
Exploring the Potential of Plant Bioactive Compounds against Male Infertility: An In Silico and In Vivo Study


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
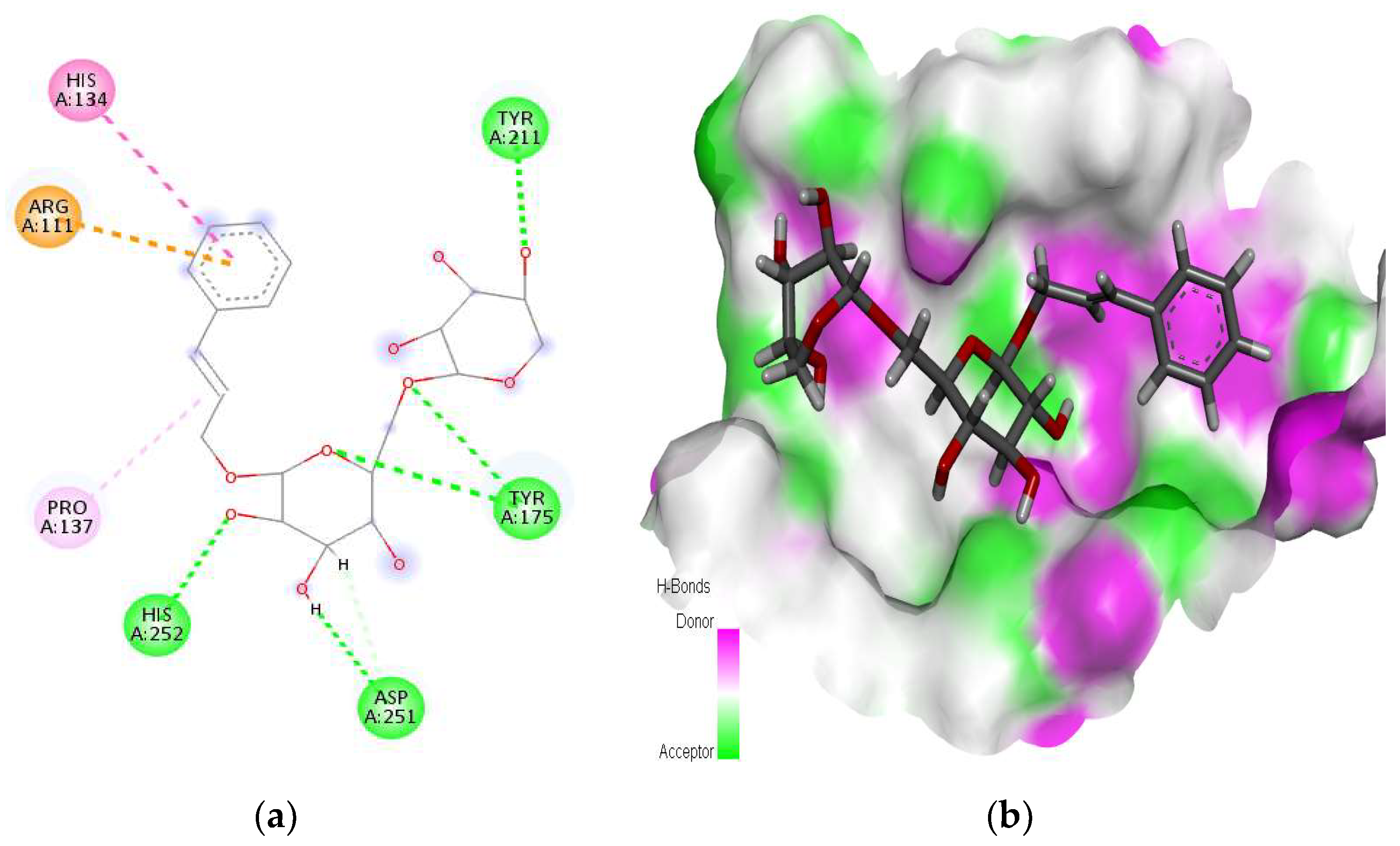
2.1. Interaction Analysis
2.2. Drug Scanning
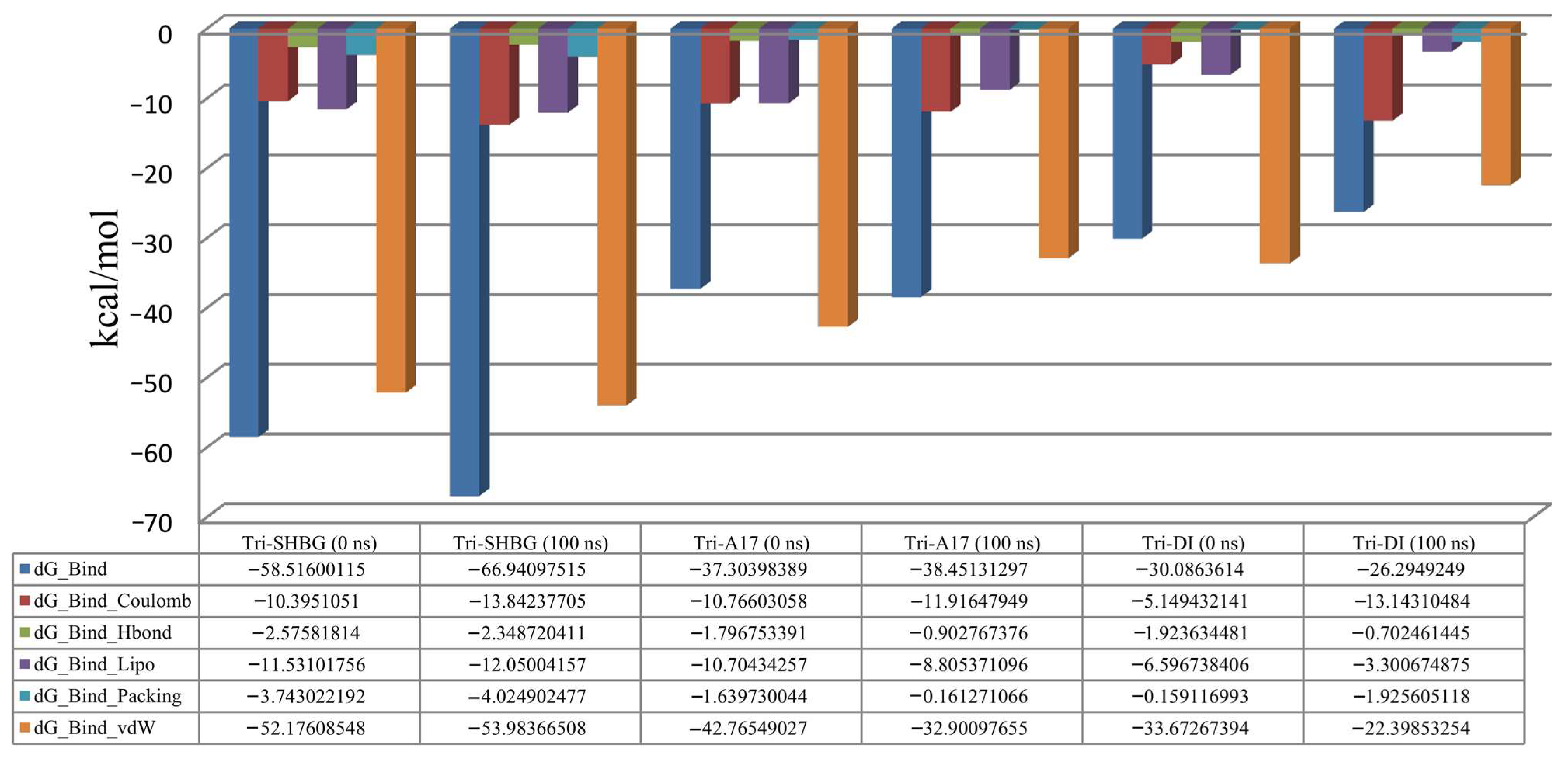
2.3. Molecular Dynamics Simulation Study
2.4. In Vivo Evaluation
2.4.1. Liver Function and Lipid Profile
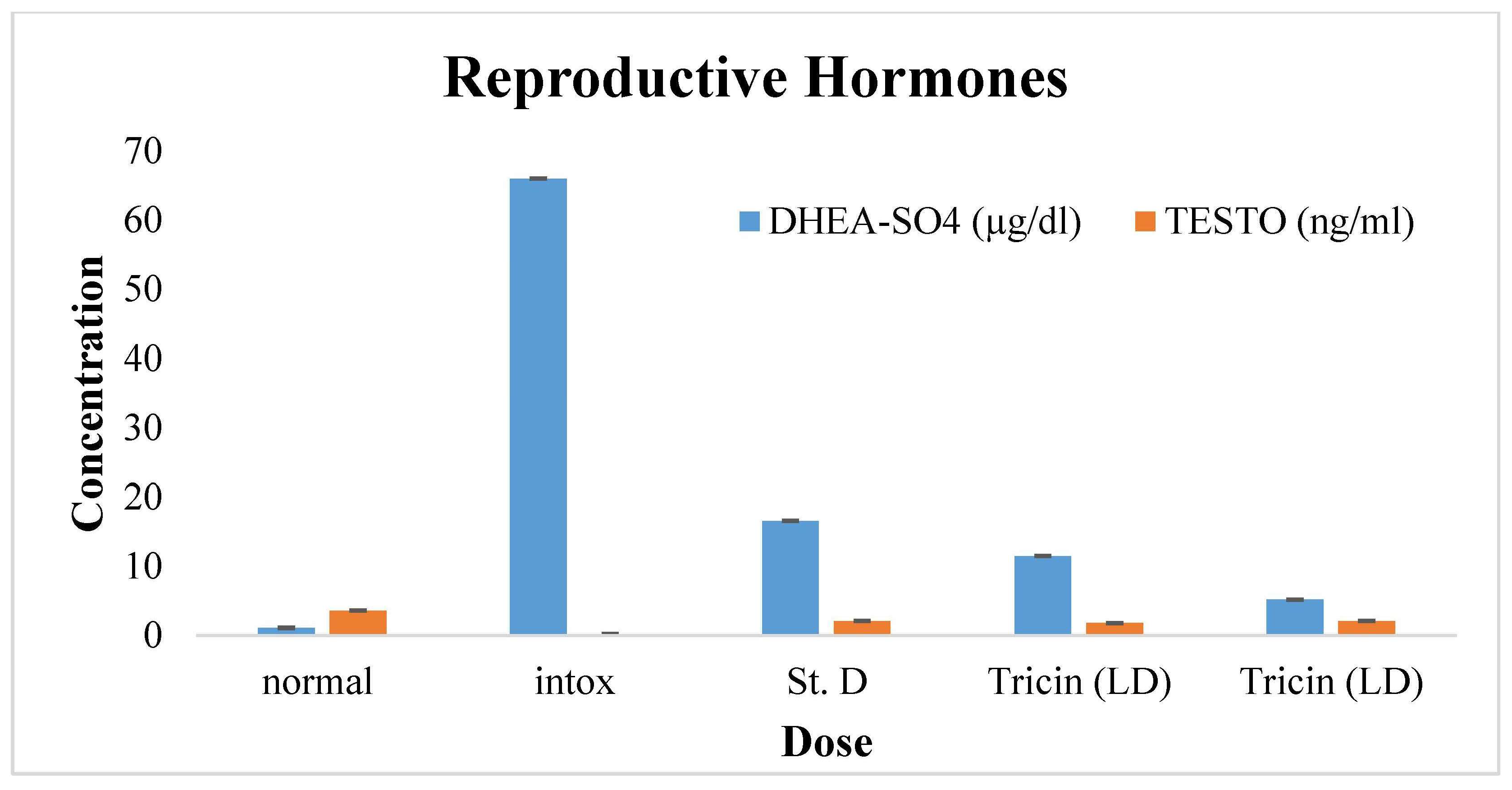
2.4.2. Reproductive Hormones
2.4.3. Histo-Architecture of Liver and Testicular Tissue
3. Discussion
4. Materials and Methods
4.1. Retrieval and Optimization of Phytochemicals
4.2. Retrieval and Preparation of Receptor Proteins
4.3. Molecular Docking Study
4.4. Druggability Analyses
4.5. Molecular Dynamics Simulation Study
4.6. In Vivo Evaluation
4.6.1. Grouping of Animals and Doses Plan
4.6.2. Biochemical Determination
4.6.3. Determination of Hormones
4.6.4. Histological Examination of Tissues
4.6.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Münevver, S. Erkek İnfertilite Nedenleri. Experimed 2020, 10, 144–147. [Google Scholar]
- Horta, F.; Vollenhoven, B.; Healey, M.; Busija, L.; Catt, S.; Temple-Smith, P. Male ageing is negatively associated with the chance of live birth in IVF/ICSI cycles for idiopathic infertility. Hum. Reprod. 2019, 34, 2523–2532. [Google Scholar] [CrossRef]
- Tüttelmann, F.; Ruckert, C.; Röpke, A. Disorders of spermatogenesis. Med. Genet. 2018, 30, 12–20. [Google Scholar] [CrossRef]
- Dong, Y.; Du, R.; Jiang, Y.; Wu, J.; Li, L.; Liu, R. Impact of chromosomal translocations on male infertility, semen quality, testicular volume and reproductive hormone levels. J. Int. Med. Res. 2012, 40, 2274–2283. [Google Scholar] [CrossRef] [PubMed]
- Selvam, M.K.P.; Agarwal, A. Update on the proteomics of male infertility: A systematic review. Arab J. Urol. 2018, 16, 103–112. [Google Scholar] [CrossRef]
- Cui, Y.-R.; Guo, Y.-H.; Qiao, S.-D.; Leng, L.-F.; Xie, Z.-H.; Chen, H.; Wang, X.-L. Correlation between SHBG gene polymorphism and male infertility in Han population of Henan province of China: A STROBE-compliant article. Medicine 2017, 96, e7753. [Google Scholar] [CrossRef]
- Wan, Q.; Xie, Y.; Zhou, Y.; Shen, X. Research progress on the relationship between sex hormone-binding globulin and male reproductive system diseases. Andrologia 2021, 53, e13893. [Google Scholar] [CrossRef]
- Mohamed, D.I.; Abou-Bakr, D.A.; Ezzat, S.F.; El-Kareem, H.F.A.; Nahas, H.H.A.; Saad, H.A.; Mehana, A.E.; Saied, E.M. Vitamin D3 prevents the deleterious effects of testicular torsion on testis by targeting miRNA-145 and ADAM17: In silico and in vivo study. Pharmaceuticals 2021, 14, 1222. [Google Scholar] [CrossRef] [PubMed]
- Ilić, B.S.; Kolarević, A.; Kocić, G.; Šmelcerović, A. Ascorbic acid as DNase I inhibitor in prevention of male infertility. Biochem. Biophys. Res. Commun. 2018, 498, 1073–1077. [Google Scholar] [CrossRef]
- Ertl, P.; Schuffenhauer, A. Estimation of synthetic accessibility score of drug-like molecules based on molecular complexity and fragment contributions. J. Cheminform. 2009, 1, 8. [Google Scholar] [CrossRef]
- Pastori, D.; Pani, A.; Di Rocco, A.; Menichelli, D.; Gazzaniga, G.; Farcomeni, A.; D’Erasmo, L.; Angelico, F.; Del Ben, M.; Baratta, F. Statin liver safety in non-alcoholic fatty liver disease: A systematic review and metanalysis. Br. J. Clin. Pharmacol. 2022, 88, 441–451. [Google Scholar] [CrossRef]
- Ramgir, S.S.; Renu, K.; Vellingiri, B.; George, A.; Tirupapuliyur, D.; Thiagarajan, P.; Valsala Gopalakrishnan, A. Phytomedicinal therapeutics for male infertility: Critical insights and scientific updates. J. Nat. Med. 2022, 76, 546–573. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zheng, Y.; Zeng, W.; Chen, L.; Yang, S.; Du, P.; Wang, Y.; Yu, X.; Zhang, X. Proteomic profile of sperm in infertile males reveals changes in metabolic pathways. Protein J. 2021, 40, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Panner Selvam, M.K.; Finelli, R.; Agarwal, A.; Henkel, R. Proteomics and metabolomics—Current and future perspectives in clinical andrology. Andrologia 2021, 53, e13711. [Google Scholar] [CrossRef] [PubMed]
- Esther, M.Y.J.; Subramaniyan, V.; Kumar, A.P.; Subramanian, M.; Palani, M. Molecular docking, ADMET analysis and dynamics approach to potent natural inhibitors against sex hormone binding globulin in male infertility. Pharmacogn. J. 2017, 9, s35–s43. [Google Scholar] [CrossRef]
- Schulte, R.T.; Ohl, D.A.; Sigman, M.; Smith, G.D. Sperm DNA damage in male infertility: Etiologies, assays, and outcomes. J. Assist. Reprod. Genet. 2010, 27, 3–12. [Google Scholar] [CrossRef]
- Taiwo, O.A.; Dosumu, O.A.; Oni, E.O.; Akomolafe, V.O.; Elazab, S.T.; Qusti, S.; Alshammari, E.M.; Batiha, G.E.S.; Ojo, O.A. Preclinical prediction of phytochemicals identified from cannabis as novel inhibitors of TEX 11, DHCR24, and CatSper 1 in fertility drug design. Inform. Med. Unlocked 2021, 26, 100747. [Google Scholar] [CrossRef]
- Nweze, C.C.; Mustapha, A.A.; Alkali, I.M. Aqueous leaf extracts of Tobacco plant (Nicotiana tabaccum) causes hepatotoxicity in male Wistar albino rats. Asian J. Pharmacol. Toxicol. 2015, 3, 27–30. [Google Scholar]
- Shakya, A.K.; Sharma, N.; Bhadauria, M.; Nirala, S.K.; Shrivastava, S.; Shukla, S. Ameliorative impact of herbal formulation—Majoon-Dabeed-ul-ward and Sharbat-e-Deenar against CCl4 induced liver toxicity via regulation of antioxidant enzymes and oxidative stress. Toxicol. Res. 2022, 11, 819–830. [Google Scholar] [CrossRef]
- Uemura, Y.; Shibata, R.; Takemoto, K.; Uchikawa, T.; Koyasu, M.; Watanabe, H.; Mitsuda, T.; Miura, A.; Imai, R.; Watarai, M. Elevation of red blood cell distribution width during hospitalization predicts mortality in patients with acute decompensated heart failure. J. Cardiol. 2016, 67, 268–273. [Google Scholar] [CrossRef]
- Bansal, S. Sexual dysfunction in hypertensive men. A critical review of the literature. Hypertension 1988, 12, 1–10. [Google Scholar] [CrossRef]
- Biswas, S.; Mita, M.A.; Afrose, S.; Hasan, M.R.; Islam, M.T.; Rahman, M.A.; Ara, M.J.; Chowdhury, M.B.A.; Meem, H.N.; Mamunuzzaman, M. Integrated Computational Approaches for Inhibiting Sex Hormone-Binding Globulin in Male Infertility by Screening Potent Phytochemicals. Life 2023, 13, 476. [Google Scholar] [CrossRef]
- Masuku, N.P.; Unuofin, J.O.; Lebelo, S.L. Phytochemical content, antioxidant activities and androgenic properties of four South African medicinal plants. J. Herbmed Pharmacol. 2020, 9, 245–256. [Google Scholar] [CrossRef]
- Usharani, P.; Merugu, P.L.; Nutalapati, C. Evaluation of the effects of a standardized aqueous extract of Phyllanthus emblica fruits on endothelial dysfunction, oxidative stress, systemic inflammation and lipid profile in subjects with metabolic syndrome: A randomised, double blind, placebo controlled clinical study. BMC Complement. Altern. Med. 2019, 19, 97. [Google Scholar]
- Rahmouni, F.; Saoudi, M.; Amri, N.; El-Feki, A.; Rebai, T.; Badraoui, R. Protective effect of Teucrium polium on carbon tetrachloride induced genotoxicity and oxidative stress in rats. Arch. Physiol. Biochem. 2018, 124, 1–9. [Google Scholar] [CrossRef]
- Salonia, A.; Corona, G.; Giwercman, A.; Maggi, M.; Minhas, S.; Nappi, R.E.; Sofikitis, N.; Vignozzi, L. SARS-CoV-2, testosterone and frailty in males (PROTEGGIMI): A multidimensional research project. Andrology 2021, 9, 19–22. [Google Scholar] [CrossRef]
- Munir, N.; Mahmood, Z.; Yameen, M.; Mustafa, G. Therapeutic response of Epimedium gandiflorum’s different doses to restore the antioxidant potential and reproductive hormones in male albino rats. Dose-Response 2020, 18, 1559325820959563. [Google Scholar] [CrossRef]
- Kubba, M.A. In vivo histological study the potential of Borago officinalis on spleen and testis histo-architecture on CCL4 induced albino male mice. Int. J. Res. Appl. Sci. Biotechnol. 2021, 8, 79–83. [Google Scholar] [CrossRef]
- Lam, P.Y.; Lui, A.C.; Wang, L.; Liu, H.; Umezawa, T.; Tobimatsu, Y.; Lo, C. Tricin biosynthesis and bioengineering. Front. Plant Sci. 2021, 12, 733198. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Pu, Y.; Yoo, C.G.; Ragauskas, A.J. The occurrence of tricin and its derivatives in plants. Green Chem. 2016, 18, 1439–1454. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-molecule library screening by docking with PyRx. Chem. Biol. Methods Protoc. 2015, 1263, 243–250. [Google Scholar]
- Burley, S.K.; Berman, H.M.; Kleywegt, G.J.; Markley, J.L.; Nakamura, H.; Velankar, S. Protein Data Bank (PDB): The single global macromolecular structure archive. Protein Crystallogr. Methods Protoc. 2017, 1607, 627–641. [Google Scholar]
- Vilar, S.; Cozza, G.; Moro, S. Medicinal chemistry and the molecular operating environment (MOE): Application of QSAR and molecular docking to drug discovery. Curr. Top. Med. Chem. 2008, 8, 1555–1572. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, G.; Mahrosh, H.S.; Arif, R. In silico characterization of growth differentiation factors as inhibitors of TNF-alpha and IL-6 in immune-mediated inflammatory disease rheumatoid arthritis. BioMed Res. Int. 2021, 2021, 5538535. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sharma, A.; Gupta, U. Molecular Docking studies on the Anti-fungal activity of Allium sativum (Garlic) against Mucormycosis (black fungus) by BIOVIA discovery studio visualizer 21.1. 0.0. Ann. Antivir. Antiretrovir. 2021, 5, 028–032. [Google Scholar]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Xiong, G.; Wu, Z.; Yi, J.; Fu, L.; Yang, Z.; Hsieh, C.; Yin, M.; Zeng, X.; Wu, C.; Lu, A. ADMETlab 2.0: An integrated online platform for accurate and comprehensive predictions of ADMET properties. Nucleic Acids Res. 2021, 49, W5–W14. [Google Scholar] [CrossRef]
- Pires, D.E.; Blundell, T.L.; Ascher, D.B. pkCSM: Predicting small-molecule pharmacokinetic and toxicity properties using graph-based signatures. J. Med. Chem. 2015, 58, 4066–4072. [Google Scholar] [CrossRef]
- Bowers, K.J.; Chow, E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D. Scalable algorithms for molecular dynamics simulations on commodity clusters. In Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006; p. 84-es. [Google Scholar]
- Azeem, M.; Mustafa, G.; Mahrosh, H.S. Virtual screening of phytochemicals by targeting multiple proteins of severe acute respiratory syndrome coronavirus 2: Molecular docking and molecular dynamics simulation studies. Int. J. Immunopathol. Pharmacol. 2022, 36, 03946320221142793. [Google Scholar] [CrossRef]
- Bergmeyer, H.; Herder, M.; Ref, R. International federation of clinical chemistry (IFCC). J. Clin. Chem. Clin. Biochem. 1986, 24, 497–510. [Google Scholar] [PubMed]
- Burtis, C.A.; Ashwood, E.R. Tietz Textbook of Clinical Chemistry; Amer Assn for Clinical Chemistry: Philadelphia, PA, USA, 1994. [Google Scholar]
- Ehsani, A.H.; Mortazavi, H.; Balighi, K.; Hosseini, M.S.; Azizpour, A.; Hejazi, S.P.; Goodarzi, A.; Darvari, S.B. Changes in body mass index and lipid profile in psoriatic patients after treatment with standard protocol of infliximab. Acta Medica Iran. 2016, 54, 570–575. [Google Scholar]
- Mäkelä, R.; Leinonen, A.; Suominen, T. Analysis of luteinizing hormone (LH): Validation of a commercial ELISA kit for LH analysis and quantification in doping control samples. Drug Test. Anal. 2020, 12, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Suvarna, S.K.; Layton, C.; Bancroft, J.D. Theory and Practice of Histological Techniqueseighth; Elsevier Health Science: Oxford, UK, 2019. [Google Scholar]
- Montgomery, D.C. Design and Analysis of Experiments; John Wiley & Sons: Hoboken, NJ, USA, 2017. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | PubChem ID | Phytochemical | Receptor | Docking Score (kcal/mol) | Interacting Residues |
|---|---|---|---|---|---|
| 1 | 5281702 | Tricin | SHBG (6PYF) | −11.09 | Ser42, Phe56, Asp65, Phe67, Leu80, Asn82, Met107, Val127, Leu131, Lys134, Ile141, Leu171 |
| 2 | 5281672 | Myricetin | −10.72 | Asn61, Asp64, Asp65, Gly129, Lys134, Arg135 | |
| 3 | 5280343 | Quercetin | −9.70 | Asp64, Asp65, His83, Trp84, Ser133, Lys134, Arg135 | |
| 4 | 159287 | Malvidin | −9.65 | Thr60, Asp64, Asn82, His83, Trp84, Ser133, Lys134, Arg135 | |
| 5 | 5281691 | Rhamnetin | −8.07 | Asn61, Asp64, His83, Gly129, Ser133, Lys134, Arg135 | |
| 6 | 5281702 | Tricin | ADAM17 (2I47) | −13.15 | Leu348, Gly349, Leu401, Val402, His405, His415, Val434, Pro437, Ala439, Val440 |
| 7 | 44256621 | Callistephin | −10.68 | Leu348, Leu401, Val402, His405, Glu406, His409, Pro437, Ala439, Val440 | |
| 8 | 370 | Gallic acid | −9.21 | Glu398, Val402, His405, Val434, Ala439, Val440 | |
| 9 | 9823887 | Rosavin | −9.14 | Gly349, Met345, Leu348, Tyr390, Leu401, Val402, His405, Glu406, Ala439, Val440 | |
| 10 | 489941 | Moronic acid | −9.07 | Met345, Leu350, His405, His409, His415, Ile438, Ala439 | |
| 11 | 9823887 | Rosavin | DNase 1 | −9.46 | Arg111, His134, Pro137, Tyr175, Tyr211, Asp251, His252 |
| 12 | 5281702 | Tricin | −9.23 | Glu39, Tyr6, Arg111, His252 | |
| 13 | 5281417 | Esculin | −8.55 | His134, Asn170, Tyr175, Tyr211, Asp251 | |
| 14 | 5281294 | Okanin | −8.00 | Glu39, Arg41, His134, Asn170, Asp251, His252 | |
| 15 | 442428 | Naringin | −7.82 | Arg111, His134, Pro137, Asn170, Ser174, Tyr175, Asp251 |
| Sr. | Ligands | Target | Molecular Weight (<500 Dalton) | HBD (≤5) | HBA (≤10) | LogP (≤5) | Molar Refractivity (40–130) | Violations |
|---|---|---|---|---|---|---|---|---|
| 1 | Tricin | SHBG ADAM17 DNaseI | 330.29 | 3 | 7 | 2.15 | 86.97 | 0 |
| 2 | Myricetin | SHBG | 318.24 | 6 | 8 | 0.79 | 80.06 | 1 |
| 3 | Quercetin | 302.24 | 5 | 7 | 1.23 | 78.03 | 0 | |
| 4 | Malvidin | 331.30 | 4 | 7 | 0.92 | 87.13 | 0 | |
| 5 | Rhamnetin | 316.26 | 4 | 7 | 1.63 | 82.50 | 0 | |
| 6 | Isorhamnetin | 316.26 | 4 | 7 | 1.65 | 82.50 | 0 | |
| 7 | Callistephin | ADAM17 | 468.84 | 7 | 10 | −1.71 | 112.12 | 1 |
| 8 | Gallic acid | 170.12 | 4 | 5 | 0.21 | 40.47 | 0 | |
| 9 | Kaempferol | 286.24 | 4 | 6 | 1.58 | 76.01 | 0 | |
| 10 | Moronic acid | 454.7 | 1 | 3 | 6.03 | 135.69 | 2 | |
| 11 | Rosavin | ADAM17 DNaseI | 428.4 | 6 | 10 | −1.07 | 101.30 | 1 |
| 12 | Esculin | DNaseI | 340.28 | 5 | 9 | −0.62 | 78.65 | 0 |
| 13 | Robinetin | 302.24 | 5 | 7 | 1.12 | 78.03 | 0 | |
| 14 | Okanin | 288.25 | 5 | 6 | 1.69 | 76.36 | 0 | |
| 15 | Naringin | 580.53 | 8 | 14 | −0.87 | 134.91 | 4 |
| Category | Property | Myricetin | Malvidin | Rhamnetin | Quercetin | Isorhamnetin | Okanin | Callistephin |
|---|---|---|---|---|---|---|---|---|
| Absorption | Caco-2 > −5.15 | −5.653 | −5.159 | −5.109 | −5.204 | −5.056 | −5.375 | −6.196 |
| Pgp-Substrate | −−− | ++ | −−− | −−− | −−− | −−− | + | |
| Pgp-Inhibitor | −−− | −−− | −−− | −−− | −−− | −−− | −−− | |
| HIA | −−− | −−− | −−− | −−− | −−− | −−− | ++ | |
| Distribution | BBB | −−− | −−− | −−− | −−− | −−− | −−− | −− |
| PPB | 92.77% | 93.13% | 96.30% | 95.45% | 96.24% | 99.08% | 89.62% | |
| Metabolism | CYP1A2-Inhibitor | ++ | ++ | +++ | +++ | +++ | + | −−− |
| CYP1A2 substrate | −− | ++ | + | −− | ++ | −−− | −−− | |
| CYP2C19 inhibitor | −−− | −−− | −−− | −−− | −− | −−− | −−− | |
| CYP2C19 substrate | −−− | −−− | −−− | −−− | −−− | −−− | −−− | |
| CYP2C9 inhibitor | + | −−− | + | + | + | + | −−− | |
| CYP2C9 substrate | − | ++ | ++ | + | ++ | − | − | |
| CYP2D6 inhibitor | −−− | −−− | − | − | + | −−− | −−− | |
| CYP2D6 substrate | −− | + | − | −− | − | −− | −− | |
| CYP3A4 inhibitor | −− | −− | −− | − | + | −− | −−− | |
| CYP3A4 substrate | −−− | −− | −−− | −− | −−− | −− | −−− | |
| Excretion | Clearance | 7.716 | 11.093 | 8.284 | 6.991 | 16.361 | 7.971 | |
| Toxicity | AMES | − | −− | + | + | + | ++ | ++ |
| hERG | −− | −− | −−− | −−− | −−− | −−− | −−− | |
| FDAMDD | + | +++ | − | −−− | −−− | − | −−− | |
| H-HT | −−− | −−− | −−− | −−− | −−− | −− | −− | |
| Category | Property | Kaempferol | Gallic Acid | Rosavin | Esculin | Robinetin | Tricin | |
| Absorption | Caco-2 > −5.15 | −4.974 | −5.728 | −5.406 | −5.950 | −5.325 | −4.970 | |
| Pgp-Substrate | −−− | −−− | − | − | −−− | +++ | ||
| Pgp-Inhibitor | −−− | −−− | −−− | −−− | −−− | −−− | ||
| HIA | −−− | −−− | +++ | ++ | −−− | −− | ||
| Distribution | BBB | −−− | −−− | − | ++ | −−− | −−− | |
| PPB | 97.86% | 53.49% | 43.91% | 56.10% | 94.64% | 91.48% | ||
| Metabolism | CYP1A2-Inhibitor | +++ | −−− | −−− | −−− | ++ | +++ | |
| CYP1A2 substrate | −− | −−− | −−− | −−− | −− | +++ | ||
| CYP2C19 inhibitor | −− | −−− | −−− | −−− | −−− | −− | ||
| CYP2C19 substrate | −−− | −−− | −−− | −−− | −−− | −−− | ||
| CYP2C9 inhibitor | + | −− | −−− | −−− | + | + | ||
| CYP2C9 substrate | ++ | −−− | −− | − | − | ++ | ||
| CYP2D6 inhibitor | ++ | −−− | −−− | −−− | −− | + | ||
| CYP2D6 substrate | −− | −− | −− | −− | −− | ++ | ||
| CYP3A4 inhibitor | + | −−− | −−− | −−− | − | + | ||
| CYP3A4 substrate | −−− | −−− | −−− | −−− | −−− | −− | ||
| Excretion | Clearance | 6.868 | 10.108 | 1.218 | 4.015 | 7.773 | 6.626 | |
| Toxicity | AMES | + | −−− | −− | −− | + | − | |
| hERG | −−− | −−− | −−− | −−− | −−− | −− | ||
| FDAMDD | −− | −−− | −−− | −−− | − | ++ | ||
| H-HT | −−− | −− | −− | −− | −− | −−− | ||
| Control | Intoxicated | Positive Control | Tricin (LD) | Tricin (HD) | |
|---|---|---|---|---|---|
| ALT (IU/L) | 46.2 ± 4.11 D | 113.8 ± 5.38 A | 68 ± 4.90 C | 89.5 ± 5.32 B | 35 ± 3.27 E |
| AST (IU/L) | 51.8 ± 4.11 CD | 125.8 ± 6.55 A | 55.8 ± 3.30 C | 72 ± 5.72 B | 26 ± 3.27 E |
| Urea (mg/dL) | 22 ± 1.63 C | 54 ± 4.90 A | 24 ± 2.45 B | 19 ± 0.82 D | 17 ± 0.82 DE |
| Creatinine (mg/dL) | 0.8 ± 0.13 BC | 1.3 ± 0.08 A | 0.7 ± 0.16 BC | 0.7 ± 0.16 BC | 0.8 ± 0.13 BC |
| Cholesterol (mg/dL) | 152 ± 5.72 DE | 198 ± 9.80 A | 160.8 ± 7.37 CD | 178 ± 8.16 B | 138 ± 4.90 F |
| TG (mg/dL) | 173 ± 9.80 B | 221 ± 16.33 A | 122 ± 8.16 EF | 148 ± 7.35 CD | 137 ± 5.32 DE |
| HDL (mg/dL) | 38 ± 1.63 CDE | 35 ± 2.45 F | 45 ± 2.45 A | 41 ± 2.45 ABCD | 42 ± 2.45 ABC |
| LDL (mg/dL) | 80 ± 2.45 E | 87 ± 2.45 A | 85 ± 4.08 B | 84 ± 3.27 CD | 78 ± 2.53 F |
| Control | Intoxicated | Positive Control | Tricin (LD) | Tricin (HD) | |
|---|---|---|---|---|---|
| TESTO (ng/mL) | 3.6 ± 0.02 A | 0.2 ± 0.01D | 2.1 ± 0.08 B | 1.8 ± 0.01 C | 2.1 ± 0.01 B |
| LH (IU/L) | 0.8 ± 0.02 A | 0.2 ± 0.01 E | 0.6 ± 0.04 B | 0.4 ± 0.01 CD | 0.5 ± 0.02 C |
| FSH (IU/L) | 0.8 ± 0.04 A | 0.3 ± 0.04 E | 0.7 ± 0.01 B | 0.3 ± 0.01 DE | 0.53 ± 0.02 C |
| PROL (ng/mL) | 0.2 ± 0.04 C | 0.1 ± 0.01 D | 0.3 ± 0.02 B | 0.3 ± 0.02 B | 0.5 ± 0.02 A |
| DHEA-SO4 (µg/dL) | 1.1 ± 0.10 E | 66.0 ± 3.27 A | 16.6 ± 0.82 B | 11.5 ± 0.82 C | 5.2 ± 0.01 D |
| Group Number | Architecture Intact/Distorted | Necrosis | Periportal Inflammation Acute/Chronic | Congestion | Edema | Apoptosis | Hemorrhage |
|---|---|---|---|---|---|---|---|
| Control (G-1) | Intact | Not Seen | Not Seen | Not Seen | Not Seen | Not Seen | Not Seen |
| Intoxicated (G-2) | Intact | Seen | Moderate, chronic++ | Not seen | Seen | Seen | Not seen |
| Standard drug (G-3) | Intact | Not seen | Mild, chronic+ | Not seen | Seen | Seen | Not seen |
| Test group (G-4) | Intact | Not seen | Mild, chronic+ | Not seen | Not seen | Seen | Not seen |
| Group Name | Capsule | Seminiferous Tubules | Sertoli Cells | Germinal Cell Layer | Spermatogenesis | Tubular Necrosis | Edema | Inflammation |
|---|---|---|---|---|---|---|---|---|
| Control (G-1) | Intact | Intact | Seen | Seen | Seen | Not seen | Not seen | Not seen |
| Intoxicated (G-2) | Intact | Altered | Seen | Cellular depletion | Not seen | Seen | Seen | Chronic++ |
| Standard drug (G-3) | Intact | Intact | Seen | Cellular depletion | Diminished | Not seen | Not seen | Mild+ |
| Test group (G-4) | Intact | Intact | Seen | Seen | Seen | Not seen | Not seen | Not seen |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jahangeer, M.; Mustafa, G.; Munir, N.; Ahmed, S.; Al-Anazi, K.M. Exploring the Potential of Plant Bioactive Compounds against Male Infertility: An In Silico and In Vivo Study. Molecules 2023, 28, 7693. https://doi.org/10.3390/molecules28237693
Jahangeer M, Mustafa G, Munir N, Ahmed S, Al-Anazi KM. Exploring the Potential of Plant Bioactive Compounds against Male Infertility: An In Silico and In Vivo Study. Molecules. 2023; 28(23):7693. https://doi.org/10.3390/molecules28237693
Chicago/Turabian StyleJahangeer, Muhammad, Ghulam Mustafa, Naveed Munir, Sibtain Ahmed, and Khalid Mashai Al-Anazi. 2023. "Exploring the Potential of Plant Bioactive Compounds against Male Infertility: An In Silico and In Vivo Study" Molecules 28, no. 23: 7693. https://doi.org/10.3390/molecules28237693
APA StyleJahangeer, M., Mustafa, G., Munir, N., Ahmed, S., & Al-Anazi, K. M. (2023). Exploring the Potential of Plant Bioactive Compounds against Male Infertility: An In Silico and In Vivo Study. Molecules, 28(23), 7693. https://doi.org/10.3390/molecules28237693