
Chemical Composition, Antioxidant, and Antibiofilm Properties of Essential Oil from Thymus capitatus Plants Organically Cultured on the Greek Island of Lemnos

 ,
,  ,
, 
Abstract
:
1. Introduction
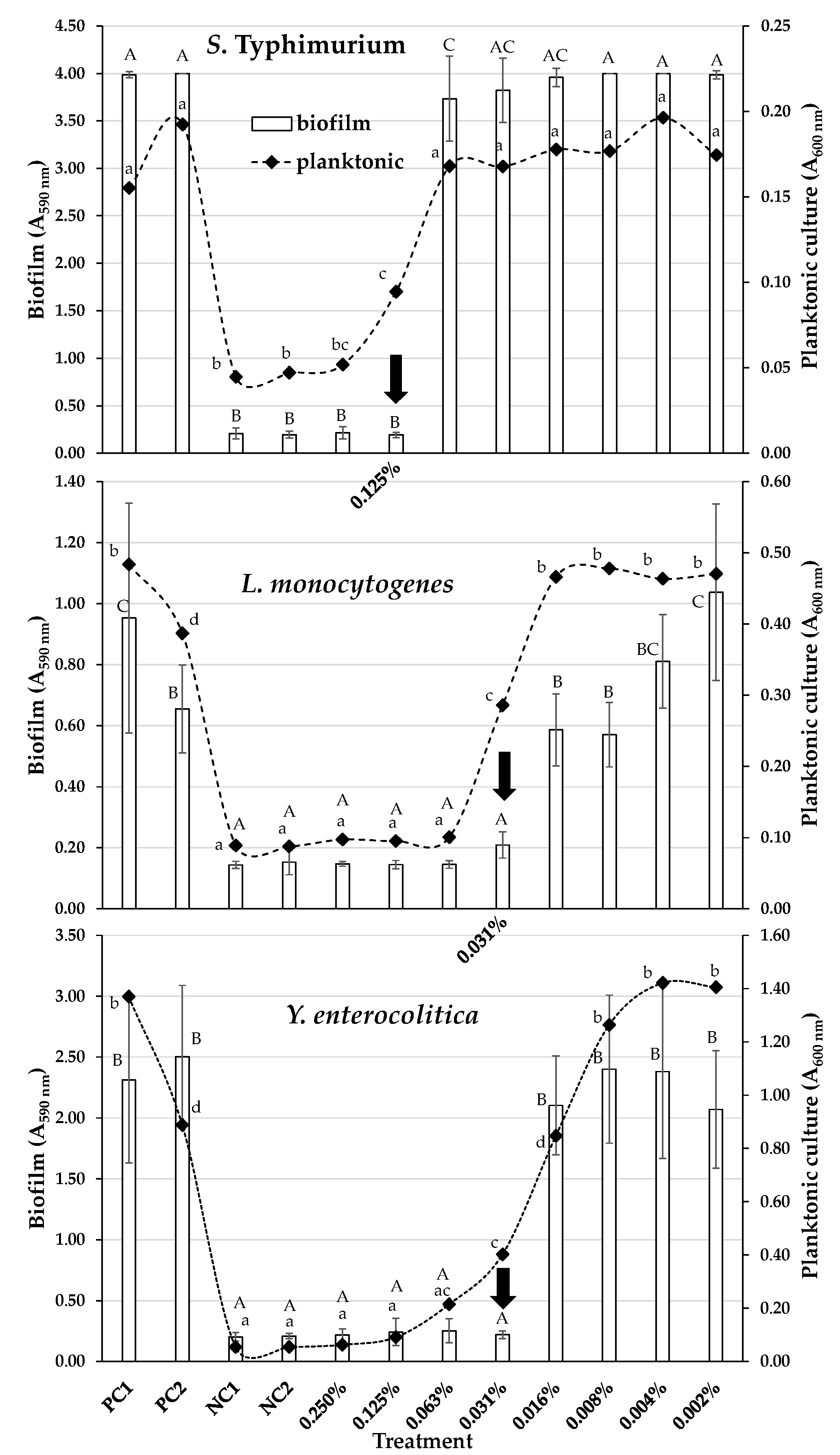
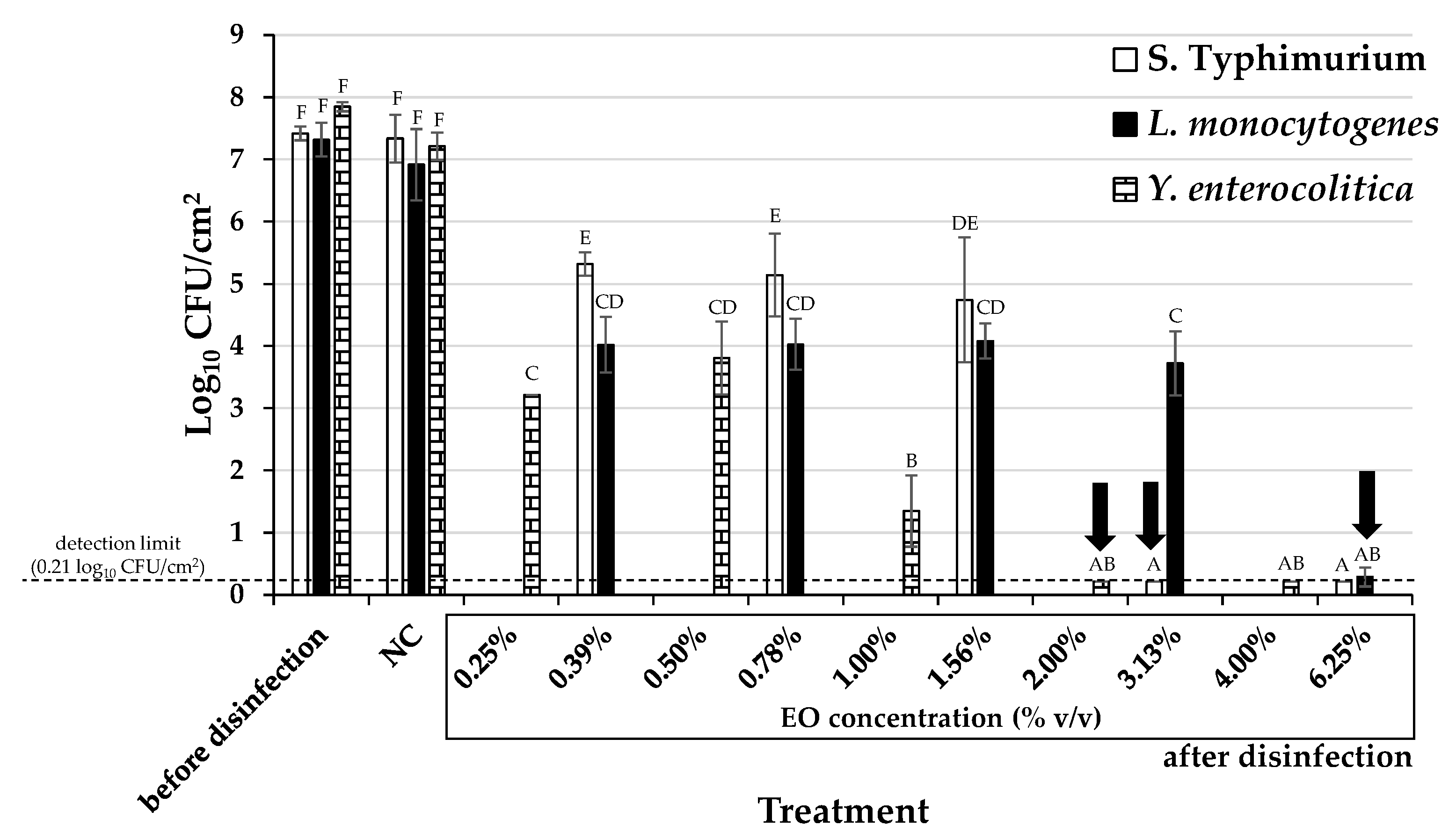
2. Results and Discussion
3. Materials and Methods
3.1. Chemicals
3.2. Hydrodistillation and Receipt of EO
3.3. Bacterial Strains and Preparation of Their Working Cultures
3.4. Determination of Minimum Inhibitory and Bactericidal Concentrations of EO against Planktonic Bacteria (MICs, MBCs)
3.5. Determination of Minimum Biofilm Inhibitory Concentrations (MBICs)
3.6. Determination of Minimum Biofilm Eradication Concentrations (MBECs)
3.7. Determination of the Antioxidant Activity (AA) of the EO
3.7.1. DPPH• Assay
3.7.2. Cupric Ion Reducing Antioxidant Capacity (CUPRAC) Assay
3.8. Determination of the Total Phenolic Content (TPC) of the EO
3.9. Chemical Analysis of EO (GC)
3.9.1. GC-FID
3.9.2. GC-MS
3.10. Statistics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- McEwen, S.A.; Collignon, P.J. Antimicrobial resistance: A One Health perspective. Microbiol. Spectr. 2018, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- One Health High-Level Expert Panel (OHHLEP). One Health: A new definition for a sustainable and healthy future. PLoS Pathog. 2022, 18, e1010537. [Google Scholar] [CrossRef]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial resistance in bacteria: Mechanisms, evolution, and persistence. J. Mol. Evol. 2020, 88, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Florez-Cuadrado, D.; Moreno, M.A.; Ugarte-Ruíz, M.; Domínguez, L. Antimicrobial resistance in the food chain in the European Union. Adv. Food Nutr. Res. 2018, 86, 115–136. [Google Scholar] [CrossRef] [PubMed]
- Antunes, P.; Novais, C.; Peixe, L. Food-to-humans bacterial transmission. Microbiol. Spectr. 2020, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odeyemi, O.A.; Alegbeleye, O.O.; Strateva, M.; Stratev, D. Understanding spoilage microbial community and spoilage mechanisms in foods of animal origin. Compr. Rev. Food Sci. Food Saf. 2020, 19, 311–331. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiology Reference Group 2007–2015; World Health Organization: Geneva, Switzerland, 2015; Available online: http://apps.who.int/iris/bitstream/10665/199350/1/9789241565165_eng.pdf (accessed on 12 December 2022).
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States—Major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Chlebicz, A.; Śliżewska, K. Campylobacteriosis, salmonellosis, yersiniosis, and listeriosis as zoonotic foodborne diseases: A review. Int. J. Environ. Res. Public Health 2018, 15, 863. [Google Scholar] [CrossRef] [Green Version]
- Fàbrega, A.; Vila, J. Yersinia enterocolitica: Pathogenesis, virulence and antimicrobial resistance. Enferm. Infecc. Microbiol. Clin. 2012, 30, 24–32. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union One Health 2020 zoonoses report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Lamas, A.; Regal, P.; Vázquez, B.; Miranda, J.M.; Cepeda, A.; Franco, C.M. Salmonella and Campylobacter biofilm formation: A comparative assessment from farm to fork. J. Sci. Food Agric. 2018, 98, 4014–4032. [Google Scholar] [CrossRef]
- Lenchenko, E.; Lozovoy, D.; Strizhakov, A.; Vatnikov, Y.; Byakhova, V.; Kulikov, E.; Sturov, N.; Kuznetsov, V.; Avdotin, V.; Grishin, V. Features of formation of Yersinia enterocolitica biofilms. Vet. World 2019, 12, 136–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-López, P.; Rodríguez-Herrera, J.J.; Vázquez-Sánchez, D.; López Cabo, M. Current knowledge on Listeria monocytogenes biofilms in food-related environments: Incidence, resistance to biocides, ecology and biocontrol. Foods 2018, 7, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.R.; Tay, M.; Palmer, J.; Flint, S. Biofilm formation of Yersinia enterocolitica and its persistence following treatment with different sanitation agents. Food Control 2017, 73, 433–437. [Google Scholar] [CrossRef]
- Bridier, A.; Sanchez-Vizuete, P.; Guilbaud, M.; Piard, J.C.; Naïtali, M.; Briandet, R. Biofilm-associated persistence of food-borne pathogens. Food Microbiol. 2015, 45(Pt B), 167–178. [Google Scholar] [CrossRef]
- Yan, J.; Bassler, B.L. Surviving as a community: Antibiotic tolerance and persistence in bacterial biofilms. Cell Host Microbe 2019, 26, 15–21. [Google Scholar] [CrossRef]
- Ciofu, O.; Moser, C.; Jensen, P.Ø.; Høiby, N. Tolerance and resistance of microbial biofilms. Nat. Rev. Microbiol. 2022, 20, 621–635. [Google Scholar] [CrossRef]
- Hall, C.W.; Mah, T.F. Molecular mechanisms of biofilm-based antibiotic resistance and tolerance in pathogenic bacteria. FEMS Microbiol. Rev. 2017, 41, 276–301. [Google Scholar] [CrossRef] [Green Version]
- Romeu, M.J.; Rodrigues, D.; Azeredo, J. Effect of sub-lethal chemical disinfection on the biofilm forming ability, resistance to antibiotics and expression of virulence genes of Salmonella Enteritidis biofilm-surviving cells. Biofouling 2020, 36, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Nadar, S.; Khan, T.; Patching, S.G.; Omri, A. Development of antibiofilm therapeutics strategies to overcome antimicrobial drug resistance. Microorganisms 2022, 10, 303. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef] [Green Version]
- Anghel, I.; Grumezescu, A.M.; Holban, A.M.; Ficai, A.; Anghel, A.G.; Chifiriuc, M.C. Biohybrid nanostructured iron oxide nanoparticles and Satureja hortensis to prevent fungal biofilm development. Int. J. Mol. Sci. 2013, 14, 18110–18123. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, R.; Rolta, R.; Dev, K.; Sourirajan, A. Synergistic potential of essential oils with antibiotics to combat fungal pathogens: Present status and future perspectives. Phytother. Res. 2021, 35, 6089–6100. [Google Scholar] [CrossRef]
- Borges, A.; Abreu, A.C.; Dias, C.; Saavedra, M.J.; Borges, F.; Simões, M. New perspectives on the use of phytochemicals as an emergent strategy to control bacterial infections including biofilms. Molecules 2016, 21, 877. [Google Scholar] [CrossRef] [PubMed]
- Ni, P.; Wang, L.; Deng, B.; Jiu, S.; Ma, C.; Zhang, C.; Almeida, A.; Wang, D.; Xu, W.; Wang, S. Combined application of bacteriophages and carvacrol in the control of Pseudomonas syringae pv. actinidiae planktonic and biofilm forms. Microorganisms 2020, 8, 837. [Google Scholar] [CrossRef] [PubMed]
- Sakarikou, C.; Kostoglou, D.; Simões, M.; Giaouris, E. Exploitation of plant extracts and phytochemicals against resistant Salmonella spp. in biofilms. Food Res. Int. 2020, 128, 108806. [Google Scholar] [CrossRef]
- Sakkas, H.; Papadopoulou, C. Antimicrobial activity of basil, oregano, and thyme essential oils. J. Microbiol. Biotechnol. 2017, 27, 429–438. [Google Scholar] [CrossRef] [Green Version]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential oils as antimicrobial agents-myth or real alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef] [Green Version]
- Rossi, C.; Chaves-López, C.; Serio, A.; Casaccia, M.; Maggio, F.; Paparella, A. Effectiveness and mechanisms of essential oils for biofilm control on food-contact surfaces: An updated review. Crit. Rev. Food Sci. Nutr. 2022, 62, 2172–2191. [Google Scholar] [CrossRef]
- Zhang, D.; Gan, R.Y.; Zhang, J.R.; Farha, A.K.; Li, H.B.; Zhu, F.; Wang, X.H.; Corke, H. Antivirulence properties and related mechanisms of spice essential oils: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1018–1055. [Google Scholar] [CrossRef]
- Hammoudi Halat, D.; Krayem, M.; Khaled, S.; Younes, S. A focused insight into thyme: Biological, chemical, and therapeutic properties of an indigenous Mediterranean herb. Nutrients 2022, 14, 2104. [Google Scholar] [CrossRef] [PubMed]
- Pl@ntNet™. Available online: https://identify.plantnet.org/medor/species/Coridothymus%20capitatus%20(L.)%20Rchb.f./data (accessed on 12 December 2022).
- Bouyahya, A.; Chamkhi, I.; Guaouguaou, F.E.; Benali, T.; Balahbib, A.; El Omari, N.; Taha, D.; El-Shazly, M.; El Menyiy, N. Ethnomedicinal use, phytochemistry, pharmacology, and food benefits of Thymus capitatus. J. Ethnopharmacol. 2020, 259, 112925. [Google Scholar] [CrossRef]
- Reganold, J.P.; Wachter, J.M. Organic agriculture in the twenty-first century. Nat. Plants 2016, 2, 15221. [Google Scholar] [CrossRef] [PubMed]
- Alabdullatif, M.; Boujezza, I.; Mekni, M.; Taha, M.; Kumaran, D.; Yi, Q.L.; Landoulsi, A.; Ramirez-Arcos, S. Enhancing blood donor skin disinfection using natural oils. Transfusion 2017, 57, 2920–2927. [Google Scholar] [CrossRef] [PubMed]
- Almeida, L.; Lopes, N.; Gaio, V.; Cavaleiro, C.; Salgueiro, L.; Silva, V.; Poeta, P.; Cerca, N. Thymbra capitata essential oil has a significant antimicrobial activity against methicillin-resistant Staphylococcus aureus pre-formed biofilms. Lett. Appl. Microbiol. 2022, 74, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Kapustová, M.; Puškárová, A.; Bučková, M.; Granata, G.; Napoli, E.; Annušová, A.; Mesárošová, M.; Kozics, K.; Pangallo, D.; Geraci, C. Biofilm inhibition by biocompatible poly(ε-caprolactone) nanocapsules loaded with essential oils and their cyto/genotoxicity to human keratinocyte cell line. Int. J. Pharm. 2021, 606, 120846. [Google Scholar] [CrossRef] [PubMed]
- Machado, D.; Gaspar, C.; Palmeira-de-Oliveira, A.; Cavaleiro, C.; Salgueiro, L.; Martinez-de-Oliveira, J.; Cerca, N. Thymbra capitata essential oil as potential therapeutic agent against Gardnerella vaginalis biofilm-related infections. Future Microbiol. 2017, 12, 407–416. [Google Scholar] [CrossRef]
- Palmeira-de-Oliveira, A.; Gaspar, C.; Palmeira-de-Oliveira, R.; Silva-Dias, A.; Salgueiro, L.; Cavaleiro, C.; Pina-Vaz, C.; Martinez-de-Oliveira, J.; Queiroz, J.A.; Rodrigues, A.G. The anti-Candida activity of Thymbra capitata essential oil: Effect upon pre-formed biofilm. J. Ethnopharmacol. 2012, 140, 379–383. [Google Scholar] [CrossRef]
- Rosca, A.S.; Castro, J.; Sousa, L.G.V.; França, A.; Cavaleiro, C.; Salgueiro, L.; Cerca, N. Six bacterial vaginosis-associated species can form an in vitro and ex vivo polymicrobial biofilm that is susceptible to Thymbra capitata essential oil. Front. Cell. Infect. Microbiol. 2022, 12, 824860. [Google Scholar] [CrossRef]
- Vrenna, G.; Artini, M.; Ragno, R.; Relucenti, M.; Fiscarelli, E.V.; Tuccio Guarna Assanti, V.; Papa, R.; Selan, L. Anti-virulence properties of Coridothymus capitatus essential oil against Pseudomonas aeruginosa clinical isolates from cystic fibrosis patients. Microorganisms 2021, 9, 2257. [Google Scholar] [CrossRef]
- Bounatirou, S.; Smiti, S.; Miguel, M.G.; Faleiro, L.; Rejeb, M.N.; Neffati, M.; Costa, M.M.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G. Chemical composition, antioxidant and antibacterial activities of the essential oils isolated from Tunisian Thymus capitatus Hoff. et Link. Food Chem. 2007, 105, 146–155. [Google Scholar] [CrossRef]
- Ben Jemaa, M.; Falleh, H.; Serairi, R.; Neves, M.A.; Snoussi, M.; Isoda, H.; Nakajima, M.; RKsouri, R. Nanoencapsulated Thymus capitatus essential oil as natural preservative. Innov. Food Sci. Emerg. Technol. 2018, 45, 92–97. [Google Scholar] [CrossRef]
- Al Hafi, M.; El Beyrouthy, M.; Ouaini, N.; Stien, D.; Rutledge, D.; Chaillou, S. Chemical composition and antimicrobial activity of Satureja, Thymus, and Thymbra species grown in Lebanon. Chem. Biodivers. 2017, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Faleiro, L.; Miguel, G.; Gomes, S.; Costa, L.; Venâncio, F.; Teixeira, A.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G. Antibacterial and antioxidant activities of essential oils isolated from Thymbra capitata L. (Cav.) and Origanum vulgare L. J. Agric. Food Chem. 2005, 53, 8162–8168. [Google Scholar] [CrossRef] [PubMed]
- Džamić, A.M.; Nikolić, B.J.; Giweli, A.A.; Mitić-Ćulafić, D.S.; Soković, M.D.; Ristić, M.S.; Knežević-Vukčević, J.B.; Marin, P.D. Libyan Thymus capitatus essential oil: Antioxidant, antimicrobial, cytotoxic and colon pathogen adhesion-inhibition properties. J. Appl. Microbiol. 2015, 119, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Peñalver, P.; Huerta, B.; Borge, C.; Astorga, R.; Romero, R.; Perea, A. Antimicrobial activity of five essential oils against origin strains of the Enterobacteriaceae family. APMIS 2005, 113, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tagnaout, I.; Zerkani, H.; Hadi, N.; El Moumen, B.; El Makhoukhi, F.; Bouhrim, M.; Al-Salahi, R.; Nasr, F.A.; Mechchate, H.; Zair, T. Chemical composition, antioxidant and antibacterial activities of Thymus broussonetii Boiss and Thymus capitatus (L.) Hoffmann and Link essential oils. Plants 2022, 11, 954. [Google Scholar] [CrossRef] [PubMed]
- Moumni, S.; Elaissi, A.; Trabelsi, A.; Merghni, A.; Chraief, I.; Jelassi, B.; Chemli, R.; Ferchichi, S. Correlation between chemical composition and antibacterial activity of some Lamiaceae species essential oils from Tunisia. BMC Complement. Med. Ther. 2020, 20, 103. [Google Scholar] [CrossRef] [Green Version]
- Čabarkapa, I.; Čolović, R.; Đuragić, O.; Popović, S.; Kokić, B.; Milanov, D.; Pezo, L. Anti-biofilm activities of essential oils rich in carvacrol and thymol against Salmonella Enteritidis. Biofouling 2019, 35, 361–375. [Google Scholar] [CrossRef]
- Strantzali, D.; Kostoglou, D.; Perikleous, A.; Zestas, M.; Ornithopoulou, S.; Dubois-Brissonnet, F.; Giaouris, E. Comparative assessment of the disinfection effectiveness of thymol and benzalkonium chloride against adapted and non-adapted to thymol biofilm cells of a Salmonella Typhimurium epidemic phage type DT193 strain. Food Control 2021, 129, 108239. [Google Scholar] [CrossRef]
- Vetas, D.; Dimitropoulou, E.; Mitropoulou, G.; Kourkoutas, Y.; Giaouris, E. Disinfection efficiencies of sage and spearmint essential oils against planktonic and biofilm Staphylococcus aureus cells in comparison with sodium hypochlorite. Int. J. Food Microbiol. 2017, 257, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef] [PubMed]
- Kostoglou, D.; Protopappas, I.; Giaouris, E. Common plant-derived terpenoids present increased anti-biofilm potential against Staphylococcus bacteria compared to a quaternary ammonium biocide. Foods 2020, 9, 697. [Google Scholar] [CrossRef] [PubMed]
- Megdiche-Ksouri, W.; Saada, M.; Soumaya, B.; Snoussi, M.; Zaouali, Y.; Ksouri, R. Potential use of wild Thymus algeriensis and Thymus capitatus as source of antioxidant and antimicrobial agents. J. New Sci. 2015, 23, 11. [Google Scholar]
- Valerio, F.; Mezzapesa, G.N.; Ghannouchi, A.; Mondelli, D.; Logrieco, A.F.; Perrino, E.V. Characterization and antimicrobial properties of essential oils from four wild taxa of Lamiaceae family growing in Apulia. Agronomy 2021, 11, 1431. [Google Scholar] [CrossRef]
- Aljabeili, H.S.; Barakat, H.; Abdel-Rahman, H.A. Chemical composition, antibacterial and antioxidant activities of thyme essential oil (Thymus vulgaris). Food Nutr. Sci. 2018, 09, 433–446. [Google Scholar]
- Takó, M.; Kerekes, E.B.; Zambrano, C.; Kotogán, A.; Papp, T.; Krisch, J.; Vágvölgyi, C. Plant phenolics and phenolic-enriched extracts as antimicrobial agents against food-contaminating microorganisms. Antioxidants 2020, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Benoutman, A.; Erbiai, E.H.; Edderdaki, F.Z.; Cherif, E.K.; Saidi, R.; Lamrani, Z.; Pintado, M.; Pinto, E.; Esteves da Silva, J.C.G.; Maouni, A. Phytochemical composition, antioxidant and antifungal activity of Thymus capitatus, a medicinal plant collected from northern Morocco. Antibiotics 2022, 11, 681. [Google Scholar] [CrossRef]
- Tammar, S.; Salem, N.; Bettaieb Rebey, I.; Sriti, J.; Hammami, M.; Khammassi, S.; Marzouk, B.; Ksouri, R.; Msaada, K. Regional effect on essential oil composition and antimicrobial activity of Thymus capitatus L. J. Essent. Oil Res. 2019, 31, 129–137. [Google Scholar] [CrossRef]
- El-Jalel, L.F.A.; Elkady, W.M.; Gonaid, M.H.; El-Gareeb, K.A. Difference in chemical composition and antimicrobial activity of Thymus capitatus L. essential oil at different altitudes. Future J. Pharm. Sci. 2018, 4, 156–160. [Google Scholar] [CrossRef]
- Hajlaoui, H.; Mighri, H.; Aouni, M. Screening of antibacterial, antifungal and antioxidant activities of Tunisian Thymus capitatus (L.) Hoffm. et Link. essential oil. Tunis. J. Med. Plants Nat. Prod. 2015, 13, 20–38. [Google Scholar]
- Mkaddem, M.G.; Romdhane, M.; Ibrahim, H.; Ennajar, M.; Lebrihi, A.; Mathieu, F.; Bouajila, J. Essential oil of Thymus capitatus Hoff. et Link. from Matmata, Tunisia: Gas chromatography-mass spectrometry analysis and antimicrobial and antioxidant activities. J. Med. Food. 2010, 13, 1500–1504. [Google Scholar] [CrossRef] [PubMed]
- Hanoglu, A.; Hanoglu, D.Y.; Demirci, B.; Yavuz, D.Ö. Chemical composition of essential oil of the aerial parts of wild growing Thymus capitatus (L.) Hoffm. & link species collected from three different locations in northern Cyprus. J. Essent. Oil Bear. Plants 2017, 20, 546–551. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Shukla, I.; Sharifi-Rad, M.; Contreras, M.D.M.; Segura-Carretero, A.; Fathi, H.; Nasrabadi, N.N.; Kobarfard, F.; Sharifi-Rad, J. Thymol, thyme, and other plant sources: Health and potential uses. Phytother. Res. 2018, 32, 1688–1706. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, M.; Varoni, E.M.; Iriti, M.; Martorell, M.; Setzer, W.N.; Del Mar Contreras, M.; Salehi, B.; Soltani-Nejad, A.; Rajabi, S.; Tajbakhsh, M.; et al. Carvacrol and human health: A comprehensive review. Phytother. Res. 2018, 32, 1675–1687. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; van Stee, L.L.P.; Williams, J.; Beens, J.; Adahchour, M.; Vreuls, R.J.J.; Brinkman, U.A.Th.; Lelieveld, J. Comprehensive two-dimensional gas chromatography (GC×GC) measurements of volatile organic compounds in the atmosphere. Atmos. Chem. Phys. 2003, 3, 665–682. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Lai, W.-C.; Madhusudan Reddy, K.; Wei, B. Temperature-programmed retention indices for GC and GC-MS of hydrocarbon fuels and simulated distillation GC of heavy oils in Analytical advances for hydrocarbon research. In Analytical Advances for Hydrocarbon Research. Modern Analytical Chemistry; Hsu, C.S., Ed.; Springer: Boston, MA, USA, 2003; pp. 147–210. [Google Scholar] [CrossRef]
- Vagionas, K.; Ngassapa, O.; Runyoro, D.; Graikou, K.; Gortzi, O.; Chinou, I. Chemical analysis of edible aromatic plants growing in Tanzania. Food Chem. 2007, 105, 1711–1717. [Google Scholar] [CrossRef]
- Yu, E.J.; Kim, T.H.; Kim, K.H.; Lee, H.J. Characterization of aroma-active compounds of Abies nephrolepis (Khingan fir) needles using aroma extract dilution analysis. Flavour Fragr. J. 2004, 19, 74–79. [Google Scholar] [CrossRef]
- Smadja, J.; Rondeau, P.; Sing, A.S.C. Volatile constituents of five Citrus Petitgrain essential oils from Reunion. Flavour Fragr. J. 2005, 20, 399–402. [Google Scholar] [CrossRef]
- Weissbecker, B.; Holighaus, G.; Schütz, S. Gas chromatography with mass spectrometric and electroantennographic detection: Analysis of wood odorants by direct coupling of insect olfaction and mass spectrometry. J. Chromatogr. A 2004, 1056, 209–216. [Google Scholar] [CrossRef]
- Saroglou, V.; Dorizas, N.; Kypriotakis, Z.; Skaltsa, H.D. Analysis of the essential oil composition of eight Anthemis species from Greece. J. Chromatogr. A 2006, 1104, 313–322. [Google Scholar] [CrossRef]
- Afsharypuor, S.; Suleimany, M. Volatile oil constituents of Brassica oleracea var. gongylodes seeds. J. Essent. Oil Res. 2002, 14, 18–19. [Google Scholar] [CrossRef]
- Estevez, M.; Ventanas, S.; Ramirez, R.; Cava, R. Influence of the addition of rosemary essential oil on the volatiles pattern of porcine frankfurters. J. Agric. Food Chem. 2005, 53, 8317–8324. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.-X.; Liang, Y.-Z.; Fang, H.-Z.; Li, X.-N. Temperature-programmed retention indices for gas chromatography-mass spectroscopy analysis of plant essential oils. J. Chromatogr. A 2005, 1096, 76–85. [Google Scholar] [CrossRef]
- Mardarowicz, M.; Wianowska, D.; Dawidowicz, A.L.; Sawicki, R. The influence of sample treatment on SPME extracts from conifers. I. Comparison of terpene composition in Engelmann Spruce (Picea engelmanii) using hydrodistillation, SPME and PLE. Ann. Univ. Mariae Curie-Sklodowska Lub.—Pol. 2004, 59, 25–42. [Google Scholar] [CrossRef]
- Cho, I.H.; Namgung, H.-J.; Choi, H.-K.; Kim, Y.-S. Volatiles and key odorants in the pileus and stipe of pine-mushroom (Tricholoma matsutake Sing.). Food Chem. 2008, 106, 71–76. [Google Scholar] [CrossRef]
- Andriamaharavo, N.R. Retention Data. NIST Mass Spectrometry Data Center. Available online: https://chemdata.nist.gov/ (accessed on 12 December 2022)2014.
- Zhao, C.X.; Li, X.N.; Liang, Y.Z.; Fang, H.Z.; Huang, L.F.; Guo, F.Q. Comparative analysis of chemical components of essential oils from different samples of Rhododendron with the help of chemometrics methods. Chemom. Intell. Lab. Syst. 2006, 82, 218–228. [Google Scholar] [CrossRef]
- Kostaki, M.; Chorianopoulos, N.; Braxou, E.; Nychas, G.J.; Giaouris, E. Differential biofilm formation and chemical disinfection resistance of sessile cells of Listeria monocytogenes strains under monospecies and dual-species (with Salmonella enterica) conditions. Appl. Environ. Microbiol. 2012, 78, 2586–2595. [Google Scholar] [CrossRef] [Green Version]
- Kostoglou, D.; Tsaklidou, P.; Iliadis, I.; Garoufallidou, N.; Skarmoutsou, G.; Koulouris, I.; Giaouris, E. Advanced killing potential of thymol against a time and temperature optimized attached Listeria monocytogenes population in lettuce broth. Biomolecules 2021, 11, 397. [Google Scholar] [CrossRef]
- Schleifstein, J.I.; Coleman, M.B. An unidentified microorganism resembling B. ligniere and Past. pseudotuberculosis, and pathogenic for man. N. Y. State J. Med. 1939, 39, 1749–1753. [Google Scholar]
- Dimou, I.; Dritsas, S.; Aggelopoulou, P.; Vassilatou, K.; Damianaki, S.; Giaouris, E. Development of a herbal mouthwash containing a mixture of essential oils and plant extracts and in vitro testing of its antimicrobial efficiency against the planktonic and biofilm-enclosed cariogenic bacterium Streptococcus mutans. Biofouling 2021, 37, 397–409. [Google Scholar] [CrossRef]
- Postali, E.; Peroukidou, P.; Giaouris, E.; Papachristoforou, A. Investigating possible synergism in the antioxidant and antibacterial actions of honey and propolis from the Greek Island of Samothrace through their combined application. Foods 2022, 11, 2041. [Google Scholar] [CrossRef] [PubMed]
- Apak, R.; Güçlü, K.; Ozyürek, M.; Karademir, S.E. Novel total antioxidant capacity index for dietary polyphenols and vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef] [PubMed]
- Mantzourani, I.; Daoutidou, M.; Dasenaki, M.; Nikolaou, A.; Alexopoulos, A.; Terpou, A.; Thomaidis, N.; Plessas, S. Plant extract and essential oil application against food-borne pathogens in raw pork meat. Foods 2022, 11, 861. [Google Scholar] [CrossRef] [PubMed]
- IUPAC. Compendium of Chemical Terminology, 2nd ed.; (the “Gold Book”); McNaught, A.D., Wilkinson, A., Eds.; Blackwell Scientific Publications: Oxford, UK, 1997; Online (2019-) created by S. J. Chalk. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Bacterial Species | Antibacterial Action | Antibiofilm Action | ||||||
|---|---|---|---|---|---|---|---|---|
| MIC 1 | MBC 2 | MBIC 3 | MBEC 4 | |||||
| % (v/v) | mg/mL | % (v/v) | mg/mL | % (v/v) | mg/mL | % (v/v) | mg/mL | |
| S. Typhimurium | 0.06 | 0.6 | 0.06 | 0.6 | 0.13 | 1.1 | 3.13 (= 52.2 × MΒC) | 28.4 |
| L. monocytogenes | 0.03 | 0.3 | 0.06 | 0.6 | 0.03 | 0.3 | 6.25 (= 104.2 × MΒC) | 56.9 |
| Y. enterocolitica | 0.03 | 0.3 | 0.03 | 0.3 | 0.03 | 0.3 | 2.00 (= 66.7 × MΒC) | 18.2 |
| Method | DPPH• (μmol TRE 1/g EO) | CUPRAC (μmol TRE 1/g EO) | TPC (mg GAE 2/g EO) |
|---|---|---|---|
| Results | 144.66 ± 1.71 | 763.90 ± 13.93 | 231.32 ± 16.71 |
| Compounds Detected | GC-FID | GC-MS | ||
|---|---|---|---|---|
| % Area | LRΙ | LRIref | % Area | |
| methyl-cyclopentane | <700 | <700 [68] | 0.8 | |
| Cyclohexane | <700 | <700 [69] | 0.1 | |
| α-Thujene | 930 | 930 [70] | 0.2 | |
| α-Pinene | 2.9 | 936 | 937 [70] | 2.8 |
| Camphene | 949 | 951 [70] | 1.6 | |
| β-Pinene | 1.7 | 977 | 978 [70] | 0.3 |
| β-Myrcene | 1.5 | 999 | 992 [71] | 1.0 |
| α-Phellandrene | 1008 | 1006 [72] | 0.2 | |
| 3-Carene | 1012 | 1015 [73] | <0.1 | |
| α-Terpinene (1-methyl-4-(1-methylethyl)-1,3-cyclohexadiene) | 0.2 | 1020 | 1016 [74] | 0.4 |
| Carvomenthene (1-methyl-4-(1-methylethyl)-cyclohexene) | 1024 | - | 0.1 | |
| p-Cymene (1-methyl-4-(1-methylethyl)-benzene) | 28.9 | 1034 | 1030 [75] | 31.0 |
| Limonene | 4.2 | 1035 | 1033 [75] | 3.9 |
| p-Cymenene (1-methyl-4-(1-methylethenyl)-benzene) | 1044 | 1078 [72] | 0.1 | |
| γ-Terpinene (1-methyl-4-(1-methylethyl)-1,4-cyclohexadiene) | 2.8 | 1070 | 1063 [73] | 4.3 |
| β-Thujone ([1S-(1α,4β,5α)]-4-methyl-1-(1-methylethyl)-bicyclo[3.1.0]hexan-3-one) | 1114 | 1116 [74] | 0.1 | |
| Linalool (3,7-dimethyl-1,6-octadien-3-ol) | 5.1 | 1123 | 1121 [76] | 3.8 |
| Camphor | 1154 | 1152 [77] | 0.2 | |
| β-Terpineol (1-methyl-4-(1-methylethenyl)-cyclohexanol) | 1169 | 1179 [76] | <0.1 | |
| Isoborneol | 1173 | 1164 [78] | <0.1 | |
| Borneol | 1183 | 1174 [73] | 0.9 | |
| Terpinen-4-ol (4-methyl-1-(1-methylethyl)-3-cyclohexen-1-ol) | 1191 | 1187 [71] | 0.3 | |
| α-Terpineol (α,α-4-trimethyl-3-cyclohexene-1-methanol) | 0.9 | 1209 | 1207 [79] | 0.2 |
| γ-Terpineol (1-methyl-4-(1-methylethylidene)-cyclohexanol) | 1222 | 1218 [80] | <0.1 | |
| Thymol | 44.5 | 1341 | 1302 [74] | 39.8 |
| Carvacrol (2-methyl-5-(1-methylethyl)-phenol) | 4.2 | 1346 | 1311 [74] | 5.7 |
| Caryophyllene | 1.0 | 1439 | 1437 [81] | 1.1 |
| α-Caryophyllene | 1475 | 1465 [81] | 0.1 | |
| Caryophyllene oxide | 1610 | 1595 [71] | 0.2 | |
| Total | 97.9 | 99.2 | ||
| Bacterial Species | Gram Reaction | Strain Code | Isolation Origin | Other Strain Information | Reference |
|---|---|---|---|---|---|
| Salmonellla enterica | - | FMCC 1_B137 | human, salmonellosis outbreak | serovar Typhimurium, phage type DT193 | [82] |
| Listeria monocytogenes | + | AAL 2 20107 | mixed green salad | serovar 1/2b | [83] |
| Yersinia enterocolitica | - | DSM 3 4780 | human, glanders-like infection of the face | subsp. enterocolitica, type strain, ATCC 4 33114 | [84] |
| Bacterial Species | Strain Code | Growth Medium | Temperature (°C) | Medium Renewal (h) |
|---|---|---|---|---|
| S. Typhimurium | FMCC_B137 | 1/10 TSB 1 | 20 | 48 |
| L. monocytogenes | AAL 20107 | BHI 2 broth | 37 | 48 |
| Y. enterocolitica | DSM 4780 | TSB | 20 | 48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maniki, E.; Kostoglou, D.; Paterakis, N.; Nikolaou, A.; Kourkoutas, Y.; Papachristoforou, A.; Giaouris, E. Chemical Composition, Antioxidant, and Antibiofilm Properties of Essential Oil from Thymus capitatus Plants Organically Cultured on the Greek Island of Lemnos. Molecules 2023, 28, 1154. https://doi.org/10.3390/molecules28031154
Maniki E, Kostoglou D, Paterakis N, Nikolaou A, Kourkoutas Y, Papachristoforou A, Giaouris E. Chemical Composition, Antioxidant, and Antibiofilm Properties of Essential Oil from Thymus capitatus Plants Organically Cultured on the Greek Island of Lemnos. Molecules. 2023; 28(3):1154. https://doi.org/10.3390/molecules28031154
Chicago/Turabian StyleManiki, Eirini, Dimitra Kostoglou, Nikolaos Paterakis, Anastasios Nikolaou, Yiannis Kourkoutas, Alexandros Papachristoforou, and Efstathios Giaouris. 2023. "Chemical Composition, Antioxidant, and Antibiofilm Properties of Essential Oil from Thymus capitatus Plants Organically Cultured on the Greek Island of Lemnos" Molecules 28, no. 3: 1154. https://doi.org/10.3390/molecules28031154
APA StyleManiki, E., Kostoglou, D., Paterakis, N., Nikolaou, A., Kourkoutas, Y., Papachristoforou, A., & Giaouris, E. (2023). Chemical Composition, Antioxidant, and Antibiofilm Properties of Essential Oil from Thymus capitatus Plants Organically Cultured on the Greek Island of Lemnos. Molecules, 28(3), 1154. https://doi.org/10.3390/molecules28031154