

Structure-Based Profiling of Potential Phytomolecules with AKT1 a Key Cancer Drug Target


Abstract
:
1. Introduction
2. Results
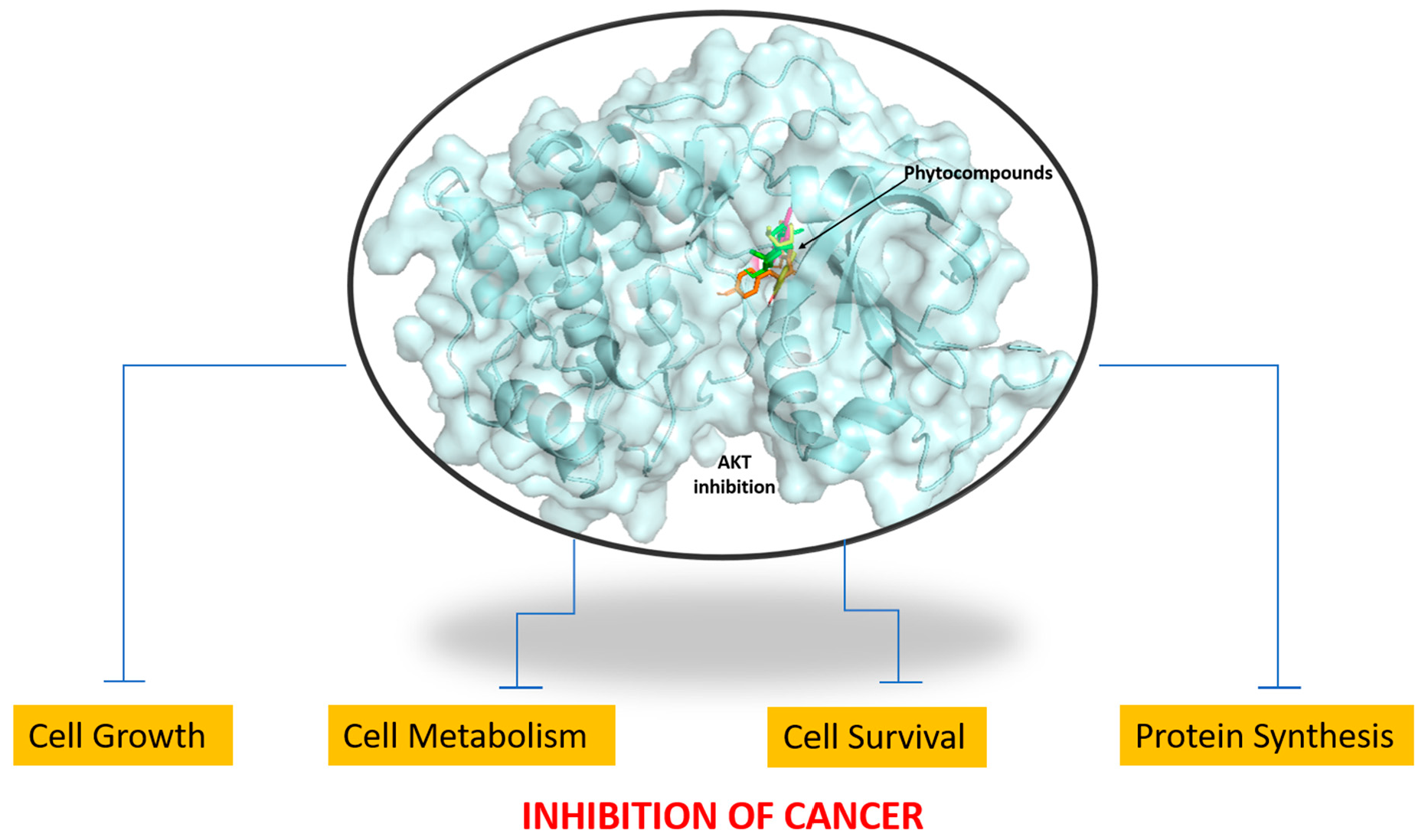
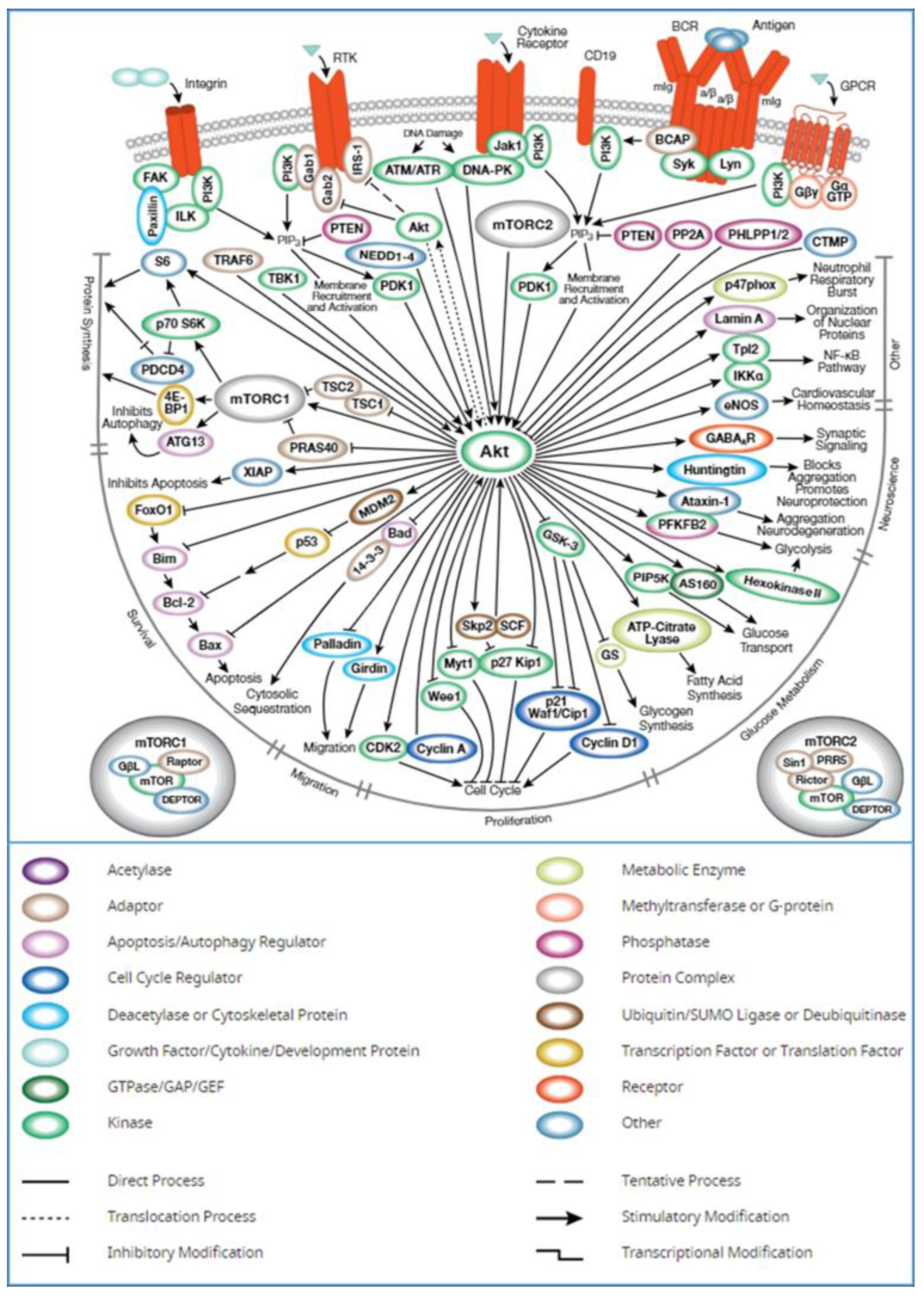
2.1. Identification of AKT1 as a Potential Biomarker
2.2. Protein Structure Analysis
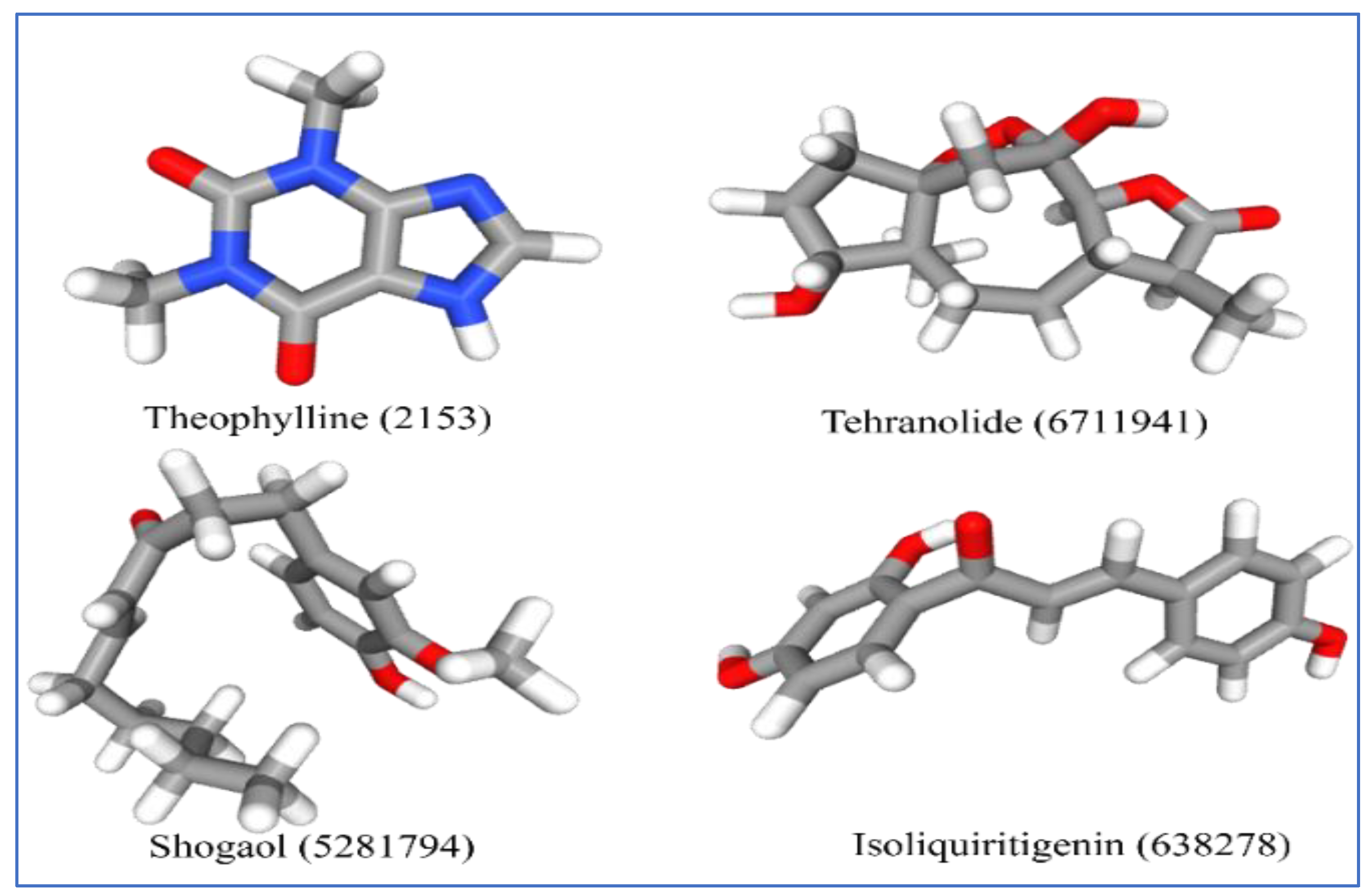
2.3. Ligand Selection and Preparation
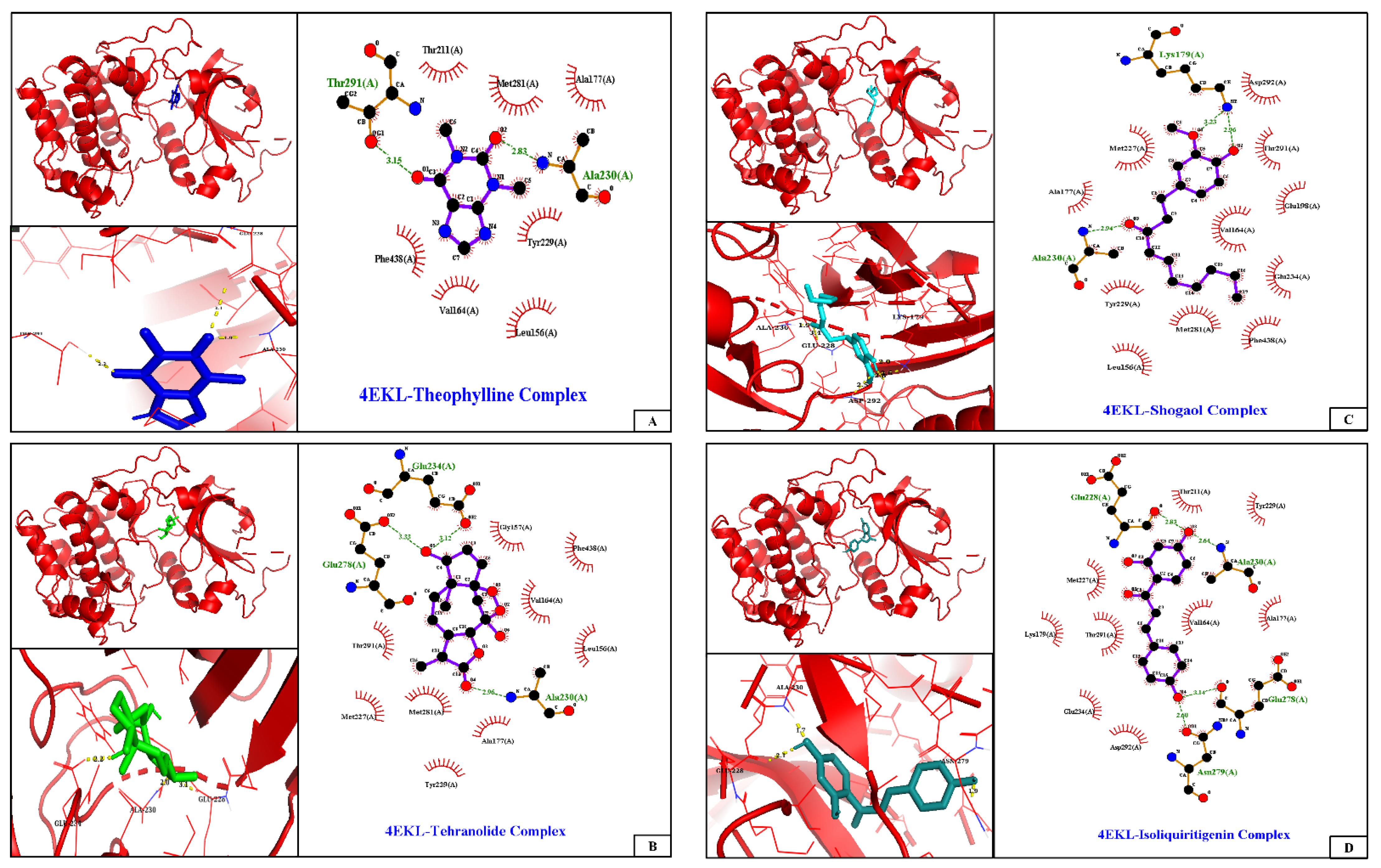
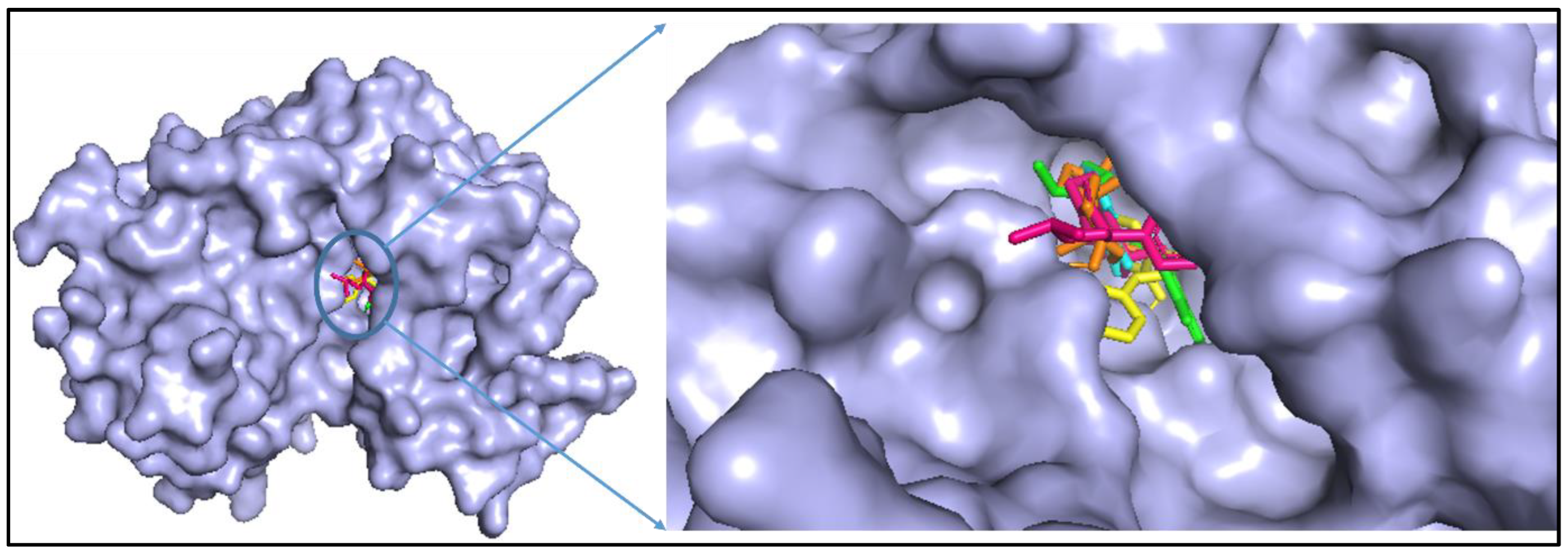
2.4. Molecular Docking and Analysis of the Complex
2.5. Analysis of ADMET Properties
3. Discussion
4. Materials and Methods
4.1. Identification of Biomarker
4.2. Selection and Preparation of Protein Target
4.3. Ligand Preparation
4.4. Molecular Docking and Protein–Ligand Interaction
4.5. Prediction of ADME and Toxicity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.-L.; Yu, Y.-C.; Hsia, S.-M. Perspectives on the Role of Isoliquiritigenin in Cancer. Cancers 2021, 13, 115. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.K.; Chakraborty, D.; Sarkar, I.; Khan, T.; Sa, G. New insights into therapeutic activity and anticancer properties of curcumin. J. Exp. Pharmacol. 2017, 9, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Karim, S.; Merdad, A.; Schulten, H.-J.; Jayapal, M.; Dallol, A.; Buhmeida, A.; Al-Thubaity, F.; Mirza, Z.; Gari, M.A.; Chaudhary, A.G.; et al. Low expression of leptin and its association with breast cancer: A transcriptomic study. Oncol. Rep. 2016, 36, 43–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karim, S.; Al-Maghrabi, J.A.; Farsi, H.M.A.; Al-Sayyad, A.J.; Schulten, H.-J.; Buhmeida, A.; Mirza, Z.; Al-Boogmi, A.A.; Ashgan, F.T.; Shabaad, M.M.; et al. Cyclin D1 as a therapeutic target of renal cell carcinoma-a combined transcriptomics, tissue microarray and molecular docking study from the Kingdom of Saudi Arabia. BMC Cancer 2016, 16, 741. [Google Scholar] [CrossRef] [Green Version]
- Carino, A.; Graziosi, L.; Marchianò, S.; Biagioli, M.; Marino, E.; Sepe, V.; Zampella, A.; Distrutti, E.; Donini, A.; Fiorucci, S. Analysis of Gastric Cancer Transcriptome Allows the Identification of Histotype Specific Molecular Signatures with Prognostic Potential. Front. Oncol. 2021, 11, 1–17. [Google Scholar] [CrossRef]
- Karim, S.; Mirza, Z.; Chaudhary, A.G.; Abuzenadah, A.M.; Gari, M.; Al-Qahtani, M.H. Assessment of Radiation Induced Therapeutic Effect and Cytotoxicity in Cancer Patients Based on Transcriptomic Profiling. Int. J. Mol. Sci. 2016, 17, 250. [Google Scholar] [CrossRef] [Green Version]
- Hua, H.; Zhang, H.; Chen, J.; Wang, J.; Liu, J.; Jiang, Y. Targeting Akt in cancer for precision therapy. J. Hematol. Oncol. 2021, 14, 128. [Google Scholar] [CrossRef]
- Bellantoni, A.; Wagner, L. Pursuing Precision: Receptor Tyrosine Kinase Inhibitors for Treatment of Pediatric Solid Tumors. Cancers 2021, 13, 3531. [Google Scholar] [CrossRef]
- Samra, B.; Jabbour, E.; Ravandi, F.; Kantarjian, H.; Short, N.J. Evolving therapy of adult acute lymphoblastic leukemia: State-of-the-art treatment and future directions. J. Hematol. Oncol. 2020, 13, 70. [Google Scholar] [CrossRef]
- Nasrazadani, A.; Thomas, R.A.; Oesterreich, S.; Lee, A.V. Precision Medicine in Hormone Receptor-Positive Breast Cancer. Front. Oncol. 2018, 8, 144. [Google Scholar] [CrossRef] [Green Version]
- Cheung, M.; Testa, J.R. Diverse mechanisms of AKT pathway activation in human malignancy. Curr. Cancer Drug Targets 2013, 13, 234–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellacosa, A.; Kumar, C.C.; Di Cristofano, A.; Testa, J.R. Activation of AKT Kinases in Cancer: Implications for Therapeutic Targeting. Adv. Cancer Res. 2005, 94, 29–86. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.M. Proteus syndrome review: Molecular, clinical, and pathologic features. Clin. Genet. 2013, 85, 111–119. [Google Scholar] [CrossRef]
- Moghtaderi, H.; Sepehri, H.; Delphi, L.; Attari, F. Gallic acid and curcumin induce cytotoxicity and apoptosis in human breast cancer cell MDA-MB-231. Bioimpacts 2018, 8, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Vladu, A.F.; Ficai, D.; Ene, A.G.; Ficai, A. Combination Therapy Using Polyphenols: An Efficient Way to Improve Antitumoral Activity and Reduce Resistance. Int. J. Mol. Sci. 2022, 23, 10244. [Google Scholar] [CrossRef]
- Bakrim, S.; El Omari, N.; El Hachlafi, N.; Bakri, Y.; Lee, L.-H.; Bouyahya, A. Dietary Phenolic Compounds as Anticancer Natural Drugs: Recent Update on Molecular Mechanisms and Clinical Trials. Foods 2022, 11, 3323. [Google Scholar] [CrossRef]
- Ali, R.; Mirza, Z.; Ashraf, G.M.D.; Kamal, M.A.; Ansari, S.A.; Damanhouri, G.A.; Abuzenadah, A.M.; Chaudhary, A.G.; Sheikh, I.A. New anticancer agents: Recent developments in tumor therapy. Anticancer Res. 2012, 32, 2999. [Google Scholar]
- Smith, U.; Carvalho, E.; Mosialou, E.; Beguinot, F.; Formisano, P.; Rondinone, C. PKB Inhibition Prevents the Stimulatory Effect of Insulin on Glucose Transport and Protein Translocation but Not the Antilipolytic Effect in Rat Adipocytes. Biochem. Biophys. Res. Commun. 2000, 268, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Reuveni, H.; Livnah, N.; Geiger, T.; Klein, S.; Ohne, O.; Cohen, I.; Benhar, M.; Gellerman, G.; Levitzki, A. Toward a PKB Inhibitor: Modification of a Selective PKA Inhibitor by Rational Design. Biochemistry 2002, 41, 10304–10314. [Google Scholar] [CrossRef]
- Kozikowski, A.P.; Sun, H.; Brognard, J.; Dennis, P.A. Novel PI Analogues Selectively Block Activation of the Pro-survival Serine/Threonine Kinase Akt. J. Am. Chem. Soc. 2003, 125, 1144–1145. [Google Scholar] [CrossRef]
- Van Ummersen, L.; Binger, K.; Volkman, J.; Marnocha, R.; Tutsch, K.; Kolesar, J.; Arzoomanian, R.; Alberti, D.; Wilding, G. A Phase I Trial of Perifosine (NSC 639966) on a Loading Dose/Maintenance Dose Schedule in Patients with Advanced Cancer. Clin. Cancer Res. 2004, 10, 7450–7456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambardella, V.; Tarazona, N.; Cejalvo, J.M.; Lombardi, P.; Huerta, M.; Roselló, S.; Fleitas, T.; Roda, D.; Cervantes, A. Personalized Medicine: Recent Progress in Cancer Therapy. Cancers 2020, 12, 1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prajapati, R.; Seong, S.H.; Park, S.E.; Paudel, P.; Jung, H.A.; Choi, J.S. Isoliquiritigenin, a potent human monoamine oxidase inhibitor, modulates dopamine D1, D3, and vasopressin V1A receptors. Sci. Rep. 2021, 11, 23528. [Google Scholar] [CrossRef]
- Han, Y.J.; Kang, B.; Yang, E.-J.; Choi, M.-K.; Song, I.-S. Simultaneous Determination and Pharmacokinetic Characterization of Glycyrrhizin, Isoliquiritigenin, Liquiritigenin, and Liquiritin in Rat Plasma Following Oral Administration of Glycyrrhizae Radix Extract. Molecules 2019, 24, 1816. [Google Scholar] [CrossRef] [Green Version]
- Peng, F.; Du, Q.; Peng, C.; Wang, N.; Tang, H.; Xie, X.; Shen, J.; Chen, J. A Review: The Pharmacology of Isoliquiritigenin. Phytother. Res. 2015, 29, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.K.; Na, H.-K.; Surh, Y.-J. Ginger-Derived Phenolic Substances with Cancer Preventive and Therapeutic Potential. Forum Nutr. 2009, 61, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.H.; Blunden, G.; Tanira, M.O.; Nemmar, A. Some phytochemical, pharmacological and toxicological properties of ginger (Zingiber officinale Roscoe): A review of recent research. Food Chem. Toxicol. 2008, 46, 409–420. [Google Scholar] [CrossRef]
- Pan, M.-H.; Hsieh, M.-C.; Kuo, J.-M.; Lai, C.-S.; Wu, H.; Sang, S.; Ho, C.-T. 6-Shogaol induces apoptosis in human colorectal carcinoma cellsviaROS production, caspase activation, and GADD 153 expression. Mol. Nutr. Food Res. 2008, 52, 527–537. [Google Scholar] [CrossRef]
- Clark, I.M.; Swingler, T.E.; Sampieri, C.L.; Edwards, D. The regulation of matrix metalloproteinases and their inhibitors. Int. J. Biochem. Cell Biol. 2008, 40, 1362–1378. [Google Scholar] [CrossRef] [Green Version]
- Overall, C.M.; López-Otín, C. Strategies for MMP inhibition in cancer: Innovations for the post-trial era. Nat. Rev. Cancer 2002, 2, 657–672. [Google Scholar] [CrossRef] [PubMed]
- Bogenrieder, T.; Herlyn, M. Axis of evil: Molecular mechanisms of cancer metastasis. Oncogene 2003, 22, 6524–6536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadwick, M.; Trewin, H.; Gawthrop, F.; Wagstaff, C. Sesquiterpenoids Lactones: Benefits to Plants and People. Int. J. Mol. Sci. 2013, 14, 12780–12805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noori, S.; Hassan, Z.M. Tehranolide inhibits cell proliferation via calmodulin inhibition, PDE, and PKA activation. Tumor Biol. 2013, 35, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Noori, S.; Hassan, Z.M. Tehranolide inhibits proliferation of MCF-7 human breast cancer cells by inducing G0/G1 arrest and apoptosis. Free Radic. Biol. Med. 2012, 52, 1987–1999. [Google Scholar] [CrossRef] [PubMed]
- Noori, S.; Taghikhani, M.; Hassan, Z.M.; Allameha, A.; Mostafaei, A. Tehranolide molecule modulates the immune response, reduce regulatory T cell and inhibits tumor growth in vivo. Mol. Immunol. 2010, 47, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, M.; Shah, I.; Ali, N.; Adhikari, A.; Tahir, M.N.; Shah, S.W.A.; Ishtiaq, S.; Khan, J.; Khan, S.; Umer, M.N. Sesquiterpene lactone! a promising antioxidant, anticancer and moderate antinociceptive agent from Artemisia macrocephala jacquem. BMC Complement. Altern. Med. 2017, 17, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Song, J.; Zhou, Y.; Cao, L.; Gong, Y.; Wei, Y.; Yang, H.; Tang, L. Methylxanthine derivatives promote autophagy in gastric cancer cells targeting PTEN. Anti-Cancer Drugs 2019, 30, 347–355. [Google Scholar] [CrossRef]
- Cornejo, M.G.; Mabialah, V.; Sykes, S.M.; Khandan, T.; Celso, C.L.; Lopez, C.K.; Rivera-Muñoz, P.; Rameau, P.; Tothova, Z.; Aster, J.C.; et al. Crosstalk between NOTCH and AKT signaling during murine megakaryocyte lineage specification. Blood 2011, 118, 1264–1273. [Google Scholar] [CrossRef]
- Trotman, L.C.; Alimonti, A.; Scaglioni, P.P.; Koutcher, J.A.; Cordon-Cardo, C.; Pandolfi, P.P. Identification of a tumour suppressor network opposing nuclear Akt function. Nature 2006, 441, 523–527. [Google Scholar] [CrossRef] [Green Version]
- Hollander, M.C.; Blumenthal, G.M.; Dennis, P.A. PTEN loss in the continuum of common cancers, rare syndromes and mouse models. Nat. Rev. Cancer 2011, 11, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.-T.; Yang, Z.; Lu, N.-H. Roles of PTEN (Phosphatase and Tensin Homolog) in Gastric Cancer Development and Progression. Asian Pac. J. Cancer Prev. 2014, 15, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Alers, S.; Löffler, A.S.; Wesselborg, S.; Stork, B. Role of AMPK-mTOR-Ulk1/2 in the Regulation of Autophagy: Cross Talk, Shortcuts, and Feedbacks. Mol. Cell. Biol. 2012, 32, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular Docking and Structure-Based Drug Design Strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, P.H.M.; Sodero, A.C.R.; Jofily, P.; Silva, F.P., Jr. Key Topics in Molecular Docking for Drug Design. Int. J. Mol. Sci. 2019, 20, 4574. [Google Scholar] [CrossRef] [Green Version]
- Carracedo-Reboredo, P.; Liñares-Blanco, J.; Rodríguez-Fernández, N.; Cedrón, F.; Novoa, F.J.; Carballal, A.; Maojo, V.; Pazos, A.; Fernandez-Lozano, C. A review on machine learning approaches and trends in drug discovery. Comput. Struct. Biotechnol. J. 2021, 19, 4538–4558. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Srivastava, D.; Sahu, M.; Tiwari, S.; Ambasta, R.K.; Kumar, P. Artificial intelligence to deep learning: Machine intelligence approach for drug discovery. Mol. Divers. 2021, 25, 1315–1360. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Abbasi, B.A.; Batool, R.; Mahmood, T.; Ali, B.; Khalil, A.T.; Kanwal, S.; Shah, S.A.; Ahmad, R. Potential phytocompounds for developing breast cancer therapeutics: Nature’s healing touch. Eur. J. Pharmacol. 2018, 827, 125–148. [Google Scholar] [CrossRef]
- Song, M.; Bode, A.M.; Dong, Z.; Lee, M.-H. AKT as a Therapeutic Target for Cancer. Cancer Res. 2019, 79, 1019–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foukas, L.C.; Daniele, N.; Ktori, C.; Anderson, K.; Jensen, J.; Shepherd, P.R. Direct Effects of Caffeine and Theophylline on p110δ and Other Phosphoinositide 3-Kinases. J. Biol. Chem. 2002, 277, 37124–37130. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Mao, L.; Xie, L.; Zhang, R.; Liu, Y.; Peng, L.; Yang, J.; Li, Q.; Yuan, M. Discovery of a Series of Theophylline Derivatives Containing 1,2,3-Triazole for Treatment of Non-Small Cell Lung Cancer. Front. Pharmacol. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Chang, Y.-L.; Hsu, Y.-J.; Chen, Y.; Wang, Y.-W.; Huang, S.-M. Theophylline exhibits anti-cancer activity via suppressing SRSF3 in cervical and breast cancer cell lines. Oncotarget 2017, 8, 101461–101474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, H.; Yang, H.; Tan, S.-H.; Chui, W.-K.; Chew, E.-H. 6-Shogaol, an active constituent of ginger, inhibits breast cancer cell invasion by reducing matrix metalloproteinase-9 expression via blockade of nuclear factor-κB activation. Br. J. Pharmacol. 2010, 161, 1763–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.O.; Lee, M.-H.; Oi, N.; Kim, S.-H.; Bae, K.B.; Huang, Z.; Kim, D.J.; Reddy, K.; Lee, S.-Y.; Park, S.J.; et al. [6]-Shogaol inhibits growth and induces apoptosis of non-small cell lung cancer cells by directly regulating Akt1/2. Carcinogenesis 2013, 35, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, C.-J.; Chou, C.-P.; Ho, C.-T.; Yen, G.-C. Molecular mechanism inhibiting human hepatocarcinoma cell invasion by 6-shogaol and 6-gingerol. Mol. Nutr. Food Res. 2012, 56, 1304–1314. [Google Scholar] [CrossRef]
- Wang, C.; Chen, Y.; Wang, Y.; Liu, X.; Liu, Y.; Li, Y.; Chen, H.; Fan, C.; Wu, D.; Yang, J. Inhibition of COX-2, mPGES-1 and CYP4A by isoliquiritigenin blocks the angiogenic Akt signaling in glioma through ceRNA effect of miR-194-5p and lncRNA NEAT1. J. Exp. Clin. Cancer Res. 2019, 38, 371. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Liu, C.; Zeng, W.-C.; Xu, G.-Y.; Wu, J.-M.; Li, Z.-W.; Huang, X.-Y.; Lin, R.-J.; Shi, X. Isoliquiritigenin inhibits the proliferation, migration and metastasis of Hep3B cells via suppressing cyclin D1 and PI3K/AKT pathway. Biosci. Rep. 2020, 40, BSR20192727. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Chen, H.; Luo, X.; An, B.; Wu, W.; Cao, S.; Ruan, S.; Wang, Z.; Weng, L.; Zhu, H.; et al. Isoliquiritigenin suppresses human melanoma growth by targeting miR-301b/LRIG1 signaling. J. Exp. Clin. Cancer Res. 2018, 37, 184. [Google Scholar] [CrossRef] [PubMed]
- Noori, S.; Taghikhani, M.; Hassan, Z.M.; Allameh, A.; Mostafaei, A. Tehranolide could shift the immune response towards Th1 and modulate the intra-tumor infiltrated T regulatory cells. Iran. J. Immunol. 2009, 6, 216–224. [Google Scholar] [PubMed]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef]
- Uhlén, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, 2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fährrolfes, R.; Bietz, S.; Flachsenberg, F.; Meyder, A.; Nittinger, E.; Otto, T.; Volkamer, A.; Rarey, M. ProteinsPlus: A web portal for structure analysis of macromolecules. Nucleic Acids Res. 2017, 45, W337–W343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the scope of the protein–ligand interaction profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligands | Binding Energy (kcal/mol) | Inhibition Constant (Ki) | Interacting Residues | |
|---|---|---|---|---|
| H-Bonds (Distances) | Others | |||
| Tehranolide (6711941) | −9.22 | 173.21 nM | Ala230 (2.0 Å), Glu234 (2.2 Å), Glu278 (3.33 Å) | Leu156, Gly157, Val164, Ala177, Met227, Tyr229, Met281, Thr291, Phe438 |
| Shogaol (5281794) | −8.19 | 992.48 nM | Lys179 (2.96), Glu228 (3.4 Å), Ala230 (1.9 Å) | Leu156, Val164, Ala177, Glu198, Met227, Tyr229, Glu234, Met281, Thr291, Asp292, Phe438 |
| Isoliquiritigenin (638278) | −8.29 | 841.68 nM | Glu228 (2.1 Å), Ala230 (1.7 Å), Glu278 (3.14 Å), Asn279 (1.9 Å), Thr291 (2.1 Å) | Val164, Ala177, Lys179, Thr211, Met227, Tyr229, Glu234, Asp292 |
| Theophylline (2153) | −7.26 | 4.73 µM | Glu228 (3.1 Å), Ala230 (1.9 Å), Thr291 (2.2 Å) | Leu156, Val164, Ala177, Thr211, Tyr229, Met281, Phe438 |
| Compound Property | Theophylline | Tehranolide | Shogaol | Isoliquiritigenin |
|---|---|---|---|---|
| cLogP | −0.31 | 1.18 | 4.33 | 2.27 |
| Solubility | −1.48 | −2.81 | −3.42 | −2.95 |
| Molecular weight | 180.0 | 298.33 | 276.0 | 256.0 |
| Drug-likeness | 2.51 | 0.71 | −14.4 | 0.76 |
| Mutagenic | High | None | High | High |
| Tumorigenic | High | High | None | None |
| Irritant | None | None | None | Medium |
| Reproductive effects | High | None | None | None |
| Drug-score | 0.2 | 0.45 | 0.22 | 0.359 |
| Compound Property | Isoliquiritigenin 638278 | Shogaol 5281794 | Tehranolide 6711941 | Theophylline 2153 |
|---|---|---|---|---|
| General Properties | ||||
| Structure |  |  |  |  |
| Bioavailability radar |  |  |  |  |
| Smiles | C1=CC(=CC=C1C=CC(=O)C2=C(C=C(C=C2)O)O)O | CCCCCC=CC(=O)CCC1=CC(=C(C=C1)O)OC | CC1C2CCC3(C(CCC34CC(C2OC1=O)(OO4)O)O)C | CN1C2=C(C(=O)N(C1=O)C)NC=N2 |
| Physiochemical Properties | ||||
| Formula | C15H12O4 | C17H24O3 | C15H22O6 | C7H8N4O2 |
| Molecular weight | 256.25 g/mol | 276.37 g/mol | 298.33 g/mol | 180.16 g/mol |
| XLogP3-AA | 0.9 | |||
| Rotatable bonds | 3 | 9 | 0 | 0 |
| H-bond acceptors | 4 | 3 | 6 | 3 |
| H-bond donors | 3 | 1 | 2 | 1 |
| Topological polar surface area | 77.76 Å2 | 46.53 Å2 | 85.22 Å2 | 72.68 Å2 |
| Lipophilicity (Consensus Log Po/w) | 2.37 | 3.76 | 1.20 | −0.19 |
| Water solubility | Soluble | Moderately soluble | Soluble | Soluble |
| Pharmacokinetics | ||||
| GI absorption | High | High | High | High |
| BBB permeant | Yes | Yes | No | No |
| P-gp substrate | No | No | Yes | No |
| CYP1A2 inhibitor | Yes | Yes | No | No |
| CYP2C19 inhibitor | No | Yes | No | No |
| CYP2C9 inhibitor | Yes | No | No | No |
| CYP2D6 inhibitor | No | Yes | No | No |
| CYP3A4 inhibitor | Yes | No | No | No |
| Log Kp (skin permeation) | −5.61 cm/s | −5.15 cm/s | −7.45 cm/s | −7.41 cm/s |
| Drug-likeness | ||||
| Lipinski | Yes; 0 violation | Yes; 0 violation | Yes; 0 violation | Yes; 0 violation |
| Ghose | Yes | Yes | Yes | No; 1 violation: WLOGP < −0.4 |
| Veber | Yes | Yes | Yes | Yes |
| Egan | Yes | Yes | Yes | Yes |
| Muegge | Yes | Yes | Yes | No; 1 violation: MW < 200 |
| Bioavailability score | 0.55 | 0.55 | 0.55 | 0.55 |
| Synthetic accessibility | 2.52 | 2.51 | 5.65 | 1.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirza, Z.; Karim, S. Structure-Based Profiling of Potential Phytomolecules with AKT1 a Key Cancer Drug Target. Molecules 2023, 28, 2597. https://doi.org/10.3390/molecules28062597
Mirza Z, Karim S. Structure-Based Profiling of Potential Phytomolecules with AKT1 a Key Cancer Drug Target. Molecules. 2023; 28(6):2597. https://doi.org/10.3390/molecules28062597
Chicago/Turabian StyleMirza, Zeenat, and Sajjad Karim. 2023. "Structure-Based Profiling of Potential Phytomolecules with AKT1 a Key Cancer Drug Target" Molecules 28, no. 6: 2597. https://doi.org/10.3390/molecules28062597
APA StyleMirza, Z., & Karim, S. (2023). Structure-Based Profiling of Potential Phytomolecules with AKT1 a Key Cancer Drug Target. Molecules, 28(6), 2597. https://doi.org/10.3390/molecules28062597