


Potential Anti-Rheumatoid Arthritis Activities and Mechanisms of Ganoderma lucidum Polysaccharides
Abstract
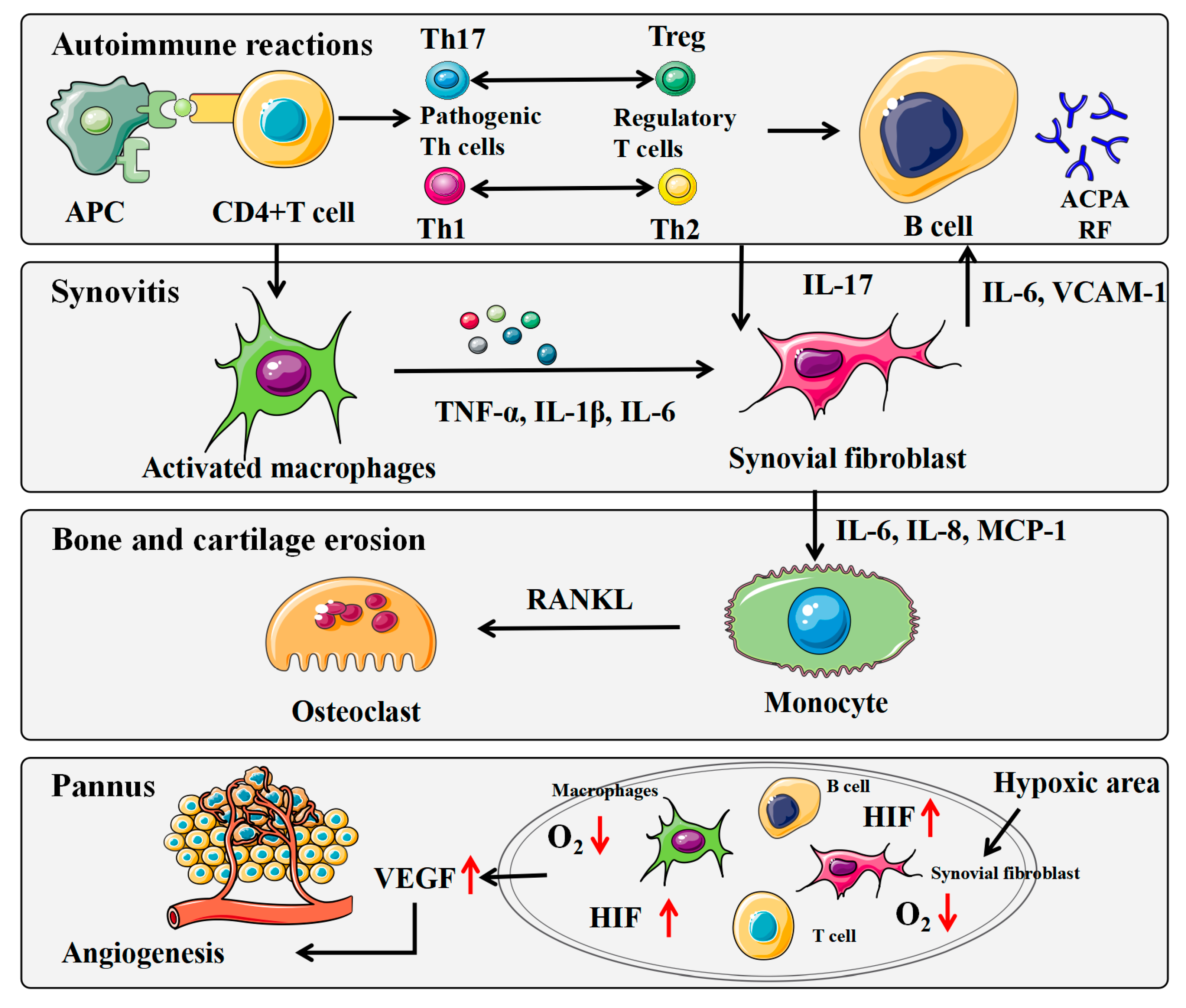
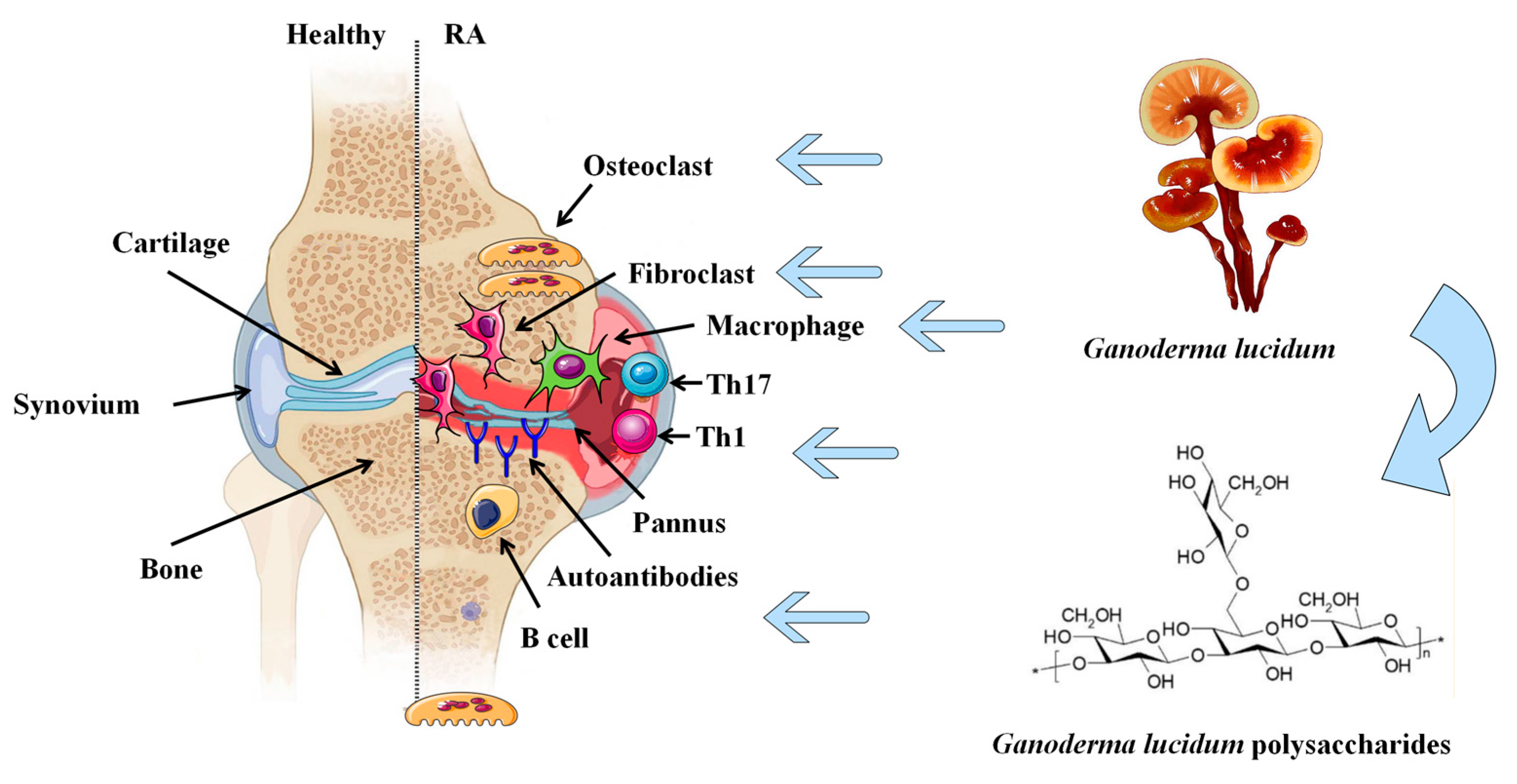
:1. Introduction
2. Methods
3. Inhibition of GLP on Synovial Joint Inflammation
3.1. Behavior of Synoviocytes
3.2. Secretion of Inflammatory Cytokines
3.3. Expression of Signaling Pathways
4. Regulation of GLP on Immune Response
4.1. Modulatory Effect of GLP on DCs
4.2. Modulatory Effect of GLP on T Cells
4.3. Modulatory Effect of GLP on B Cells
4.4. Modulatory Effect of GLP on Macrophages
5. Suppression of GLP on Angiogenesis
6. Reduction of GLP on Bone Erosion
7. Resistance of GLP on Anemia
8. Nano-Formulation of GLP
9. Safety of GLP
10. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid Arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef] [PubMed]
- Finckh, A.; Gilbert, B.; Hodkinson, B.; Bae, S.C.; Thomas, R.; Deane, K.D.; Alpizar-Rodriguez, D.; Lauper, K. Global Epidemiology of Rheumatoid Arthritis. Nat. Rev. Rheumatol. 2022, 18, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Scherer, H.U.; Häupl, T.; Burmester, G.R. The Etiology of Rheumatoid Arthritis. J. Autoimmun. 2020, 110, 102400. [Google Scholar] [CrossRef]
- van Delft, M.A.; Huizinga, T.W. An Overview of Autoantibodies in Rheumatoid Arthritis. J. Autoimmun. 2020, 110, 102392. [Google Scholar] [CrossRef] [PubMed]
- Sparks, J.A. Rheumatoid Arthritis. Ann. Intern. Med. 2019, 170, Itc1–Itc16. [Google Scholar] [CrossRef]
- Cush, J.J. Rheumatoid Arthritis: Early Diagnosis and Treatment. Med. Clin. N. Am. 2021, 105, 355–365. [Google Scholar] [CrossRef]
- Figus, F.A.; Piga, M.; Azzolin, I.; McConnell, R.; Iagnocco, A. Rheumatoid Arthritis: Extra-Articular Manifestations and Comorbidities. Autoimmun. Rev. 2021, 20, 102776. [Google Scholar] [CrossRef]
- Giannini, D.; Antonucci, M.; Petrelli, F.; Bilia, S.; Alunno, A.; Puxeddu, I. One Year in Review 2020: Pathogenesis of Rheumatoid Arthritis. Clin. Exp. Rheumatol. 2020, 38, 387–397. [Google Scholar] [CrossRef]
- Jang, S.; Kwon, E.J.; Lee, J.J. Rheumatoid Arthritis: Pathogenic Roles of Diverse Immune Cells. Int. J. Mol. Sci. 2022, 23, 905. [Google Scholar] [CrossRef]
- Mueller, A.L.; Payandeh, Z.; Mohammadkhani, N.; Mubarak, S.M.H.; Zakeri, A.; Bahrami, A.A.; Brockmueller, A.; Shakibaei, M. Recent Advances in Understanding the Pathogenesis of Rheumatoid Arthritis: New Treatment Strategies. Cells 2021, 10, 3017. [Google Scholar] [CrossRef]
- Croia, C.; Bursi, R.; Sutera, D.; Petrelli, F.; Alunno, A.; Puxeddu, I. One Year in Review 2019: Pathogenesis of Rheumatoid Arthritis. Clin. Exp. Rheumatol. 2019, 37, 347–357. [Google Scholar] [PubMed]
- Abbasi, M.; Mousavi, M.J.; Jamalzehi, S.; Alimohammadi, R.; Bezvan, M.H.; Mohammadi, H.; Aslani, S. Strategies toward Rheumatoid Arthritis Therapy; the Old and the New. J. Cell Physiol. 2019, 234, 10018–10031. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, H.; Deng, R. Angiogenesis as a Potential Treatment Strategy for Rheumatoid Arthritis. Eur. J. Pharmacol. 2021, 910, 174500. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Fu, X.; Chen, X.; Li, Z.; Huang, Y.; Liang, C. Promising Therapeutic Targets for Treatment of Rheumatoid Arthritis. Front. Immunol. 2021, 12, 686155. [Google Scholar] [CrossRef] [PubMed]
- Prasad, P.; Verma, S.; Surbhi, N.; Ganguly, K.; Chaturvedi, V.; Mittal, S.A. Rheumatoid Arthritis: Advances in Treatment Strategies. Mol. Cell Biochem. 2023, 478, 69–88. [Google Scholar] [CrossRef] [PubMed]
- García-González, C.M.; Baker, J. Treatment of Early Rheumatoid Arthritis: Methotrexate and Beyond. Curr. Opin. Pharmacol. 2022, 64, 102227. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, S.; Du, K.; Liang, C.; Wang, S.; Boadi, E.O.; Li, J.; Pang, X.; He, J.; Chang, Y.X. Traditional Herbal Medicine: Therapeutic Potential in Rheumatoid Arthritis. J. Ethnopharmacol. 2021, 279, 114368. [Google Scholar] [CrossRef] [PubMed]
- Zeng, P.; Chen, Y.; Zhang, L.; Xing, M. Ganoderma lucidum Polysaccharide Used for Treating Physical Frailty in China. Prog. Mol. Biol. Transl. Sci. 2019, 163, 179–219. [Google Scholar]
- Ahmad, M.F. Ganoderma lucidum: Persuasive Biologically Active Constituents and Their Health Endorsement. Biomed. Pharmacother. 2018, 107, 507–519. [Google Scholar] [CrossRef]
- Lam, F.F.; Ko, I.W.; Ng, E.S.; Tam, L.S.; Leung, P.C.; Li, E.K. Analgesic and Anti-Arthritic Effects of Lingzhi and San Miao San Supplementation in a Rat Model of Arthritis Induced by Freund′s Complete Adjuvant. J. Ethnopharmacol. 2008, 120, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.X.; Wong, C.K.; Li, E.K.; Tam, L.S.; Leung, P.C.; Yin, Y.B.; Lam, C.W. Immunomodulatory Effects of Lingzhi and San-Miao-San Supplementation on Patients with Rheumatoid Arthritis. Immunopharmacol. Immunotoxicol. 2006, 28, 197–200. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, R.; Riaz, M.; Khan, A.; Aljamea, A.; Algheryafi, M.; Sewaket, D.; Alqathama, A. Ganoderma lucidum (Reishi) an Edible Mushroom; a Comprehensive and Critical Review of Its Nutritional, Cosmeceutical, Mycochemical, Pharmacological, Clinical, and Toxicological Properties. Phytother. Res. 2021, 35, 6030–6062. [Google Scholar] [CrossRef] [PubMed]
- Seweryn, E.; Ziała, A.; Gamian, A. Health-Promoting of Polysaccharides Extracted from Ganoderma lucidum. Nutrients 2021, 13, 2725. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, N.; Bhatt, L.K.; Prabhavalkar, K.S. Experimental Animal Models for Rheumatoid Arthritis. Immunopharmacol. Immunotoxicol. 2018, 40, 193–200. [Google Scholar] [CrossRef]
- Bessis, N.; Decker, P.; Assier, E.; Semerano, L.; Boissier, M.C. Arthritis Models: Usefulness and Interpretation. Semin. Immunopathol. 2017, 39, 469–486. [Google Scholar] [CrossRef]
- Damerau, A.; Gaber, T. Modeling Rheumatoid Arthritis In Vitro: From Experimental Feasibility to Physiological Proximity. Int. J. Mol. Sci. 2020, 21, 7916. [Google Scholar] [CrossRef] [PubMed]
- Kondo, N.; Kuroda, T.; Kobayashi, D. Cytokine Networks in the Pathogenesis of Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 10922. [Google Scholar] [CrossRef] [PubMed]
- O′Neil, L.J.; Kaplan, M.J. Neutrophils in Rheumatoid Arthritis: Breaking Immune Tolerance and Fueling Disease. Trends Mol. Med. 2019, 25, 215–227. [Google Scholar] [CrossRef]
- Rivellese, F.; Pitzalis, C. Cellular and Molecular Diversity in Rheumatoid Arthritis. Semin. Immunol. 2021, 58, 101519. [Google Scholar] [CrossRef]
- Mao, X.; Li, W.; Chen, W.; Li, Y.; Wang, Q.; Wang, X.; Pi, Z.; Wang, D.; Xu, H.; Guo, Q.; et al. Exploring and Characterizing a Novel Combination of Paeoniflorin and Talatizidine for the Treatment of Rheumatoid Arthritis. Pharmacol. Res. 2020, 153, 104658. [Google Scholar] [CrossRef]
- Yang, X.; Zhan, N.; Jin, Y.; Ling, H.; Xiao, C.; Xie, Z.; Zhong, H.; Yu, X.; Tang, R.; Ma, J.; et al. Tofacitinib Restores the Balance of Γδtreg/Γδt17 Cells in Rheumatoid Arthritis by Inhibiting the Nlrp3 Inflammasome. Theranostics 2021, 11, 1446–1457. [Google Scholar] [CrossRef]
- Yoshitomi, H. Regulation of Immune Responses and Chronic Inflammation by Fibroblast-Like Synoviocytes. Front. Immunol. 2019, 10, 1395. [Google Scholar] [CrossRef] [Green Version]
- Masoumi, M.; Mehrabzadeh, M.; Mahmoudzehi, S.; Mousavi, M.J.; Jamalzehi, S.; Sahebkar, A.; Karami, J. Role of Glucose Metabolism in Aggressive Phenotype of Fibroblast-Like Synoviocytes: Latest Evidence and Therapeutic Approaches in Rheumatoid Arthritis. Int. Immunopharmacol. 2020, 89, 107064. [Google Scholar] [CrossRef]
- Ho, Y.W.; Yeung, J.S.; Chiu, P.K.; Tang, W.M.; Lin, Z.B.; Man, R.Y.; Lau, C.S. Ganoderma lucidum Polysaccharide Peptide Reduced the Production of Proinflammatory Cytokines in Activated Rheumatoid Synovial Fibroblast. Mol. Cell. Biochem. 2007, 301, 173–179. [Google Scholar] [CrossRef]
- Chen, Z.; Bozec, A.; Ramming, A.; Schett, G. Anti-Inflammatory and Immune-Regulatory Cytokines in Rheumatoid Arthritis. Nat. Rev. Rheumatol. 2019, 15, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Markovics, A.; Rosenthal, K.S.; Mikecz, K.; Carambula, R.E.; Ciemielewski, J.C.; Zimmerman, D.H. Restoring the Balance between Pro-Inflammatory and Anti-Inflammatory Cytokines in the Treatment of Rheumatoid Arthritis: New Insights from Animal Models. Biomedicines 2021, 10, 44. [Google Scholar] [CrossRef]
- Mateen, S.; Zafar, A.; Moin, S.; Khan, A.Q.; Zubair, S. Understanding the Role of Cytokines in the Pathogenesis of Rheumatoid Arthritis. Clin. Chim. Acta 2016, 455, 161–171. [Google Scholar] [CrossRef]
- Kim, E.Y.; Moudgil, K.D. Immunomodulation of Autoimmune Arthritis by Pro-Inflammatory Cytokines. Cytokine 2017, 98, 87–96. [Google Scholar] [CrossRef]
- Hu, Y.; Wang, S.X.; Wu, F.Y.; Wu, K.J.; Shi, R.P.; Qin, L.H.; Lu, C.F.; Wang, S.Q.; Wang, F.F.; Zhou, S. Effects and Mechanism of Ganoderma lucidum Polysaccharides in the Treatment of Diabetic Nephropathy in Streptozotocin-Induced Diabetic Rats. BioMed Res. Int. 2022, 2022, 4314415. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Sheng, Z.; Wang, J.; Jiang, Y.; Yang, B. Structure of Water-Soluble Polysaccharides in Spore of Ganoderma lucidum and Their Anti-Inflammatory Activity. Food Chem. 2022, 373, 131374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liu, Y.; Zhao, X.; Tang, Q.; Dernedde, J.; Zhang, J.; Fan, H. Anti-Inflammatory Properties of Glpss58, a Sulfated Polysaccharide from Ganoderma lucidum. Int. J. Biol. Macromol. 2018, 107, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.X.; Wang, B.X.; Fei, X.F.; Zhang, J.; Gong, Y.J.; Minami, M.; Nagata, T.; Ikejima, T. Effects of Polysaccharides (Fi0-C) from Mycelium of Ganoderma tsugae on Proinflammatory Cytokine Production by Thp-1 Cells and Human Pbmc (II). Acta Pharmacol. Sin. 2000, 21, 1186–1192. [Google Scholar] [PubMed]
- Fang, L.; Zhao, Q.; Guo, C.; Guo, D.; Li, Z.; Xu, J.; Guo, C.; Sang, T.; Wang, Y.; Chen, J.; et al. Removing the Sporoderm from the Sporoderm-Broken Spores of Ganoderma lucidum Improves the Anticancer and Immune-Regulatory Activity of the Water-Soluble Polysaccharide. Front. Nutr. 2022, 9, 1006127. [Google Scholar] [CrossRef]
- Cao, Q.Z.; Lin, S.Q.; Wang, S.Z. Effect of Ganoderma lucidum Polysaccharides Peptide on Invasion of Human Lung Carcinoma Cells In Vitro. Beijing Da Xue Xue Bao Yi Xue Ban 2007, 39, 653–656. [Google Scholar]
- Wang, J.; Zhang, Y.; Yuan, Y.; Yue, T. Immunomodulatory of Selenium Nano-Particles Decorated by Sulfated Ganoderma lucidum Polysaccharides. Food Chem. Toxicol. 2014, 68, 183–189. [Google Scholar] [CrossRef]
- Li, H.; Xiao, Y.; Han, L.; Jia, Y.; Luo, S.; Zhang, D.; Zhang, L.; Wu, P.; Xiao, C.; Kan, W.; et al. Ganoderma lucidum Polysaccharides Ameliorated Depression-Like Behaviors in the Chronic Social Defeat Stress Depression Model Via Modulation of Dectin-1 and the Innate Immune System. Brain Res. Bull. 2021, 171, 16–24. [Google Scholar] [CrossRef]
- Liu, S.; Ma, H.; Zhang, H.; Deng, C.; Xin, P. Recent Advances on Signaling Pathways and Their Inhibitors in Rheumatoid Arthritis. Clin. Immunol. 2021, 230, 108793. [Google Scholar] [CrossRef]
- Yu, X.; Zhou, J.; Zhao, F.; Liu, X.; Mao, Y.; Diao, L.; Wen, C.; Liu, M. Tomatidine Suppresses the Destructive Behaviors of Fibroblast-Like Synoviocytes and Ameliorates Type II Collagen-Induced Arthritis in Rats. Front. Pharmacol. 2021, 12, 670707. [Google Scholar] [CrossRef]
- Shang, Z.Z.; Qin, D.Y.; Li, Q.M.; Zha, X.Q.; Pan, L.H.; Peng, D.Y.; Luo, J.P. Dendrobium Huoshanense Stem Polysaccharide Ameliorates Rheumatoid Arthritis in Mice Via Inhibition of Inflammatory Signaling Pathways. Carbohydr. Polym. 2021, 258, 117657. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.Y.; Wang, H.; Chen, X.Y.; Zhang, L.; Yuan, Y. An Alcohol Extract Prepared from the Male Flower of Eucommia Ulmoides Oliv. Promotes Synoviocyte Apoptosis and Ameliorates Bone Destruction in Rheumatoid Arthritis. Chin. Med. 2021, 16, 113. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Tan, L.; Lu, T.; Wang, K.; Wang, H.; Han, B.; Zhao, Y.; Chen, H.; Li, Y.; Chen, H.; et al. Glytabastan B, a Coumestan Isolated from Glycine tabacina, Alleviated Synovial Inflammation, Osteoclastogenesis and Collagen-Induced Arthritis through Inhibiting Mapk and Pi3k/Akt Pathways. Biochem. Pharmacol. 2022, 197, 114912. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.J.; Lee, C.W.; Sung, H.C.; Chen, Y.H.; Chiang, Y.C.; Hsu, H.Y.; Tseng, Y.C.; Li, C.Y.; Wang, S.H.; Chen, Y.L. Ganoderma lucidum Polysaccharides Reduce Lipopolysaccharide-Induced Interleukin-1 Β Expression in Cultured Smooth Muscle Cells and in Thoracic Aortas in Mice. Evid. Based Complement. Alternat. Med. 2014, 2014, 305149. [Google Scholar] [CrossRef] [Green Version]
- Sang, T.; Guo, C.; Guo, D.; Wu, J.; Wang, Y.; Wang, Y.; Chen, J.; Chen, C.; Wu, K.; Na, K.; et al. Suppression of Obesity and Inflammation by Polysaccharide from Sporoderm-Broken Spore of Ganoderma lucidum Via Gut Microbiota Regulation. Carbohydr. Polym. 2021, 256, 117594. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yuan, Y.; Yue, T. Immunostimulatory Activities of Β-D-Glucan from Ganoderma lucidum. Carbohydr. Polym. 2014, 102, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, N.; Takayanagi, H. Mechanisms of Joint Destruction in Rheumatoid Arthritis—Immune Cell-Fibroblast-Bone Interactions. Nat. Rev. Rheumatol. 2022, 18, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Falconer, J.; Murphy, A.N.; Young, S.P.; Clark, A.R.; Tiziani, S.; Guma, M.; Buckley, C.D. Review: Synovial Cell Metabolism and Chronic Inflammation in Rheumatoid Arthritis. Arthritis Rheumatol. 2018, 70, 984–999. [Google Scholar] [CrossRef] [Green Version]
- Weyand, C.M.; Goronzy, J.J. The Immunology of Rheumatoid Arthritis. Nat. Immunol. 2021, 22, 10–18. [Google Scholar] [CrossRef]
- Brandum, E.P.; Jørgensen, A.S.; Rosenkilde, M.M.; Hjortø, G.M. Dendritic Cells and Ccr7 Expression: An Important Factor for Autoimmune Diseases, Chronic Inflammation, and Cancer. Int. J. Mol. Sci. 2021, 22, 8340. [Google Scholar] [CrossRef]
- Karmakar, U.; Vermeren, S. Crosstalk between B Cells and Neutrophils in Rheumatoid Arthritis. Immunology 2021, 164, 689–700. [Google Scholar] [CrossRef]
- Guo, C.; Guo, D.; Fang, L.; Sang, T.; Wu, J.; Guo, C.; Wang, Y.; Wang, Y.; Chen, C.; Chen, J.; et al. Ganoderma lucidum Polysaccharide Modulates Gut Microbiota and Immune Cell Function to Inhibit Inflammation and Tumorigenesis in Colon. Carbohydr. Polym. 2021, 267, 118231. [Google Scholar] [CrossRef]
- Lu, J.; He, R.; Sun, P.; Zhang, F.; Linhardt, R.J.; Zhang, A. Molecular Mechanisms of Bioactive Polysaccharides from Ganoderma lucidum (Lingzhi), a Review. Int. J. Biol. Macromol. 2020, 150, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Suzuki, M.; Sakaguchi, R.; Tani, I.; Kotani, H.; Shudo, N.; Yoshimura, A. Preferential Induction of Th17 Cells In Vitro and in Vivo by Fucogalactan from Ganoderma lucidum (Reishi). Biochem. Biophys. Res. Commun. 2012, 422, 174–180. [Google Scholar] [CrossRef]
- Gao, X.X.; Fei, X.F.; Wang, B.X.; Zhang, J.; Gong, Y.J.; Minami, M.; Nagata, T.; Ikejima, T. Effects of Polysaccharides (Fi0-B) from Mycelium of Ganoderma tsugae on Proinflammatory Cytokine Production by Thp-1 Cells and Human Pbmc (I). Acta Pharmacol. Sin. 2000, 21, 1179–1185. [Google Scholar] [PubMed]
- Sohretoglu, D.; Huang, S. Ganoderma lucidum Polysaccharides as an Anti-Cancer Agent. Anticancer Agents Med. Chem. 2018, 18, 667–674. [Google Scholar] [CrossRef]
- Kladar, N.V.; Gavarić, N.S.; Božin, B.N. Ganoderma: Insights into Anticancer Effects. Eur. J. Cancer Prev. 2016, 25, 462–471. [Google Scholar] [CrossRef]
- Wehr, P.; Purvis, H.; Law, S.C.; Thomas, R. Dendritic Cells, T Cells and Their Interaction in Rheumatoid Arthritis. Clin. Exp. Immunol. 2019, 196, 12–27. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.K.; Law, H.K.; Lin, Z.B.; Lau, Y.L.; Chan, G.C. Response of Human Dendritic Cells to Different Immunomodulatory Polysaccharides Derived from Mushroom and Barley. Int. Immunol. 2007, 19, 891–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cope, A.P.; Schulze-Koops, H.; Aringer, M. The Central Role of T Cells in Rheumatoid Arthritis. Clin. Exp. Rheumatol. 2007, 25, S4–S11. [Google Scholar]
- Qi, G.; Hua, H.; Gao, Y.; Lin, Q.; Yu, G.Y. Effects of Ganoderma lucidum Spores on Sialoadenitis of Nonobese Diabetic Mice. Chin. Med. J. (Engl.) 2009, 122, 556–560. [Google Scholar]
- Notley, C.A.; Ehrenstein, M.R. The Yin and Yang of Regulatory T Cells and Inflammation in Ra. Nat. Rev. Rheumatol. 2010, 6, 572–577. [Google Scholar] [CrossRef]
- Jiang, Q.; Yang, G.; Liu, Q.; Wang, S.; Cui, D. Function and Role of Regulatory T Cells in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 626193. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, Y.; Liu, Q.; Chu, Y.; Mi, R.; Jiang, F.; Zhao, J.; Hu, K.; Luo, R.; Feng, Y.; et al. Malt1 Regulates Th2 and Th17 Differentiation Via Nf-Κb and Jnk Pathways, as Well as Correlates with Disease Activity and Treatment Outcome in Rheumatoid Arthritis. Front. Immunol. 2022, 13, 913830. [Google Scholar] [CrossRef]
- Wójcik, P.; Gęgotek, A.; Žarković, N.; Skrzydlewska, E. Oxidative Stress and Lipid Mediators Modulate Immune Cell Functions in Autoimmune Diseases. Int. J. Mol. Sci. 2021, 22, 723. [Google Scholar] [CrossRef] [PubMed]
- Peter, J.; Sabu, V.; Aswathy, I.S.; Krishnan, S.; Preethi, S.S.L.; Simon, M.; Helen, A. Dietary Amaranths Modulate the Immune Response Via Balancing Th1/Th2 and Th17/Treg Response in Collagen-Induced Arthritis. Mol. Cell Biochem. 2020, 472, 57–66. [Google Scholar] [CrossRef]
- Strehl, C.; Ehlers, L.; Gaber, T.; Buttgereit, F. Glucocorticoids-All-Rounders Tackling the Versatile Players of the Immune System. Front. Immunol. 2019, 10, 1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leipe, J.; Pirronello, F.; Schulze-Koops, H.; Skapenko, A. Altered T Cell Plasticity Favours Th17 Cells in Early Arthritis. Rheumatology 2020, 59, 2754–2763. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, S.; Li, H.; Wang, X.; Song, L.; Xue, J. Polysaccharide from Ganoderma lucidum Alleviates Cognitive Impairment in a Mouse Model of Chronic Cerebral Hypoperfusion by Regulating Cd4(+)Cd25(+)Foxp3(+) Regulatory T Cells. Food Funct. 2022, 13, 1941–1952. [Google Scholar] [CrossRef]
- Wei, B.; Zhang, R.; Zhai, J.; Zhu, J.; Yang, F.; Yue, D.; Liu, X.; Lu, C.; Sun, X. Suppression of Th17 Cell Response in the Alleviation of Dextran Sulfate Sodium-Induced Colitis by Ganoderma lucidum Polysaccharides. J. Immunol. Res. 2018, 2018, 2906494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habijanic, J.; Berovic, M.; Boh, B.; Plankl, M.; Wraber, B. Submerged Cultivation of Ganoderma lucidum and the Effects of Its Polysaccharides on the Production of Human Cytokines Tnf-A, Il-12, Ifn-Γ, Il-2, Il-4, Il-10 and Il-17. New Biotechnol. 2015, 32, 85–95. [Google Scholar] [CrossRef]
- Qin, Y.; Cai, M.L.; Jin, H.Z.; Huang, W.; Zhu, C.; Bozec, A.; Huang, J.; Chen, Z. Age-Associated B Cells Contribute to the Pathogenesis of Rheumatoid Arthritis by Inducing Activation of Fibroblast-Like Synoviocytes Via Tnf-A-Mediated Erk1/2 and Jak-Stat1 Pathways. Ann. Rheum. Dis. 2022, 81, 1504–1514. [Google Scholar] [CrossRef]
- Pala, O.; Diaz, A.; Blomberg, B.B.; Frasca, D. B Lymphocytes in Rheumatoid Arthritis and the Effects of Anti-Tnf-A Agents on B Lymphocytes: A Review of the Literature. Clin. Ther. 2018, 40, 1034–1045. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Gao, J.; Kang, J.; Wang, X.; Niu, Q.; Liu, J.; Zhang, L. B Cells in Rheumatoid Arthritis: Pathogenic Mechanisms and Treatment Prospects. Front. Immunol. 2021, 12, 750753. [Google Scholar] [CrossRef] [PubMed]
- Alivernini, S.; Tolusso, B.; Fedele, A.L.; Di Mario, C.; Ferraccioli, G.; Gremese, E. The B Side of Rheumatoid Arthritis Pathogenesis. Pharmacol. Res. 2019, 149, 104465. [Google Scholar] [CrossRef]
- Kurowska-Stolarska, M.; Alivernini, S. Synovial Tissue Macrophages in Joint Homeostasis, Rheumatoid Arthritis and Disease Remission. Nat. Rev. Rheumatol. 2022, 18, 384–397. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, Y.; Wu, R.; Ding, T.; Xue, H.; Gao, C.; Li, X.; Wang, C. New Insights from Single-Cell Sequencing Data: Synovial Fibroblasts and Synovial Macrophages in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 709178. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.A.; Devitt, A.; Johnson, J.R. Macrophages: The Good, the Bad, and the Gluttony. Front. Immunol. 2021, 12, 708186. [Google Scholar] [CrossRef]
- Cutolo, M.; Campitiello, R.; Gotelli, E.; Soldano, S. The Role of M1/M2 Macrophage Polarization in Rheumatoid Arthritis Synovitis. Front. Immunol. 2022, 13, 867260. [Google Scholar] [CrossRef]
- Liu, C.; Yang, N.; Song, Y.; Wang, L.; Zi, J.; Zhang, S.; Dunkin, D.; Busse, P.; Weir, D.; Tversky, J.; et al. Ganoderic Acid C1 Isolated from the Anti-Asthma Formula, Ashmi™ Suppresses Tnf-A Production by Mouse Macrophages and Peripheral Blood Mononuclear Cells from Asthma Patients. Int. Immunopharmacol. 2015, 27, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Ma, B.; Xue, F.; Xing, Y.; Wu, P.; Li, T.; Shi, F.; Xu, C. Structure Characterization and Anti-Inflammatory Activity of Polysaccharides from Lingzhi or Reishi Medicinal Mushroom Ganoderma lucidum (Agaricomycetes) by Microwave-Assisted Freeze-Thaw Extraction. Int. J. Med. Mushrooms 2022, 24, 49–61. [Google Scholar] [CrossRef]
- Li, Y.; Tang, J.; Gao, H.; Xu, Y.; Han, Y.; Shang, H.; Lu, Y.; Qin, C. Ganoderma lucidum Triterpenoids and Polysaccharides Attenuate Atherosclerotic Plaque in High-Fat Diet Rabbits. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 1929–1938. [Google Scholar] [CrossRef]
- Szekanecz, Z.; Besenyei, T.; Szentpétery, A.; Koch, A.E. Angiogenesis and Vasculogenesis in Rheumatoid Arthritis. Curr. Opin. Rheumatol. 2010, 22, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.C.; Sivakumar, B. Hypoxia and Angiogenesis in Rheumatoid Arthritis. Curr. Opin. Rheumatol. 2005, 17, 293–298. [Google Scholar] [CrossRef]
- Leblond, A.; Allanore, Y.; Avouac, J. Targeting Synovial Neoangiogenesis in Rheumatoid Arthritis. Autoimmun. Rev. 2017, 16, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Y.; Yang, W.B.; Wong, C.H.; Shih, D.T. Effect of Reishi Polysaccharides on Human Stem/Progenitor Cells. Bioorg. Med. Chem. 2010, 18, 8583–8591. [Google Scholar] [CrossRef] [PubMed]
- Miura, T.; Yuan, L.; Sun, B.; Fujii, H.; Yoshida, M.; Wakame, K.; Kosuna, K. Isoflavone Aglycon Produced by Culture of Soybean Extracts with Basidiomycetes and Its Anti-Angiogenic Activity. Biosci. Biotechnol. Biochem. 2002, 66, 2626–2631. [Google Scholar] [CrossRef] [Green Version]
- Cao, Q.Z.; Lin, Z.B. Antitumor and Anti-Angiogenic Activity of Ganoderma lucidum Polysaccharides Peptide. Acta Pharmacol. Sin. 2004, 25, 833–838. [Google Scholar]
- Cao, Q.; Lin, Z. Ganoderma lucidum Polysaccharides Peptide Inhibits the Growth of Vascular Endothelial Cell and the Induction of Vegf in Human Lung Cancer Cell. Life Sci. 2006, 78, 1457–1463. [Google Scholar] [CrossRef]
- Sun, L.X.; Lin, Z.B.; Duan, X.S.; Qi, H.H.; Yang, N.; Li, M.; Xing, E.H.; Sun, Y.; Yu, M.; Li, W.D.; et al. Suppression of the Production of Transforming Growth Factor Β1, Interleukin-10, and Vascular Endothelial Growth Factor in the B16f10 Cells by Ganoderma lucidum Polysaccharides. J. Interferon Cytokine Res. 2014, 34, 667–675. [Google Scholar] [CrossRef]
- Lin, C.Y.; Chen, Y.H.; Lin, C.Y.; Hsu, H.Y.; Wang, S.H.; Liang, C.J.; Kuan, P.; Wu, J.; Pai, P.Y.; Wu, C.C.; et al. Ganoderma lucidum Polysaccharides Attenuate Endotoxin-Induced Intercellular Cell Adhesion Molecule-1 Expression in Cultured Smooth Muscle Cells and in the Neointima in Mice. J. Agric. Food Chem. 2010, 58, 9563–9571. [Google Scholar] [CrossRef]
- Wang, S.H.; Liang, C.J.; Weng, Y.W.; Chen, Y.H.; Hsu, H.Y.; Chien, H.F.; Tsai, J.S.; Tseng, Y.C.; Li, C.Y.; Chen, Y.L. Ganoderma lucidum Polysaccharides Prevent Platelet-Derived Growth Factor-Stimulated Smooth Muscle Cell Proliferation In Vitro and Neointimal Hyperplasia in the Endothelial-Denuded Artery in Vivo. J. Cell Physiol. 2012, 227, 3063–3071. [Google Scholar] [CrossRef]
- Nygaard, G.; Firestein, G.S. Restoring Synovial Homeostasis in Rheumatoid Arthritis by Targeting Fibroblast-Like Synoviocytes. Nat. Rev. Rheumatol. 2020, 16, 316–333. [Google Scholar] [CrossRef]
- Maeda, K.; Yoshida, K.; Nishizawa, T.; Otani, K.; Yamashita, Y.; Okabe, H.; Hadano, Y.; Kayama, T.; Kurosaka, D.; Saito, M. Inflammation and Bone Metabolism in Rheumatoid Arthritis: Molecular Mechanisms of Joint Destruction and Pharmacological Treatments. Int. J. Mol. Sci. 2022, 23, 2871. [Google Scholar] [CrossRef]
- Li, E.K.; Tam, L.S.; Wong, C.K.; Li, W.C.; Lam, C.W.; Wachtel-Galor, I.; Benzie, F.; Bao, Y.X.; Leung, P.; Tomlinson, C. Safety and Efficacy of Ganoderma lucidum (Lingzhi) and San Miao San Supplementation in Patients with Rheumatoid Arthritis: A Double-Blind, Randomized, Placebo-Controlled Pilot Trial. Arthritis Rheum. 2007, 57, 1143–1150. [Google Scholar] [CrossRef]
- Tran, P.T.; Dat, N.T.; Dang, N.H.; Van Cuong, P.; Lee, S.; Hwangbo, C.; Van Minh, C.; Lee, J.H. Ganomycin I from Ganoderma lucidum Attenuates Rankl-Mediated Osteoclastogenesis by Inhibiting Mapks and Nfatc1. Phytomedicine 2019, 55, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Laçin, N.; İzol, S.B.; İpek, F.; Tuncer, M.C. Ganoderma lucidum, a Promising Agent Possessing Antioxidant and Anti-Inflammatory Effects for Treating Calvarial Defects with Graft Application in Rats. Acta Cir. Bras. 2019, 34, e201900904. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Lin, Z. Anti-Aging Effect of Ganoderma (Lingzhi) with Health and Fitness. Adv. Exp. Med. Biol. 2019, 1182, 299–309. [Google Scholar]
- Klein, A.; Molad, Y. Hematological Manifestations among Patients with Rheumatic Diseases. Acta Haematol. 2021, 144, 403–412. [Google Scholar] [CrossRef]
- Vreugdenhil, G.; Swaak, A.J. Anaemia in Rheumatoid Arthritis: Pathogenesis, Diagnosis and Treatment. Rheumatol. Int. 1990, 9, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Masson, C. Rheumatoid Anemia. Jt. Bone Spine 2011, 78, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Tang, M.; Xie, X.; Yang, Y.; Li, F. Ginsenoside Compound K- a Potential Drug for Rheumatoid Arthritis. Pharmacol. Res. 2021, 166, 105498. [Google Scholar] [CrossRef]
- Zhu, X.L.; Liu, J.H.; Li, W.D.; Lin, Z.B. Promotion of Myelopoiesis in Myelosuppressed Mice by Ganoderma lucidum Polysaccharides. Front. Pharmacol. 2012, 3, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emam, H.E.; Ahmed, H.B. Polysaccharides Templates for Assembly of Nanosilver. Carbohydr. Polym. 2016, 135, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Zhao, J.; Li, Y.; Zhu, L.; Jin, M.; Wang, L.; Liu, J.; Lei, J.; Li, Z. Self-Assembled Ph-Sensitive Nanoparticles Based on Ganoderma lucidum Polysaccharide-Methotrexate Conjugates for the Co-Delivery of Anti-Tumor Drugs. ACS Biomater. Sci. Eng. 2021, 7, 3764–3773. [Google Scholar] [CrossRef]
- Lee, S.Y.; Rhee, H.M. Cardiovascular Effects of Mycelium Extract of Ganoderma lucidum: Inhibition of Sympathetic Outflow as a Mechanism of Its Hypotensive Action. Chem. Pharm. Bull. (Tokyo) 1990, 38, 1359–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, H.B.; Yamamoto, A.; Matsumoto, S.; Ito, H.; Igami, K.; Miyazaki, T.; Kondo, R.; Shimizu, K. Hypotensive Effects and Angiotensin-Converting Enzyme Inhibitory Peptides of Reishi (Ganoderma lingzhi) Auto-Digested Extract. Molecules 2014, 19, 13473–13485. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.Y.; Eo, S.K.; Kim, Y.S.; Lee, C.K.; Han, S.S. Antimicrobial Activity of Ganoderma lucidum Extract Alone and in Combination with Some Antibiotics. Arch. Pharm. Res. 1994, 17, 438–442. [Google Scholar] [CrossRef]
- Karwa, A.; Gaikwad, S.; Rai, M.K. Mycosynthesis of Silver Nanoparticles Using Lingzhi or Reishi Medicinal Mushroom, Ganoderma lucidum (W. Curt.:Fr.) P. Karst. And Their Role as Antimicrobials and Antibiotic Activity Enhancers. Int. J. Med. Mushrooms 2011, 13, 483–491. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Years | Authors | Doses of GLP | Experimental Model/Cell Type | Effects of GLP |
|---|---|---|---|---|
| 2004 | Cao et al. [96] | 1, 10 and 100 mg/L | HUVECs | GLPP inhibits angiogenesis by directly inhibiting the cell proliferation of HUVEC. |
| 2006 | Cao et al. [97] | 10 and 100 μg/mL | HUVECs | GLPP induced cell death in HUVEC by decreasing the expression of Bcl-2 and increasing the expression of Bax, which reduced VEGF secretion under hypoxic conditions. |
| 2007 | Ho et al. [33] | 50, 125 and 250 μg/mL | RASF | GLPP significantly inhibited the proliferation of RASF and suppressed their production by inhibiting the NF-κB pathway. |
| 2010 | Lin et al. [99] | 0.25 and 0.5 μg/mL | HASMCs | GLP suppressed LPS-induced ICAM-1 mRNA and protein expression and reduced the binding of human monocytes to LPS-stimulated HASMCs. |
| 2012 | Wang et al. [100] | 100 mg/kg | male C57BL/6 J mice | GLP decreased the neointimal area in vivo. |
| 2014 | Wang et al. [53] | 5, 20, 50 and 100 μg/mL | RAW264.7 | GLP down-regulated the expression of iNOS and TNF-α and severely inhibited the phosphorylation of IκBα and JNK1/2. |
| 2014 | Zhong et al. [107] | 50 mg/kg | Male Kunming mice | GLP could increase antioxidant enzyme activities and decrease the MDA levels in the skeletal muscle of mice. |
| 2018 | Wei et al. [77] | 100 mg/kg | male C57BL/6 J mice | GLP decreased the secretion of proinflammatory cytokines, such as TNF-α, IL-6, IL-1β, and IL-17, and decreased the populations of Th17 cells and NK cells. |
| 2021 | Sang et al. [52] | 100 and 300 mg/kg | male C57BL/6 J mice | GLP inhibited macrophage infiltration and suppressed MAPK (JNK and ERK) activation. |
| 2021 | Guo et al. [59] | 0.2, 0.4, 0.8 and 1.6 mg/mL | RAW264.7 | GLP inhibited LPS-induced inflammation markers and MAPK (JNK and ERK) activation in macrophage RAW264.7. |
| 2022 | Fang et al. [42] | 0.625, 1.25 and 2.5 mg/mL | RAW264.7 | GLP inhibited the activation of macrophage RAW264.7 and the expression of the inflammatory mediators IL-1β, TNF-α, iNOS, and COX-2. |
| 2022 | Jia et al. [89] | 1, 10, and 100 μg/mL | RAW264.7 | GLP can inhibit the production of NO, TNF-α, IL-1β, and IL-6 in RAW 264.7 macrophage stimulated by LPS. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, M.; Yao, J.; Zhang, Y.; Sun, H.; Liu, M. Potential Anti-Rheumatoid Arthritis Activities and Mechanisms of Ganoderma lucidum Polysaccharides. Molecules 2023, 28, 2483. https://doi.org/10.3390/molecules28062483
Meng M, Yao J, Zhang Y, Sun H, Liu M. Potential Anti-Rheumatoid Arthritis Activities and Mechanisms of Ganoderma lucidum Polysaccharides. Molecules. 2023; 28(6):2483. https://doi.org/10.3390/molecules28062483
Chicago/Turabian StyleMeng, Meng, Jialin Yao, Yukun Zhang, Huijun Sun, and Mozhen Liu. 2023. "Potential Anti-Rheumatoid Arthritis Activities and Mechanisms of Ganoderma lucidum Polysaccharides" Molecules 28, no. 6: 2483. https://doi.org/10.3390/molecules28062483
APA StyleMeng, M., Yao, J., Zhang, Y., Sun, H., & Liu, M. (2023). Potential Anti-Rheumatoid Arthritis Activities and Mechanisms of Ganoderma lucidum Polysaccharides. Molecules, 28(6), 2483. https://doi.org/10.3390/molecules28062483