. Space Res. 2021, 28, 66–73. [Google Scholar] [CrossRef]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Ghosh, S. Intestinal barrier function and metabolic/liver diseases. Liver Res. 2020, 4, 81–87. [Google Scholar] [CrossRef]
- Gilreath, J.A.; Rodgers, G.M. How I treat cancer-associated anemia. Blood 2020, 136, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Granwehr, B.P.; Kontoyiannis, D.P. Fungal Infections in Cancer Patients. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Ding, N.; He, K.; Tian, H.; Li, L.; Li, Q.; Lu, S.; Ding, K.; Liu, J.; Nice, E.C.; Zhang, W.; et al. Carrier-free delivery of thymopentin-regulated injectable nanogels via an enhanced cancer immunity cycle against melanoma metastasis. Mater. Today Bio 2023, 20, 100645. [Google Scholar] [CrossRef] [PubMed]
- Horwood, N.J. Macrophage Polarization and Bone Formation: A review. Clin. Rev. Allergy Immunol. 2016, 51, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.M.; Bakan, C.E.; Mishra, A.; Hofmeister, C.C.; Efebera, Y.; Becknell, B.; Baiocchi, R.A.; Zhang, J.; Yu, J.; Smith, M.K.; et al. The PD-1/PD-L1 axis modulates the natural killer cell versus multiple myeloma effect: A therapeutic target for CT-011, a novel monoclonal anti–PD-1 antibody. Blood 2010, 116, 2286–2294. [Google Scholar] [CrossRef]
- Oh, D.Y.; Kwek, S.S.; Raju, S.S.; Li, T.; McCarthy, E.; Chow, E.; Aran, D.; Ilano, A.; Pai, C.-C.S.; Rancan, C.; et al. Intratumoral CD4+ T Cells Mediate Anti-tumor Cytotoxicity in Human Bladder Cancer. Cell 2020, 181, 1612–1625.e1613. [Google Scholar] [CrossRef]
- Shalapour, S.; Karin, M. The neglected brothers come of age: B cells and cancer. Semin. Immunol. 2021, 52, 101479. [Google Scholar] [CrossRef]
- Ostoa-Saloma, P. Chapter 6—The IgM as a tool for recognition of early tumoral antigens. In Immunotherapy in Resistant Cancer: From the Lab Bench Work to Its Clinical Perspectives; Morales-Montor, J., Segovia-Mendoza, M., Eds.; Academic Press: Cambridge, MA, USA, 2021; Volume 2, pp. 91–101. [Google Scholar] [CrossRef]
- Yang, S.; Cui, M.; Liu, Q.; Liao, Q. Glycosylation of immunoglobin G in tumors: Function, regulation and clinical implications. Cancer Lett. 2022, 549, 215902. [Google Scholar] [CrossRef]
- Varışlı, B.; Caglayan, C.; Kandemir, F.M.; Gür, C.; Ayna, A.; Genç, A.; Taysı, S. Chrysin mitigates diclofenac-induced hepatotoxicity by modulating oxidative stress, apoptosis, autophagy and endoplasmic reticulum stress in rats. Mol. Biol. Rep. 2023, 50, 433–442. [Google Scholar] [CrossRef]
- Liu, C.; Chu, D.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G. Cytokines: From Clinical Significance to Quantification. Adv. Sci. 2021, 8, 2004433. [Google Scholar] [CrossRef] [PubMed]
- Emre Kızıl, H.; Gür, C.; Ayna, A.; Darendelioğlu, E.; Küçükler, S.; Sağ, S. Contribution of Oxidative Stress, Apoptosis, Endoplasmic Reticulum Stress and Autophagy Pathways to the Ameliorative Effects of Hesperidin in NaF-Induced Testicular Toxicity. Chem. Biodivers. 2023, 20, e202200982. [Google Scholar] [CrossRef]
- Gui, L.; Zeng, Q.; Xu, Z.; Zhang, H.; Qin, S.; Liu, C.; Xu, C.; Qian, Z.; Zhang, S.; Huang, S.; et al. IL-2, IL-4, IFN-γ or TNF-α enhances BAFF-stimulated cell viability and survival by activating Erk1/2 and S6K1 pathways in neoplastic B-lymphoid cells. Cytokine 2016, 84, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Sahu, A.K.; Srivastava, A.; Chowdhury, R.; Mukherjee, S. Exploring the extensive crosstalk between the antagonistic cytokines-TGF-β and TNF-α in regulating cancer pathogenesis. Cytokine 2021, 138, 155348. [Google Scholar] [CrossRef] [PubMed]
- Todorović-Raković, N. The role of cytokines in the evolution of cancer: IFN-γ paradigm. Cytokine 2021, 151, 155442. [Google Scholar] [CrossRef] [PubMed]
- Mizui, M. Natural and modified IL-2 for the treatment of cancer and autoimmune diseases. Clin. Immunol. 2019, 206, 63–70. [Google Scholar] [CrossRef]
- Suzuki, A.; Leland, P.; Joshi, B.H.; Puri, R.K. Targeting of IL-4 and IL-13 receptors for cancer therapy. Cytokine 2015, 75, 79–88. [Google Scholar] [CrossRef]
- Ji, H.-Y.; Liu, C.; Dai, K.-Y.; Yu, J.; Liu, A.-J.; Chen, Y.-F. The extraction, structure, and immunomodulation activities in vivo of polysaccharides from Salvia miltiorrhiza. Ind. Crops Prod. 2021, 173, 114085. [Google Scholar] [CrossRef]
- Ji, H.-Y.; Yu, J.; Liu, A.-J. Structural characterization of a low molecular weight polysaccharide from Grifola frondosa and its antitumor activity in H22 tumor-bearing mice. J. Funct. Foods 2019, 61, 103472. [Google Scholar] [CrossRef]
- Yu, J.; Ji, H.-Y.; Liu, C.; Liu, A.-J. The structural characteristics of an acid-soluble polysaccharide from Grifola frondosa and its antitumor effects on H22-bearing mice. Int. J. Biol. Macromol. 2020, 158, 1288–1298. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Units | Blank Group | Model Group | TP5 Group | WSAAH Group | WSAAL+TP5 Group | WSAAH+TP5 Group |
|---|---|---|---|---|---|---|---|
| Leukocyte | 109/L | 5.63 ± 1.01 | 8.97 ± 1.52 α | 7.60 ± 1.36 β | 9.15 ± 1.46 | 6.53 ± 1.25 β | 6.30 ± 1.41 β |
| Lymphocyte proportion | % | 72.29 ± 4.62 | 53.73 ± 5.26 α | 64.74 ± 5.15 β | 52.79 ± 4.63 | 69.22 ± 4.85 β | 74.44 ± 6.10 β |
| Intermediate cell proportion | % | 2.49 ± 0.12 | 1.90 ± 0.13 α | 1.84 ± 0.16 | 1.64 ± 0.18 | 1.84 ± 0.13 | 2.54 ± 0.19 β |
| Granulocyte proportion | % | 25.22 ± 1.83 | 44.37 ± 3.69 α | 33.42 ± 3.10 β | 45.57 ± 2.96 | 28.94 ± 2.12 β | 23.02 ± 2.05 β |
| Erythrocyte | 1012/L | 9.56 ± 0.62 | 7.18 ± 0.46 α | 8.24 ± 0.49 | 8.59 ± 0.45 | 9.19 ± 0.68 β | 9.28 ± 0.67 β |
| Hemoglobin | g/L | 181.63 ± 9.68 | 158.70 ± 8.79 α | 165.79 ± 8.07 β | 159.49 ± 8.01 | 170.41 ± 10.67 β | 174.55 ± 10.09 β |
| Mean corpuscular hemoglobin concentration | g/L | 374.78 ± 20.95 | 351.40 ± 26.63 α | 365.80 ± 16.09 β | 354.52 ± 19.37 | 370.11 ± 20.78 β | 375.23 ± 23.65 β |
| Platelet | 109/L | 353.23 ± 16.98 | 421.67 ± 21.84 α | 388.94 ± 20.59 β | 419.53 ± 29.38 | 376.23 ± 26.49 β | 368.97 ± 20.91 β |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, H.; Fan, Y.; Gao, X.; Gong, Y.; Dai, K.; Wang, Z.; Xu, B.; Yu, J. The Protective Effects of Water-Soluble Alginic Acid on the N-Terminal of Thymopentin. Molecules 2023, 28, 6445. https://doi.org/10.3390/molecules28186445
Ji H, Fan Y, Gao X, Gong Y, Dai K, Wang Z, Xu B, Yu J. The Protective Effects of Water-Soluble Alginic Acid on the N-Terminal of Thymopentin. Molecules. 2023; 28(18):6445. https://doi.org/10.3390/molecules28186445
Chicago/Turabian StyleJi, Haiyu, Yuting Fan, Xiaoji Gao, Youshun Gong, Keyao Dai, Zhenhua Wang, Bo Xu, and Juan Yu. 2023. "The Protective Effects of Water-Soluble Alginic Acid on the N-Terminal of Thymopentin" Molecules 28, no. 18: 6445. https://doi.org/10.3390/molecules28186445
APA StyleJi, H., Fan, Y., Gao, X., Gong, Y., Dai, K., Wang, Z., Xu, B., & Yu, J. (2023). The Protective Effects of Water-Soluble Alginic Acid on the N-Terminal of Thymopentin. Molecules, 28(18), 6445. https://doi.org/10.3390/molecules28186445