3.3. Alcohols
Alcohols, with their characteristic aromas, comprised the largest volatile group detected in 33 of the 35 studies reviewed. Alcohols are produced from carbohydrate degradation or amino acid catabolism [
52]. Across the 33 studies, the alcohols most commonly detected after fermentation were ethanol, 3-methyl-1-butanol (isoamyl alcohol/isopentyl alcohol), 2-methyl-1-butanol (amyl alcohol), 3-methyl-3-buten-1-ol (isoprenol), 2, 3-butanediol, 2-ethylhexanol, 1-hexanol, 2-hexen-1-ol, 3 hexen-1-ol, 2,6-dimethyl-4-heptanol, benzyl alcohol (phenyl methanol/benzene methanol), 2-phenylethyl alcohol (2-phenyl ethanol/benzene ethanol), 4-ethylphenol, 2-(4-methylphenyl)-2-propanol, 1-octanol, 1-octen-3-ol, (Z)-1,5-octadien-3-ol, octenol, 2-octenol, 1-nonanol, (Z)-3-nonen-1-ol, 2-undecanol, 3,7,11-trimethyl-1-dodecanol, and 2-tridecanol.
Ethanol is synthesised from sugars naturally present in plants; LAB utilise sugars via the phosphoketolase (PK) pathway, and yeast utilise sugars through the Embden–Meyerhof–Parnas (EMP) pathway [
13]. In mango slurry, fermentation involving the yeast
Saccharomyces cerevisiae (
S. cerevisiae) generated 30–100 times more ethanol than LAB fermentation [
41]. When
Williopsis saturnus var.
saturnus (
W. saturnus) yeast was combined with LAB for fermentation, there was also a marked increase in the ethanol concentration (six-fold increase) compared to LAB alone in durian pulp [
30]. However, ethanol can be generated by heterofermentative LAB, which possess the alcohol dehydrogenase enzyme that converts acetaldehyde into ethanol [
26]. In kiwifruit juice (cultivars of
Actinidia deliciosa cv. Xuxiang and
Actinidia chinensis cv. Hongyang), the ethanol concentration was 10,316.5, 17,249.2, and 8652.7 ng/mL in Xuxiang cultivar juice fermented by either
L. acidophilus,
L. helveticus, or
L. plantarum, respectively, compared to 6242.9 ng/mL in the unfermented juice. However, the ethanol concentration was 13,042.5, 7004.2, and 9551.9 ng/mL in Hongyang cultivar juice fermented by either
L. acidophilus,
L. helveticus, or
L. plantarum, respectively, compared to 19,642.6 ng/mL in the unfermented juice [
49]. The different ethanol concentrations produced from the two kiwifruit cultivars after LAB fermentation could be a result of the different substrate compositions of the cultivars, which are subjected to various metabolic pathways by LAB. After fermentation of orange pomace by
L. rhamnosus, 0.3 µg/mL of ethanol was detected in a distillate prepared using vacuum distillation to extract VOCs. However, ethanol was not detected in a fermented orange pomace distillate prepared using a simple distillation method. In the same study, the ethanol concentration detected in distillates prepared from
L. rhamnosus fermented melon by-product using vacuum distillation was 6.5 µg/mL, compared to 1.3 µg/mL in the distillate from the unfermented melon by-products, while ethanol was not detected in distillates prepared from the same samples using simple distillation [
44]. In papaya juice, the ethanol concentration was significantly (
p < 0.05) increased by 7 and 11 times after fermentation by either
L. plantarum or
L. acidophilus, respectively, compared to the concentration in the unfermented juice [
23]. On the other hand, only small changes in ethanol were observed in two studies: (1) Ricci et al. [
29] found that LAB fermented cherry juice had an ethanol concentration of 4.1–8.5 ng/mL, compared to 3.1–3.7 ng/mL in the unfermented juice, and (2) in watermelon juice fermented by either
L. rhamnosus, L. plantarum,
L. casei, or
Pediococcus pentosaceus (
P. pentosaceus), the ethanol concentration was 16.8, 15.2, 15.1, and 15 ng/mL, respectively, compared to 14.6 ng/mL in the unfermented juice; however, after fermentation by
Levilactobacillus brevis (
L. brevis), the ethanol concentration was 13.9 ng/mL [
48]. Six studies reported that LAB fermentation reduced the ethanol concentration in fermented fruit and vegetable juices, compared to the unfermented juices, with the synthesis of various esters speculated to have caused this decrease: (1) In unfermented Chinese wolfberry juice, the ethanol concentration was 5501.3 µg/mL, compared to 1364.7 µg/mL in the
L. acidophilus fermented juice, where it was not detected in the juice fermented by other LAB strains [
50]; (2) in two varieties of unfermented jujube (Muzao, and Hetain) juices, the ethanol concentration was 6850, and 6130 ng/mL, respectively, compared to 5740, 5100, and 1530 ng/mL in Muzao fermented with either
L. helveticus,
L. casei, or
L. plantarum, respectively, and 5380, 4400, 2660, and 2410 ng/mL in Hetain fermented with either
L. casei, L. acidophilus, L. plantarum, or
L. helveticus, respectively [
20]; (3) in okara, the initial ethanol concentration was 44 µg/g which reduced to 20.4 and 13.8 µg/g after fermentation with LAB monoculture (
L. rhamnosus or
Pediococcus acidilactic (
P. acidilactic), respectively) and to 19.6 µg/g after co-culture fermentation (
L. acidophilus,
L. rhamnosus, and
P. acidilactic). However, in okara fermented with an
L. acidophilus monoculture, the ethanol concentration increased from 44 to 57.4 µg/g [
46]; (4) in unfermented apple juice, the ethanol concentration was 188.4 ng/mL, compared to 83.4–123.5 ng/mL after fermentation with various LAB strains [
19]; (5) in non-pH-adjusted (2.7) sea buckthorn juice, the ethanol concentration was 170.7 ng/mL, which reduced after fermentation for 36 and 72 h by
L. plantarum to 165.4 and 152 ng/mL, respectively. However, if the pH of the juice was adjusted to pH 3.5, the initial ethanol concentration of 166.3 ng/mL increased after
L. plantarum fermentation for 36 and 72 h to 194.6 and 206.5 ng/mL, respectively [
42]; and (6) in tomato juice, the ethanol concentration after fermentation with either
L. plantarum or
L. casei was 2.7 and 1.2 times lower, respectively, compared to its concentration in the unfermented juice [
32].
1-Octanol is a fatty alcohol produced by micro-organisms utilizing glucose as a substrate through a fatty acid synthesis pathway using various enzymes [
53]. The 1-octanol concentration was generally reported to increase after LAB fermentation in the 5 studies reviewed: (1) In Chinese wolfberry juice fermented by either
L. plantarum,
L. casei,
Lacticaseibacillus paracasei (
L. paracasei),
L. acidophilus,
L. helveticus, or
Bifidobacterium Lactis (
B. lactis), the 1-octanol concentration was 172.9, 119.1, 137.2, 209.3, 131.4, and 142.4 µg/mL, where it was not detected in the unfermented juice [
50]; (2) the 1-octanol concentration in a distillate prepared using vacuum distillation from orange pomace fermented by
L. rhamnosus was 1.5 µg/mL, compared to 0.1 µg/mL in the distillate from unfermented pomace; in distillates prepared using a simple distillation method, the 1-octanol concentration in the fermented orange pomace distillate was 2.1 µg/mL, compared to 1.8 µg/mL in the unfermented pomace distillate. Interestingly in the same study, using the simple distillation method, the 1-octanol concentration in the fermented melon by-product distillate was 21.5 µg/mL, compared to 7.1 µg/mL in the unfermented by-product distillate, whereas in extracts prepared by vacuum distillation, the 1-octanol concentration in the distillate from melon by-product fermented by
L. rhamnosus was 1.1 µg/mL, compared to 0.2 µg/mL in the unfermented by-product distillate [
44]; (3) in kiwifruit juice (Xuxiang and Hongyang cultivars), the 1-octanol concentration was 285.5 and 325.5 ng/mL in Xuxiang cultivar juice fermented by either
L. helveticus or
L. plantarum, respectively, compared to 146.4 ng/mL in the unfermented juice, where it was not detected in Xuxiang cultivar juice fermented by
L. acidophilus. Interestingly, with the Hongyang cultivar juice, 1-octanol was not detected in the unfermented juice or in any of the LAB fermented juices. [
49]; (4) in cherry juice fermented by either
L. plantarum,
L. rhamnosus, or
L. paracasei, the 1-octanol concentration was 4.5–7.8, 8.4, and 5.2 ng/mL, respectively, compared to 3.4–3.6 ng/mL in the unfermented juice [
29]; and (5) in apple juice fermented by either
L. acidophilus,
L. rhamnosus,
L. casei or
L. plantarum, the 1-octanol concentration was 4.2, 3.5, 3.8, and 4.0 ng/g, respectively, compared to 1.0 ng/g in the unfermented juice [
21]. However, in okara fermented by LAB monoculture of either
L. rhamnosus,
P. acidilactic or co-culture (
L. acidophilus,
L. rhamnosus, and
P. acidilactic), the 1-octanol concentration was 11.7, 17.8, and 1.7 µg/g, respectively, compared to 30.0 µg/g in the unfermented okara, whereas okara fermented with
L. acidophilus had an 1-octanol concentration of 34.7 µg/g [
46].
1-Hexanol is produced from the enzymatic oxidation of the fatty acid linoleic acid [
54]. In 11 experiments, the 1-hexanol concentration increased after fermentation: (1) In Chinese wolfberry juice fermented by either
L. paracasei,
L. acidophilus, or
B. lactis, the 1-hexanol concentration was 566.3, 728.2, and 682.2 µg/mL, respectively, where it was not detected in the unfermented juice and the juice fermented by other LAB strains [
50]; (2) for kiwifruit juice (Xuxiang, and Hongyang cultivars), the 1-hexanol concentration in the Xuxiang cultivar juice fermented by either
L. acidophilus,
L. helveticus, or
L. plantarum was 11,239.2, 13,280.9, and 11,713.8 ng/mL, respectively, compared to 7054.8 ng/mL in the unfermented juice, where for the Hongyang cultivar juice fermented by either
L. acidophilus,
L. helveticus, or
L. plantarum, the 1-hexanol concentration was 12,313.5, 12,357.1, and 11,461.1 ng/mL, respectively, compared to 2462.9 ng/mL in the unfermented juice [
49]; (3) the 1-hexanol concentration in a distillate prepared using simple distillation of orange pomace fermented by
L. rhamnosus was 1.1 µg/mL, compared to 0.1 µg/mL in the unfermented orange pomace distillate. In the same study, using simple distillation, the 1-hexanol concentration in fermented melon by-product distillate was 21.2 µg/mL, compared to 10.2 µg/mL in the unfermented melon by-product distillate, whereas, when using vacuum distillation, the 1-hexanol concentration in fermented melon by-product distillate was 4.3 µg/mL, compared to 0.4 µg/mL in the unfermented melon by-product distillate [
44]; (4) in watermelon juice fermented by either
L. rhamnosus, L. casei, L. plantarum,
L. brevis, or
P. pentosaceus, the 1-hexanol concentration was 79.9, 121.0, 141.0, 170.0, and 171.0 ng/mL, respectively, compared to 74.7 ng/mL in the unfermented juice [
48]; (5) in cherry juice fermented by either
L. plantarum,
L. rhamnosus, or
L. paracasei, the 1-hexanol concentration was 1.9–3.5, 2.1, and 1.1 ng/mL, respectively, compared to 0.7–0.8 ng/mL in the unfermented juice [
29]; (6) in okara fermented by either
L. acidophilus, L. rhamnosus, or
P. acidilactic, the 1-hexanol concentration was 206.7, 217, and 217.5 µg/g, respectively, compared to 64.2 µg/g in the unfermented okara. However, for okara fermented with LAB co-cultures (
L. acidophilus,
L. rhamnosus, and
P. acidilactic), the 1-hexanol concentration reduced to 16.0 µg/g [
46]; (7) in jujube juice fermented by a mixture of
L. plantarum,
L. rhamnosus, and
S. thermophilus, the 1-hexanol concentration was 824 ng/g, and it was not detected in the unfermented juice [
47]; (8) in apple juice fermented by either
L. plantarum,
L. rhamnosus, L. acidophilus, or
L. casei, the 1-hexanol concentration was 21.1, 22.0, 47.4, and 52.4 ng/g, respectively, compared to 1.2 ng/g in the unfermented juice [
21]; (9) in grape juice fermented by LAB, the 1-hexanol concentration was 3.3-fold higher compared to the concentration in the unfermented juice [
43]; (10) in papaya juice fermented by either
L. acidophilus or
L. plantarum, the 1-hexanol concentration was 3 and 4 times higher, respectively, compared to its concentration in the unfermented juice [
23]; and (11) in
Momordica charantia juice fermented by
L. plantarum, the 1-hexanol concentration was 2.5 times higher compared to the concentration in the unfermented juice [
18]. However, the 1-hexanol concentration has also been reported to reduce after fermentation in 3 studies: (1) In goji juice fermented by different combinations of bacterial strains (either
L. plantarum,
L. rhamnosus,
Limosilactobacillus reuteri (
L. reuteri),
Bacillus velezensis (
B. velezensis), or
Bacillus licheniformis (
B. licheniformis)), the 1-hexanol concentration was 43.0–61.4 ng/g, compared to 80.2 ng/g in the unfermented juice [
25]; (2) in tomato juice fermented by
L. plantarum, the 1-hexanol concentration was reduced by half, compared to its concentration in the unfermented juice [
32]; and (3) in mung bean fermented by
L. plantarum, 1-hexanol was not detected, where it was detected in the unfermented bean [
45].
Benzyl alcohol is an aromatic alcohol produced by microbial fermentation either from glucose [
55] or the amino acid phenylalanine [
56]. Benzyl alcohol was detected after bacterial fermentation in 7 studies [
19,
25,
29,
34,
35,
45,
47] with concentrations ranging from 2.3 ng/mL to 270 ng/mL.
3-Methyl-1-butanol and 2-phenylethyl alcohol were the most common amino acid-derived alcohols detected after fermentation. 3-Methyl-1-butanol and 2-phenylethyl alcohol are synthesised from the catabolism of the amino acids leucine [
57] and phenylalanine [
58], respectively. 3-Methyl-1-butanol has a malt/alcoholic/whiskey odour that is considered unpleasant when present in concentrations greater than 400 µg/mL [
59]. In fermented fruit and vegetable juices, 3-methyl-1-butanol was detected in 10 studies [
21,
22,
23,
24,
29,
31,
38,
44,
45,
49]. However, in fermented fruit and vegetable juices, the 3-methyl-1-butanol concentration was reduced in 5 studies: (1) In Chinese wolfberry juice fermented by either
L. casei,
L. paracasei,
L. acidophilus,
L. helveticus, or
B. lactis, the 3-methyl-1-butanol concentration was 825.9, 674.0, 833.6, 799.6, and 820.6 µg/mL, respectively, compared to 2065.3 µg/mL in the unfermented juice, and it was not detected in the juice fermented by
L. plantarum [
50]; (2) in non-pH-adjusted (2.7) sea buckthorn juice, the 3-methyl-1-butanol concentration was 122.9 ng/mL, which was reduced after fermentation for 36 and 72 h by
L. plantarum to 98.9 and 91.4 ng/mL, respectively. However, if the pH of the juice was adjusted to pH 3.5, the initial 3-methyl-1-butanol concentration of 121.4 ng/mL increased after
L. plantarum fermentation for 36 and 72 h to 217.9 and 233 ng/mL, respectively [
42]; (3) in apple juice fermented by different LAB strains, the 3-methyl-1-butanol concentration ranged from 4.5–16.9 ng/mL, compared to 73.2 ng/mL in the unfermented juice, among the strains studied,
L. acidophilus fermented juice had a 93% reduction in 3-methyl-1-butanol concentration [
19]; (4) in apple juice fermented by the yeast
S. cerevisiae, the 3-methyl-1-butanol concentration was 644 ng/mL; however, the concentration of 3-methyl-1-butanol reduced to 42.1 ng/mL after
L. plantarum sequential fermentation [
51]; and (5) in cashew apple juice fermented by
L. acidophilus, the 3-methyl-1-butanol concentration reduced by 2 times, compared to other LAB strains (either
L. plantarum or
L. casei) studied [
26].
In 10 studies, the concentration of 2-phenylethyl alcohol, which has a flowery smell, was increased or it was detected after fermentation: (1) In Chinese wolfberry juice fermented by
L. plantarum, the 2-phenylethyl alcohol concentration was 246.4 µg/mL, where it was not detected in the unfermented juice or the fermented juice by other LAB strains [
50]; (2) in non-pH-adjusted (2.65) bog bilberry juice fermented by two strains of
L. plantarum, the 2-phenylethyl alcohol concentration was 1731.2 and 1775.8 ng/mL, compared to 663.5 ng/mL in the unfermented juice. However, if the pH of the juice was adjusted to pH 3.5, the initial 2-phenylethyl alcohol concentration of 617.5 ng/mL was decreased after fermentation by two strains of
L. plantarum to 459.7 and 463.4 ng/mL [
34]; (3) in grape juice fermented by LAB, the 2-phenylethyl alcohol concentration was 40.6 ng/mL, where it was not detected in the unfermented juice [
43]; (4) in horse gram sprouts fermented by two
L. plantarum strains, the 2-phenylethyl alcohol concentration was 1290 and 780 ng/g, compared to 40 ng/g in raw seed [
37]; (5) in goji juice fermented by different combinations of bacterial strains (either
L. plantarum,
L. rhamnosus,
L. reuteri,
B. velezensis, or
B. licheniformis), the 2-phenylethyl alcohol concentration ranged from 362.6 to 494.0 ng/g, compared to 103.1 ng/g in the unfermented juice [
25]; (6) in jujube pulp fermented by a mixture of
L. plantarum,
L. rhamnosus, and
S. thermophilus, the 2-phenylethyl alcohol concentration was 283 ng/g, where it was not detected in the unfermented juice [
47]; (7) in okara fermented with a monoculture of
Rhizopus oligosporus (
R. oligosporus) fungi, the 2-phenylethyl alcohol concentration increased by 20 times, compared to its concentration in the unfermented okara, whereas with a mixed culture of
R. oligosporus fungi and
Yarrowia lipolytica (
Y. lipolytica) yeast, the concentration increased by 8.5 times [
38]; (8) in mango slurry fermented by yeast
S. cerevisiae, the 2-phenylethyl alcohol concentration was 4 to 23 times higher, compared the concentration after LAB fermentation, where it was not detected in the unfermented mango slurry [
41]; (9) in papaya juice fermented by
L. plantarum, the 2-phenylethyl alcohol concentration was doubled, compared to the concentration in
L. acidophilus fermented juice, where it was not detected in the unfermented juice [
23]; and (10) in durian pulp fermented by
L. casei mixed with the yeast
W. saturnus, 2-phenylethyl alcohol was detected, where it was not detected in the unfermented pulp, or in the pulp fermented by a
L. casei monoculture [
30].
Furthermore, compared to unfermented juice, one study reported that almost half of the alcohols detected decreased in
L. casei fermented tomato juice, likewise in
L. plantarum fermented tomato juice, most of the alcohols detected decreased. However, due to the generation of new alcohols, the relative peak area (RPA) for total alcohols increased to 59.9% and 49.7% in juice fermented by either
L. casei or
L. plantarum, respectively, compared to a 49.3% RPA in the unfermented juice [
32]. On the other hand, LAB fermentation increased the overall combined alcohol concentration of fruit and vegetable juices in 4 studies: (1) LAB fermentation of apple juice increased the overall combined alcohol concentration by 10 times compared to its concentration in the unfermented juice, demonstrating that most of the alcohols were produced during fermentation [
21]; (2) LAB fermentation of grape juice increased the total combined alcohol concentration by 102.4% [
43]; (3) LAB fermentation of kiwifruit juice (Xuxiang and Hongyang cultivars) increased the total combined alcohol concentration by 39, 107, and 56% in Xuxiang cultivar juice fermented by either
L. acidophilus,
L. helveticus, or
L. plantarum, respectively, and by 25, 30, and 26% in Hongyang cultivar juice fermented by either
L. acidophilus,
L. helveticus or
L. plantarum, respectively [
49]; and (4) LAB fermentation increased the total combined alcohol concentration of jujube juice (Varieties of Muzao and Hetian) by 66.5% in
L. acidophilus fermented Muzao juice and 33.7% in
L. casei fermented Hetian juice [
20]. In another study, the total combined alcohol concentration of mango slurry fermented with yeast strains was nearly 10 times higher compared to mango slurry fermented with LAB strains [
41].
Overall, alcohols such as 1-hexanol (11 papers), 3-methyl-1-butanol (10 papers), 2-phenylethyl alcohol (10 papers), benzyl alcohol (7 papers), ethanol (7 papers), and 1-octanol (5 papers) have been reported to have increased or were only detected after the fermentation of fruit and vegetable juices, mainly by LAB. However, it is important to note that for some substrates, the concentration of ethanol (6 papers), 3-methyl-1-butanol (5 papers), 1-hexanol (3 papers), and 1-octanol (1 paper) has been reported to have decreased after fermentation.
3.4. Esters
Esters, which have sweet and fruity notes, are formed when carboxylic acids linked with coenzyme-A (CoA) are esterified with alcohols [
60]. Volatile esters were found in fermented fruit and vegetable juices in 27 of the 35 papers reviewed in this report. The sensory detection threshold for esters is lower than that of the corresponding alcohol or acid [
61]. The majority of esters reported were either ethyl esters or acetate esters. Ethanol and acyl-CoA derivatives of fatty acids combine to form ethyl esters. Acetyl-CoA and alcohols, such as ethanol or higher alcohols, produced from amino acid metabolism, combine to form acetate esters [
62]. The key ester compounds reported in the reviewed studies were the acetate esters: ethyl acetate (ethyl ethanoate); 3-methylbutyl acetate (isoamyl acetate/isopentyl acetate); 2-phenylethyl acetate and hexyl acetate, and ethyl esters: ethyl butanoate (ethyl butyrate); ethyl-3-methyl butanoate (ethyl isovalerate/ethyl isopentanoate); ethyl hexanoate (ethyl caproate); ethyl octanoate (ethyl caprylate); ethyl dodecanoate (ethyl laurate); ethyl propanoate (ethyl propionate); ethyl-2-methyl-butanoate; ethyl hexadecanoate (ethyl palmitate) and others: hexyl formate; methyl 3-methylbutanoate (methyl isovalerate/methyl isopentanoate); 3-methylbutyl 3-methylbutanoate (isoamyl isovalerate/isopentyl isopentanoate) and methyl 2-hydroxybenzoate (methyl salicylate).
Eleven of the papers stated that ethyl acetate, which is formed by alcohol acetyltransferases from the reaction between acetyl Co-A and ethanol [
20], was primarily responsible for the fruity flavour of fermented fruits and vegetable juices. A variety of LAB strains [
19,
20,
21,
22,
23,
33,
44,
45,
50] were used in all studies except two; one used LAB combined with a yeast [
30], and the other used fungi combined with a yeast [
38]. In these studies, it was reported that for Chinese wolfberry juice fermented by either
L. plantarum,
L. paracasei,
L. acidophilus, or
L. helveticus, the ethyl acetate concentration was 6931, 4827.1, 4925.4, and 7323.3 µg/mL, respectively, compared to 774.5 µg/mL in the unfermented juice, where it was not detected in the juice fermented by
L. casei or
B. lactis [
50]. In Muzao jujube juice fermented by either
L. plantarum or
L. acidophilus, the ethyl acetate concentration was 111.7 and 64.2 µg/mL, respectively, where it was not detected in the unfermented juice or juice fermented by other LAB [
20]. Further, in durian pulp fermented by
L. casei combined with a yeast
W. saturnus, ethyl acetate was detected, whereas it was not detected in sole
L. casei fermentation or unfermented pulp [
30]. However, Liu et al. [
32] found that in tomato juice, prior to fermentation, ethyl acetate was detected, where it was not detected after fermentation by LAB, and in mango slurry fermented by either yeast or LAB, the ethyl acetate concentration was reduced by 1.2–1.8 times, compared to its concentration in the unfermented slurry [
41].
In 3 experiments, ethyl butanoate, which is formed by a reaction between ethanol and butyryl-CoA during LAB fermentation [
21], was the next most common ester compound: (1) The ethyl butanoate concentration in a distillate prepared using simple distillation of
L. rhamnosus fermented melon by-product was 8760 ng/mL, compared to 700 ng/mL in the unfermented melon by-product distillate, and the ethyl butanoate concentrations of both the fermented and unfermented by-product distillates produced using vacuum distillation were at least 10 times lower than in the simple distillation distillates [
44]; (2) in apple juice fermented by different LAB, the ethyl butanoate concentration ranged from 16.3 to 23.1 ng/g, compared to 2.1 ng/g in the unfermented juice [
21]; and (3) in papaya juice fermented by LAB, ethyl butanoate was detected, where it was not detected in the unfermented juice [
23].
2-Phenylethyl acetate, which is formed by a reaction between 2-phenylethyl alcohol and acetyl CoA, was the third most commonly reported (3 studies) ester in juices after fermentation: (1) In horse gram sprouts fermented by
L. plantarum, the 2-phenylethyl acetate concentration was 220 ng/g, where it was not detected in raw seeds [
37]; (2) in grape juice fermented by a mixed culture of
L. plantarum and
L. brevis, the 2-phenylethyl acetate concentration was 3.5 ng/mL, compared to its concentration in the unfermented juice (1.6 ng/mL) [
43]; and (3) in durian pulp fermented by a combination of
L. casei and
W. saturnus yeast, 2-phenylethyl acetate was detected, whereas it was not detected during
L. casei only fermentation or in the unfermented pulp [
30].
Propyl acetate, which is formed by a reaction between propanol and acetyl-CoA, was detected in cherry juice fermented by either
L. plantarum,
L. rhamnosus, or
L. paracasei, where the propyl acetate concentration was 18.5–83.4, 1186.7, and 201.6 ng/mL, respectively, compared to about 0.01 ng/mL in the unfermented juice. In this study, the formation of propyl acetate during fermentation appeared to correlate with acetic acid production. As there was a low concentration of acetic acid after fermentation by
L. rhamnosus or
L. paracasei, this was taken as evidence of the conversion of acetic acid to the corresponding ester. In contrast, in the same study, fermentation by
L. plantarum resulted in a high concentration of acetic acid and a lower concentration of propyl acetate [
29].
Overall the total combined ester concentration increased in 6 studies after fermentation due to the availability of alcohol precursors [
52]: (1) In Muzao jujube juice fermented by either
L. acidophilus or
L. plantarum, the total combined ester concentration was 65.4 and 156.7 µg/mL, respectively, compared to 4.7 µg/mL in the unfermented juice [
20]; (2) in mixed juices (apple juice, orange juice, carrot juice, and Chinese jujube juice) fermented by LAB mixed culture (
L. plantarum,
Bifidobacterium breve (
B. breve), and
S. thermophilus), the total combined ester concentration was 415 ng/mL, compared to 239 ng/mL in the unfermented juice [
35]; (3) in apple juice fermented by different LAB, the total combined ester concentration ranged from 81.8 to 92.9 ng/g, compared to 33.7 ng/g in the unfermented juice [
21]; (4) in grape juice fermented by LAB, the total combined ester concentration increased by 83.76%, compared to its concentration in the unfermented juice [
43]; (5) in mango slurry fermented by LAB and yeast, the total combined ester concentration increased, compared to the unfermented slurry, with the yeast
S. cerevisiae generating a significantly (
p < 0.05) higher number of esters present at a high concentration than LAB [
41]; and (6) in pomegranate juice fermented by LAB, the total combined ester concentration increased, compared to the concentration in the unfermented juice [
24].
On the other hand, 3 studies reported a reduction in the total combined ester concentration after fermentation, possibly due to hydrolysis into their corresponding acids and alcohols [
47]: (1) In apple juice fermented by either
L. plantarum,
L. helveticus,
L. casei,
L. paracasei,
L. acidophilus or
B. lactis, the total combined ester concentration was 1090, 1279.6, 787.5, 695.9, 702.3, and 643.1 ng/mL, respectively, compared to 1410.7 ng/mL in the unfermented juice [
19]; (2) in jujube juice fermented by a mixture of
L. plantarum,
L. rhamnosus, and
S. thermophilus, the total combined ester concentration was 1541 ng/g, compared to 5814 ng/g in the unfermented juice [
47]; and (3) in tomato juice fermented by either
L. casei or
L. plantarum, the total combined ester concentration was 1.6 times and 7 times lower, respectively, compared to the concentration in the unfermented juice [
32].
Overall, esters such as ethyl acetate (11 papers), ethyl butanoate (3 papers), 2-phenylethyl acetate (3 papers), and propyl acetate (1 paper) have been reported to increase or were only detected after fermentation of fruit and vegetable juices, mainly by LAB. However, the concentration of ethyl acetate did decrease after fermentation in 2 studies.
3.5. Ketones
A number of ketones were identified in 26 studies investigating the fermentation of vegetable and fruit juices. 3-Hydroxy-2-butanone (acetoin), 2,3-butanedione (diacetyl), 2-propanone (acetone), 1-hydroxy-2-propanone (hydroxy acetone), 2-butanone (methyl ethyl ketone), 2-pentanone, 2-hydroxy-3-pentanone, 3-methyl-4-methylene-2-hexanone, 2-heptanone, 4-heptanone, 4-methyl-2-heptanone, 6-methyl-5-hepten-2-one (sulcatone), 2-octanone, 2-nonanone, 2-dodecanone, 2-undecanone, 2-tridecanone, 2-tetradecanone, (E)-6,10-dimethylundeca-5,9-dien-2-one (geranyl acetone), 4-cyclopentene-1,3-dione, and 1-phenylethanone (acetophenone) were identified in the reviewed papers.
3-Hydroxy-2-butanone (acetoin), which imparts a creamy/buttery note, was the most frequently detected ketone produced during the fermentation of vegetables and fruits. Citrate in vegetable and fruit juices can be directly converted to acetoin (
Figure 3) by some LAB strains exhibiting citrate permease and citrate lyase activities. Citrate can be converted by LAB to pyruvate via oxaloacetate, then to acetaldehyde-thiamine pyrophosphate (TPP) through a decarboxylation process, and finally to acetaldehyde-TPP through an enzymatic reaction involving α-acetolactate synthase, resulting in the synthesis of α-acetolactate. α-Acetolactate synthase has a low affinity for pyruvate; therefore, an excess of pyruvate is required to favour this reaction. In the presence of citrate and sugars, homofermentative LAB will convert pyruvate directly to α-acetolactate when less NADH is generated than pyruvate. Heterofermentative LAB will, however, accumulate pyruvate at low pH when citrate is the sole carbon source. Further, due to the instability of α-acetolactate, it is readily decarboxylated enzymatically or chemically to yield acetoin. Acetoin can also be synthesised from diacetyl via the enzyme diacetyl acetoin reductase [
63,
64,
65,
66]. In addition, when the pH of the medium is between 5 and 8,
Lactococcus lactis can also produce acetoin from the catabolism of aspartic amino acid [
67] (
Figure 4). The acetoin concentration increased after LAB fermentation of fruit and vegetable juices in 8 studies: (1) In Chinese wolfberry juice fermented by either
L. plantarum,
L. paracasei,
L. helveticus, or
B. lactis, the acetoin concentration was 346.3, 267.4, 528.1, and 422.1 µg/mL, respectively, compared to 29.1 µg/mL in the unfermented juice, where it was not detected in the juice fermented by
L. casei or
L. acidophilus [
50]; (2) in Muzao jujube juice fermented either
L. plantarum or
L. helveticus, the acetoin concentration was 29.9 and 30.8 µg/mL, respectively, compared to 17.5 µg/mL in the unfermented juice. However, the acetoin concentration was reduced to 10.5 µg/mL in
L. acidophilus fermented juice and acetoin was not detected in
L. casei fermented juice [
20]; (3) in kiwifruit juice (Xuxiang, and Hongyang cultivars), the acetoin concentration was 2621.6 and 1348.9 ng/mL in the Xuxiang cultivar juice fermented by either
L. helveticus or
L. plantarum, respectively, where it was not detected in the
L. acidophilus fermented juice or in the unfermented juice. The acetoin concentration was 8431.7 and 4390.6 ng/mL in Hongyang cultivar juice fermented by either
L. helveticus or
L. plantarum, respectively, where it was not detected in the
L. acidophilus fermented juice or in the unfermented juice [
49]; (4) in elderberry juice fermented by either
L. plantarum,
L. casei, or
L. rhamnosus, the acetoin concentration was 83.1–496.7, 90.7–314.5, and 41.4–456.2 ng/mL, respectively, compared to 1.4–22.1 ng/mL in the unfermented juice [
22]; (5) in cherry juice fermented by either
L. rhamnosus or
L. paracasei, the acetoin concentration was 260.7 and 5.9 ng/mL, respectively, compared to 0.001 ng/mL in the unfermented juice, and in cherry juice fermented by different
L. plantarum strains, the acetoin concentration ranged from 44 to 287.9 ng/mL, compared to 0.002 ng/mL in the unfermented juice [
29]; (6) the acetoin concentration in a distillate prepared using vacuum distillation from orange pomace fermented by
L. rhamnosus was 450 ng/mL, compared to 110 ng/mL in the unfermented pomace distillate, where acetoin was not detected in distillates from either fermented or unfermented pomace using the simple distillation method [
44]; (7) in mung beans fermented by two
L. plantarum strains, the acetoin concentration was 2.8 and 7.5 times higher, compared to the concentration in the unfermented mung beans [
45]; and (8) in papaya juice fermented by either
L. plantarum or
L. acidophilus, the acetoin concentration was 2.2 and 3.7 times higher, respectively, compared to the unfermented juice [
23]. In 5 studies, acetoin was only detected after fermentation of juices: (1) In okara fermented by a LAB co-culture (
L. acidophilus,
L. rhamnosus, and
P. acidilactici), the acetoin concentration was 166.3 µg/g, where it was not detected in LAB monocultures [
46]; (2) in horse gram sprouts fermented by
L. plantarum, the acetoin concentration was 440 ng/g [
37]; (3) in goji juice fermented by a bacterial mixture of
L. rhamnosus,
L. reuteri, and
B. velezensis, the acetoin concentration was 87.2 ng/g juice [
25]; (4) in mango slurry fermented by
S. thermophilus, acetoin was detected, where it was not detected after fermentation by yeast
S. cerevisiae or other LAB [
41]; and (5) in durian pulp fermented by
L. casei in sequential co-culture with
W. saturnus yeast, acetoin was detected, where it was not detected in
L. casei monoculture [
30]. Note, that in apple juice fermented by either
L. acidophilus,
L. helveticus, or
L. paracasei, the acetoin concentration was reported to have decreased to 0.8 ng/mL, 1.2 ng/mL, and 4.3 ng/mL, respectively, compared to 5.4 ng/mL in the unfermented juice, where it was not detected in
L. plantarum,
L. casei, or
B. lactis fermented apple juice [
19].
The second most commonly reported ketone was 2,3-butanedione (diacetyl), which imparts creamy/buttery notes. It is produced by LAB from citrate present in juice (
Figure 3). As discussed for the acetoin production pathway, α-acetolactate can be directly converted into diacetyl through nonenzymatic oxidative carboxylation in the presence of molecular oxygen [
63,
65]. Diacetyl was reported to be increased by the presence of some LAB during fermentation of juices in 6 studies: (1) In Chinese wolfberry juice fermented by either
L. plantarum,
L. casei,
L. paracasei, or
L. acidophilus, the diacetyl concentration was 45.1, 51, 71.9, and 28.1 µg/mL, respectively, where it was not detected in the unfermented juice nor the juice fermented by
L. helveticus or
B. lactis [
50]; (2) in elderberry juice fermented by either
L. rhamnosus,
L. plantarum, or
L. casei strains, the diacetyl concentration ranged from 37–586.8, 16.2–400.7, and 221.9–276.6 ng/mL, respectively, compared to 3.3–12.2 ng/mL in the unfermented juice [
22]; (3) in kiwifruit juice (Xuxiang, and Hongyang cultivars), the diacetyl concentration was 261.1 ng/mL in Xuxiang cultivar juice fermented by
L. helveticus, where it was not detected in Xuxiang cultivar juice fermented by other LAB or in the unfermented juice. Interestingly, with the Hongyang cultivar juice, diacetyl was not detected in the unfermented juice or in any of the LAB fermented juices [
49]; (4) in watermelon juice fermented by either
L. plantarum,
L. brevis,
L. casei, or
L. rhamnosus, the diacetyl concentration was 1.46, 1.47, 62.5, and 85.7 ng/mL, respectively, where it was not detected in the
P. pentosaceus fermented juice and the unfermented juice [
48]; (5) in mango slurry fermented by
L. casei, diacetyl was detected, whereas it was not detected in other LAB or yeast fermentations [
41]; and (6) in pomegranate juice fermented by
L. plantarum strains, the diacetyl concentration increased, compared to the concentration in the unfermented juice [
24]. However, in tomato and pepper pomace fermented by either
Trichoderma atroviride (
T. atroviride) or
Aspergillus sojae (
A. sojae), the diacetyl concentration was reduced compared to the concentration present in the unfermented pomace [
40].
Overall, after LAB fermentation, the total combined ketone concentration increased in 5 studies: (1) In okara fermented by LAB co-culture with
L. acidophilus,
L. rhamnosus, and
P. acidilactici, the total combined ketone concentration was 2355.6 µg/g, compared to 116.1 µg/g in the unfermented okara; however, in okara fermented by monocultures of either
L. acidophilus,
L. rhamnosus, or
P. acidilactici, the total combined ketone concentration was 98.8, 64.3, and 57.8 µg/g, respectively. During a monoculture fermentation, unstable aldehydes and ketones may be reduced to primary and secondary alcohols, whereas in a co-culture fermentation, synergic interactions between strains may instead result in the production of higher levels of ketones, which could be linked to the oxidation of alcohols [
46]; (2) in cherry juice fermented by either
L. rhamnosus or
L. paracasei, the total combined ketone concentration was 285.1 and 11.3 ng/mL, respectively, compared to 7.2 ng/mL in the unfermented juice. In the cherry juice fermented by different
L. plantarum strains, the total combined ketone concentration ranged from 48.3 to 292.4 ng/mL, compared to 6.9 ng/mL in the unfermented juice [
29]; (3) in apple juice fermented by either
L. plantarum,
L. helveticus,
L. casei,
L. paracasei,
L. acidophilus, or
B. lactis, the total combined ketone concentration was 17.0, 27.6, 29.6, 56.6, 22.5, and 26.4 ng/mL, respectively, compared to 16.6 ng/mL in the unfermented juice [
19]; (4) in another study, using apple juice fermented by different LAB strains, the total combined ketone concentration ranged from 10.1 to 11.7 ng/g, compared to 2.6 ng/g in the unfermented juice [
21]; and (5) LAB fermentation of kiwifruit juice (Xuxiang, and Hongyang cultivars) increased the total combined ketone concentration by 2.6, 5.2, and 2.6 times in Xuxiang cultivar juice fermented by either
L. acidophilus,
L. helveticus, or
L. plantarum, respectively, and by 6.3, 75, and 37 times in Hongyang cultivar juice fermented by either
L. acidophilus,
L. helveticus, or
L. plantarum, respectively [
49]. Moreover, in mango slurry fermented by LAB, the total combined ketone concentration was 1.2–1.8 times higher compared to the concentration in the unfermented juice, whereas in mango slurry fermented by the yeast
S. cerevisiae, the total combined ketone concentration was 1.4–2.7 times lower compared to the concentration in the unfermented juice [
41].
Overall, ketones, which are key contributors to dairy notes, such as acetoin (13 papers) and diacetyl (6 papers) have been reported to increase or were only detected after the fermentation of fruit and vegetable juices. However, in two studies, the concentration of acetoin (1 paper), and diacetyl (1 paper) decreased after fermentation by LAB and fungi, respectively.
In 8 studies, L. plantarum was the main LAB producing acetoin in fermented juices, followed by L. helveticus (3 studies) and L. rhamnosus (3 studies). The two main LAB that produced high diacetyl concentrations in fermented juices were L. plantarum (4 papers) and L. casei (4 papers). Overall, for the papers reviewed, L. plantarum produced more of the creamy flavours of acetoin and diacetyl compared to other LAB strains studied.
3.6. Aldehydes
Aldehydes were present at lower concentrations after the fermentation of fruit and vegetables in 26 out of 35 studies. During the fermentation process, aldehydes are generated via alcohol oxidation or acid decarboxylation. The main aldehyde compounds detected after fermentation included ethanal (acetaldehyde), phenyl methanal (benzaldehyde), 2-methyl butanal, 3-methyl butanal (isovaleraldehyde), pentanal (valeraldehyde), hexanal (caproaldehyde), (E)-2-hexenal, heptanal (enanthaldehyde), octanal (caprylaldehyde), (E)-2-octenal, nonanal (pelargonaldehyde), (E)-2-nonenal, decanal (capraldehyde), dodecanal (lauraldehyde), 2,4-dimethyl-benzaldehyde, octadecanal (stearaldehyde), 2-undecenal, tridecanal, 3,5-dimethyl-benzaldehyde, and benzeneacetaldehyde/phenylacetaldehyde.
Acetaldehyde provides fermented juices their distinct flavour, and it is produced by LAB from the amino acid threonine [
68] or from sugars via the PK (phosphoketolase) pathway, and by yeast from sugars via the EMP (Embden–Meyerhof–Parnas) pathway [
13]. At lower concentrations, acetaldehyde improves the flavour of fermented juice; however, at higher concentrations (200 µg/g or 200 µg/mL or above) [
20,
21], it may negatively influence the flavour of fermented juices. Acetaldehyde was detected in 6 studies after LAB fermentation: (1) In Muzao jujube juice fermented by
L. acidophilus, the acetaldehyde concentration was 19.9 µg/mL, compared to 1.5 µg/mL in the unfermented juice, with other LAB strains generating slightly higher or lower acetaldehyde concentrations compared to the unfermented juice [
20]; (2) in kiwifruit juice (Xuxiang and Hongyang cultivars), Xuxiang cultivar juice fermented by either
L. plantarum,
L. acidophilus, or
L. helveticus the acetaldehyde concentration was 1013.1, 136.1, and 124.3 ng/mL, respectively, compared to 109.5 ng/mL in the unfermented juice, whereas in the Hongyang cultivar juice fermented by either
L. plantarum or
L. acidophilus, the acetaldehyde concentration was 1075.6 and 159.7 ng/mL, respectively, compared to 95.7 and 25.5 ng/mL in the unfermented juice and fermented juice by
L. helveticus, respectively [
49]; (3) in apple juice fermented by either
L. casei, L. rhamnosus,
L. plantarum, or
L. acidophilus, the acetaldehyde concentration was 40.4, 15.0, 27.5, and 21.9 ng/g, respectively, whereas it was not detected in the unfermented juice [
21]; (4) in apple juice fermented by either
L. plantarum,
L. helveticus,
L. casei,
L. paracasei,
L. acidophilus, or
B. lactis, the acetaldehyde concentration was 5.4, 2.1, 4.5, 2.4, 1.9, and 3.0 ng/mL, respectively, whereas it was not detected in the unfermented juice [
19]; (5) in non-pH-adjusted (2.7) sea buckthorn juice, the acetaldehyde concentration was 10.6 ng/mL, which increased after fermentation for 36 and 72 h by
L. plantarum to 13.9 and 15.8 ng/mL, respectively. However, if the pH of the juice was adjusted to pH 3.5, the initial acetaldehyde concentration of 10.8 ng/mL decreased after
L. plantarum fermentation for 36 and 72 h to 1.1 and 1.2 ng/mL, respectively [
42]; and (6) in watermelon juice fermented by either
L. plantarum or
P. pentosaceus, the acetaldehyde concentration was 4.6 and 3.2 ng/mL, respectively, compared to 2.3, 2.0, 0.5, and 0.5 ng/mL in the unfermented juice, and
L. brevis,
L. casei, or
L. rhamnosus fermented juices, respectively [
48]. The concentration of acetaldehyde in 6 studies reported here was still well below the concentration that has been reported to adversely affect flavour, indicating that acetaldehyde may have a positive impact on the overall flavour profile of fermented juices if it is above the minimum concentration required for perception. Note, in Chinese wolfberry juice fermented by either
L. plantarum,
L. casei,
L. paracasei,
L. helveticus, or
B. lactis, the acetaldehyde concentration was reduced to 52.8, 124.2, 123.3, 23.4, and 13.3 µg/mL, respectively, compared to its concentration in the unfermented juice (155.9 µg/mL), where in the juice fermented by
L. acidophilus, the acetaldehyde concentration was 188.2 µg/mL [
50]. Further, the initial acetaldehyde concentration in a distillate prepared using simple distillation from unfermented melon by-product was 1320 ng/mL, which was reduced to 470 ng/mL in the
L. rhamnosus fermented melon by-product distillate. Moreover, when using the vacuum distillation method, in the unfermented melon by-product distillate, the acetaldehyde concentration was 160 ng/mL, where it was only 20 ng/mL in the distillate from
L. rhamnosus fermented melon by-product. Some LAB can convert acetaldehyde to ethanol and acetic acid, which could explain the decrease in acetaldehyde concentration in some fermentations [
44].
Another important aldehyde from a flavour perspective as it imparts a pleasant aroma to fermented juices is benzaldehyde, which is generated by LAB from the amino acid phenylalanine. The conversion of phenylalanine to benzaldehyde by LAB is initiated by the aminotransferase enzyme. The resulting phenyl pyruvic acid is chemically converted to benzaldehyde in the presence of oxygen and manganese [
58,
69]. The benzaldehyde concentration increased after fermentation of vegetable and fruit juices in 7 studies: (1) In Chinese wolfberry juice fermented by either
L. plantarum,
L. paracasei, or
L. acidophilus, the benzaldehyde concentration was 117.2, 68.1, and 40.7 µg/mL, respectively, where it was not detected in the unfermented juice nor in juice fermented by other LAB strains [
50]; (2) in kiwifruit juice (Xuxiang and Hongyang cultivars), Xuxiang cultivar juice fermented by
L. acidophilus, the benzaldehyde concentration was 490.3 ng/mL, compared to 369.7 ng/mL in the unfermented juice. In contrast, it was not detected in the unfermented Hongyang cultivar juice nor in the Xuxiang cultivar juice fermented by
L. helveticus or
L. plantarum and all LAB fermented Hongyang cultivar juices [
49]; (3) in bog bilberry juice fermented by two
L. plantarum strains, the benzaldehyde concentration was 55.5 and 62.3 ng/mL, compared to 41.8 ng/mL in the unfermented juice [
34]; (4) in non-pH-adjusted (2.7) sea buckthorn juice, the benzaldehyde concentration was 2.7 ng/mL, which increased after fermentation for 36 and 72 h by
L. plantarum to 5.4 and 7.9 ng/mL, respectively. However, if the pH of the juice was adjusted to pH 3.5, the initial benzaldehyde concentration of 2.3 ng/mL decreased after
L. plantarum fermentation for 36 and 72 h to 1.1 and 1.7 ng/mL, respectively [
42]; (5) in goji juice fermented by different combinations of bacterial strains (either
L. plantarum,
L. rhamnosus,
L. reuteri,
B. velezensis, or
B. licheniformis), the benzaldehyde concentration ranged from 55.5 to 101.4 ng/g, compared to 46.3 ng/g in the unfermented juice [
25]; (6) in durian pulp fermented by
L. casei monoculture, the benzaldehyde concentration was 2.9 times higher, compared to its concentration in the sequential co-culture with yeast
W. saturnus, and it was not detected in the unfermented pulp. This difference is because LAB can convert phenylalanine amino acid to benzaldehyde; however, yeast preferentially convert phenylalanine amino acid to phenylethyl alcohol via the Ehrlich pathway, resulting in a higher quantity of benzaldehyde in LAB fermentations [
30]; and (7) in papaya juice fermented by
L. plantarum, the benzaldehyde concentration was 2 times higher, compared to the concentration after the
L. acidophilus fermentation or in the unfermented juice [
23]. Though the benzaldehyde concentration increased after LAB fermentation, it also reduced in 4 studies: (1) in Hetain jujube juice fermented by LAB, the benzaldehyde concentration ranged from 22.1 to 29.7 µg/mL, compared to 33 µg/mL in the unfermented juice [
20]; (2) in jujube pulp fermented by a mixture of
L. plantarum,
L. rhamnosus, and
S. thermophilus, the benzaldehyde concentration was 3516 ng/g, compared to 4672 ng/g in the unfermented pulp [
47]; (3) in cherry juice fermented by various
L. plantarum strains, the benzaldehyde concentration ranged from 15.3 to 33.4 ng/mL, compared to 100.5 ng/mL in the unfermented juice, and in cherry juice fermented by
L. rhamnosus, the benzaldehyde concentration was 76 ng/mL, compared to 90.5 ng/mL in the unfermented juice [
29]; and (4) in okara fermented by LAB monocultures of
L. rhamnosus or
P. acidilactici and a co-culture (
L. acidophilus,
L. rhamnosus, and
P. acidilactici), the benzaldehyde concentration was 22.4, 45, and 46.2 µg/g, respectively, compared to 114.6 µg/g in the unfermented okara, and in okara fermented by
L. acidophilus, the benzaldehyde concentration was only slightly reduced to 109.9 µg/g [
46].
Hexanal and nonanal are two other important aldehydes that may influence the flavour profile of fermented juices. Hexanal, which is produced from linoleic fatty acid, imparts a fresh, green, grassy, waxy, fatty, and unpleasant aroma to fermented fruit and vegetable juices [
54]. After fermentation, the hexanal concentration was reduced in goji juice [
25], mung bean [
45], watermelon juice [
48], sea buckthorn juice [
42], kiwifruit juice [
49], and okara pulp [
38,
46]. In addition, it was not detected after the fermentation of Chinese wolfberry juice [
50]. Jin et al. [
41] reported that in mango slurry fermented by LAB, the nonanal was detected, where it was not detected after yeast fermentation. Further, the nonanal concentration was reduced in fermented yam juice [
33], okara pulp [
46], mung bean [
45], watermelon juice [
48], kiwifruit juice [
49], and sea buckthorn juice [
42]. It was also not detected during the fermentation of goji juice and durian pulp [
25,
30].
Overall, the total combined aldehyde concentration was reduced after the fermentation of vegetable and fruit juices in 8 studies. This was probably due to the conversion of aldehydes either by reduction to alcohols or oxidation to acids [
23]: (1) In Chinese wolfberry juice fermented by either
L. plantarum,
L. casei,
L. paracasei,
L. acidophilus, L. helveticus, or
B. lactis, the total combined aldehyde concentration was 332.8, 680.1, 500.5, 983.3, 422.1, and 492.6 µg/mL, respectively, compared to 1841.2 µg/mL in the unfermented juice [
50]; (2) in unfermented jujube (Muzao and Hetain varieties) juice, the total combined aldehyde concentration was 102.8 and 121.3 µg/mL, respectively, compared to 65.4–94.7 and 61.9–83.9 µg/mL in the fermented Muzao, and Hetain varietal juices by different LAB strains, respectively. However, the total combined aldehyde concentration increased by 38.3% in Hetain varietal juice fermented by
L. plantarum and 158% in Muzao varietal juice fermented by
L. acidophilus [
20]; (3) in kiwifruit juice (Xuxiang and Hongyang cultivars), Xuxiang cultivar juice fermented by either
L. acidophilus,
L. helveticus, or
L. plantarum, the total combined aldehyde concentration was 2411.9, 4248.7, and 4915.8 ng/mL, respectively, compared to 15,316.3 ng/mL in the unfermented juice, where in the Hongyang cultivar juice fermented by either
L. acidophilus,
L. helveticus, or
L. plantarum, the total combined aldehyde concentration was 2466.1, 2528.9, and 3866.4 ng/mL, respectively, compared to 23,477.9 ng/mL in the unfermented juice [
49]; (4) in cherry juice fermented by various
L. plantarum strains, the total combined aldehyde concentrations ranged from 43.5 to 78.2 ng/mL, compared to 208.5 ng/mL in the unfermented juice [
29]; (5) in okara fermented by
L. acidophilus,
P. acidilactici, or
L. rhamnosus, and a co-culture (
L. acidophilus,
P. acidilactici, and
L. rhamnosus), the total combined aldehyde concentration was 866.1, 282.9, 25.3, and 68.3 µg/g, respectively, compared to 1450.6 µg/g in the unfermented okara [
46]; (6) in watermelon juice fermented by either
L. plantarum,
L. rhamnosus,
L. casei,
L. brevis, or
P. pentosaceus, the total combined aldehyde concentration reduced by > 50% compared to the unfermented juice [
48]; (7) in grape juice fermented by LAB, the total combined aldehyde concentration reduced by 45.4% [
43]; and (8) in tomato juice fermented by either
L. plantarum or
L. casei, the total combined aldehyde concentration was 15 and 164 times lower, respectively, compared to the concentration in the unfermented juice [
32]. Note, that in apple juice fermented by either
L. plantarum,
L. helveticus,
L. casei,
L. paracasei,
L. acidophilus, or
B. lactis, the total combined aldehyde concentration was reported to have increased to 22.4, 13.3, 7.6, 4.4, 5.4, and 6.0 ng/mL, respectively, compared to 2.5 ng/mL in the unfermented juice [
19].
Higher overall combined aldehyde concentrations, particularly if these are dominated by lipid-derived aldehydes, are likely to have a negative impact on the flavour of fermented juices. Reducing the concentrations of lipid-derived aldehydes is likely to increase the fruity aroma of fermented fruits and vegetables while lowering their green odour character. In all of the studies reviewed, most lipid-derived aldehyde compounds were reduced after LAB fermentation (hexanal (8 papers) and nonanal (8 papers)), whereas acetaldehyde (6 papers) and benzaldehyde (7 papers) were increased or were only detected after fermentation and were responsible for the pleasant aroma of fermented fruit and vegetable juices.
3.7. Acids
Twenty-five studies out of the thirty-five reported that acids contributed to the flavour of fermented vegetables and fruits. An extensive range of short- to long-chain fatty acids, including acetic acid, propanoic acid (propionic acid), 2-methyl-propanoic acid (isobutyric acid), butanoic acid (butyric acid), 3-methyl-1-butanoic acid (isovaleric acid), 2-methyl-1-butanoic acid, caproic acid (hexanoic acid), enanthic acid (heptanoic acid), caprylic acid (octanoic acid), pelargonic acid (nonanoic acid), capric acid (decanoic acid), lauric acid (dodecanoic acid), palmitic acid (hexadecanoic acid), and oleic acid, were detected in the fermented fruits and vegetables, which were formed from sugars or amino acid catabolism [
52].
Acetic acid, which is a key flavour compound of fermented juices, is produced mainly by heterofermentative LAB (
L. brevis,
Limosilactobacillus fermentum (
L. fermentum),
L. reuteri,
L. plantarum,
L. rhamnosus, and
L. casei), which first utilise sugars via the PK pathway and produce acetyl phosphate, which is subsequently converted into acetic acid by the acetokinase enzyme [
13]. However, homofermentative LAB (
L. acidophilus,
S. thermophilus, and
L. helveticus), with high glycolytic flux rates, ferment sugars into solely lactic acid, while under slow growth conditions and low glycolytic flux rates, the homofermentative LAB change to mixed acid fermentation (formic acid, lactic acid, ethanol, and acetic acid) [
70]. Acetic acid can also be produced from citrate, which is found in fruit and vegetable juices [
65]. The acetic acid concentration increased after fermentation of fruit and vegetable juices in 15 studies: (1) In Chinese wolfberry juice fermented by either
L. casei,
L. paracasei, L. helveticus, or
B. lactis, the acetic acid concentration was 19,773.7, 16,093.9, 12,698.4, and 14,011.1 µg/mL, respectively, and it was not detected in the unfermented juice or juice fermented by
L. plantarum or
L. acidophilus [
50]; (2) In two varieties of jujube (Muzao and Hetain) juices fermented by different LAB strains, the acetic acid concentration ranged from 161 to 234.3 µg/mL, and 211.2 to 278.1 µg/mL, respectively, compared to 90.8 µg/mL in Muzao and 104.2 µg/mL in Hetain unfermented varietal juices. Interestingly,
L. helveticus increased the acetic acid concentration by 158.2% in Muzao varietal juice and
L. casei increased it by 166.9% in Hetian varietal juice [
20]; (3) in elderberry juice fermented by either
L. plantarum,
L. casei, or
L. rhamnosus strains, the acetic acid concentration ranged from 205.9–1012.3, 62.3–122.1, and 47.2–132.1 ng/mL, respectively, compared to 0.3–12.2 ng/mL in the unfermented juice [
22]; (4) in cherry juice fermented by various
L. plantarum strains, the acetic acid concentration ranged from 54.8 to 184.8 ng/mL, compared to 0.01 ng/mL in the unfermented juice [
29]; (5) in non-pH-adjusted (2.7) sea buckthorn juice, the acetic acid concentration was 1.0 ng/mL which increased after fermentation for 36 and 72 h by
L. plantarum to 2.1 and 3.5 ng/mL, respectively, where the pH of the juice was adjusted to pH 3.5, the initial acetic acid concentration of 0.8 ng/mL increased after fermentation with
L. plantarum for 36 and 72 h to 50.5 and 85.9 ng/mL, respectively [
42], 6. In grape juice fermented by LAB, the acetic acid concentration was 25.5 ng/mL, and it was not detected in the unfermented juice [
43]; (7) in jujube juice fermented by a mixture of
L. plantarum,
L. rhamnosus, and
S. thermophilus, the acetic acid concentration was 12.2 µg/g, compared to 2.8 µg/g in the unfermented juice [
47]; (8) in horse gram sprouts fermented by
L. plantarum strains, the acetic acid concentration ranged from 4.8 to 5 µg/g, and it was not detected in the raw seed [
37]; (9) in goji juice fermented by different combinations of bacterial strains (either
L. plantarum,
L. rhamnosus,
L. reuteri,
B. velezensis, or
B. licheniformis), the acetic acid concentration ranged from 25.4 to 88.9 ng/g, where it was not detected in the unfermented juice [
25]; (10) in mung bean fermented by
L. plantarum strains, the acetic acid concentration ranged from 0.17 to 0.29 ng/g, where it was not detected in the unfermented mung bean [
45]; (11) in papaya juice fermented by either
L. acidophilus or
L. plantarum, the acetic acid concentration was 5.7 and 2.4 times higher, respectively, compared to the concentration in the unfermented juice [
23]; (12) in mango slurry fermented by either
L. plantarum or
S. thermophilus, the acetic acid concentration was 2.4 and 2.7 times higher, respectively, compared to the concentration in the unfermented mango slurry, and in addition, it was not detected in
L. casei or yeast fermentations [
41]; (13) in tomato juice fermented by
L. plantarum, the acetic acid concentration was 8.6 times higher compared to the concentration in the unfermented juice, where it was not detected in
L. casei fermented juice [
32]; (14) in yam juice fermented by LAB, the acetic acid concentration was 1.5 times higher compared to the concentration in the unfermented yam juice [
33], and 15. in durian pulp fermented by
L. casei, the acetic acid concentration was 1.4 times higher compared to the concentration in a co-culture of
L. casei and yeast
W. saturnus, where it was not detected in the unfermented pulp [
30].
3-Methyl-1-butanoic acid is an important acid flavour compound in dairy foods, which is produced from the amino acid leucine by an aminotransferase enzyme [
57]. However, when present at high concentrations, it may negatively impact on the flavour of fermented juices [
26]. The 3-methyl-1-butanoic acid concentration increased in 3 studies after LAB fermentation: (1) In non-pH-adjusted (2.7) sea buckthorn juice, the 3-methyl-1-butanoic acid concentration was 19.2 ng/mL, which increased after fermentation for 36 and 72 h by
L. plantarum to 25 and 39 ng/mL, respectively, where the pH of the juice was adjusted to pH 3.5, the initial 3-methyl-1-butanoic acid concentration of 13.1 ng/mL increased after
L. plantarum fermentation for 36 and 72 h to 129.3 and 185 ng/mL, respectively [
42]; (2) in tomato juice fermented by
L. plantarum, the 3-methyl-1-butanoic acid concentration was 3 times higher compared to the concentration in the unfermented juice, where, it was not detected in
L. casei fermented juice [
32]; and (3) in papaya juice fermented by
L. plantarum, the 3-methyl-1-butanoic acid concentration was 4.1 times higher compared to the concentration in
L. acidophilus fermented juice, where it was not detected in the unfermented juice [
23].
Butanoic acid, which is produced from the fermentation of sugars through the fatty acid biosynthesis pathway [
71] and can confer a dairy/cheesy aroma to fermented juices, was detected in 3 studies after fermentation: (1) In jujube juice fermented by a mixture of
L. plantarum,
L. rhamnosus, and
S. thermophilus, the butanoic acid concentration was 1487 ng/g, compared to 345 ng/g in the unfermented juice [
47]; (2) in apple juice fermented by different LAB, the butanoic acid concentration ranged from 2.3 to 4.7 ng/g, where it was not detected in the unfermented juice [
21]; and (3) in okara fermented by a combination of
R. oligosporus and
Y. lipolytica, the butanoic acid concentration was 34 times higher compared to the concentration in the juice fermented by
R. oligosporus monoculture [
38]. However, in mango slurry fermented by the yeast
S. cerevisiae, the butanoic acid concentration was 4.4–5.6 times lower compared to the concentration in the unfermented mango slurry, where the butanoic acid concentration was slightly higher after LAB fermentation compared to the unfermented mango slurry [
41].
Hexanoic acid, which can be produced from the fermentation of sugars through the fatty acid biosynthesis pathway or from the cleavage of linoleic acid via hexanal, may impart a fatty/cheesy/sour flavour to fermented juices [
22,
54,
72,
73]. Hexanoic acid was detected after fermentation in 4 studies: (1) In jujube juice fermented by a mixture of
L. plantarum,
L. rhamnosus, and
S. thermophilus, the hexanoic acid concentration was 15.1 µg/g, compared to 7.1 µg/g in the unfermented juice [
47]; (2) in non-pH-adjusted (2.65) bog bilberry juice fermented by two strains of
L. plantarum, the hexanoic acid concentration was 266 and 272.1 ng/mL, compared to 147 ng/mL in the unfermented juice. However, if the pH of the juice was adjusted to pH 3.5, the initial hexanoic acid concentration of 132.6 ng/mL was slightly changed after fermented by two strains of
L. plantarum to 136.9 and 123.2 ng/mL [
34]; (3) in grape juice fermented by LAB, the hexanoic acid concentration was 14.2 ng/mL, where it was not detected in the unfermented juice [
43]; and (4) in mango slurry fermented by different LAB, the hexanoic acid concentration was 1.4–1.7 times higher than the concentration in the unfermented slurry. However, in yeast fermentation, the hexanoic acid concentration was 1.2–1.6 times lower compared to the concentration in the unfermented juice [
41]. According to Li et al. [
20], in Muzao varietal jujube juice, the initial hexanoic acid concentration of 69 µg/mL was reduced after fermentation by different LAB to between 9.2 and 54.5 µg/mL, with the highest reduction being reported for
L. acidophilus fermented juice. In the same study, in Hetain varietal jujube juice, the initial hexanoic acid concentration was 19.0 µg/mL, which increased after fermentation by different LAB to 31.2–75.9 µg/mL, with the highest concentration being obtained in
L. acidophilus fermented juice.
Octanoic acid, which is synthesised from sugars (glucose) through the fatty acid biosynthesis pathway [
74], was detected in 6 studies after the fermentation of juices: (1) In non-pH-adjusted (2.65) bog bilberry juice fermented by two strains of
L. plantarum, the octanoic acid concentration was 3921.6 and 4013.2 ng/mL, compared to 1538.2 ng/mL in the unfermented juice. However, if the pH of the juice was adjusted to pH 3.5, the initial octanoic acid concentration of 1156.5 ng/mL was reduced after fermentation by two strains of
L. plantarum to 655.3 and 722.9 ng/mL [
34]. (2) in goji juice fermented by a bacterial mixture, the octanoic acid concentration ranged from 251.8 to 321.5 ng/g, and it was not detected in the unfermented juice [
25]; (3) in grape juice fermented by LAB, the octanoic acid concentration was 5.7 ng/mL, and it was not detected in the unfermented juice [
43]; (4) in yam juice fermented by LAB, the octanoic acid concentration was 2 times higher compared to the concentration in the unfermented juice [
33]; (5) in durian pulp fermented by
L. casei, the octanoic acid concentration was 2.3 times higher compared to the concentration in the unfermented pulp, and when the pulp was fermented by
L. casei combined with yeast
W. saturnus, the octanoic acid concentration was 2 times lower compared to the concentration in the unfermented pulp [
30]; and (6) in mango slurry, the octanoic acid was detected after fermentation by the yeast
S. cerevisiae, but it was not detected after LAB fermentation [
41].
Chen et al. [
33] reported that
L. plantarum alone or in combination with
S. thermophilus increased acetic, nonanoic, and decanoic acids in fermented yam juice, reducing the astringent odour of the fermented juice.
Overall, volatile acids after LAB fermentation made contributions to the flavour of fruit and vegetable juices, where acetic acid (15 papers) was the most commonly detected volatile acid present in fermented juices, followed by octanoic acid (6 papers), hexanoic acid (4 papers), 3-methyl-1-butanoic acid (3 papers), and butanoic acid (3 papers).
3.8. Terpenes and Norisoprenoids
Terpenes, which are comprised of isoprene (C5) units, impart floral, rose, and fruity flavours. In 13 studies reported here, terpenes were classified as being terpenes or norisoprenoids; interestingly, in 14 studies, some terpene compounds (β-linalool, α-terpineol, β-citronellol, geraniol, β-damascenone, D-limonene, and trans-β-ionone) were classified as either alcohol/hydrocarbon/ketone, or as alkenes in 2 studies. Terpenes are classified based on the number of isoprene units they contain: hemiterpenoids (C5), monoterpenoids (C10), sesquiterpenoids (C15), diterpenoids (C20), sesterterpenoids (C25), triterpenoids (C30), tetraterpenoids (C40) (β-carotene), and polyterpenoids (C > 40) [
12,
75]. β-carotene can be further oxidized into β-ionone, β-damascenone, and β-ionol. Bacteria biosynthesize terpenoids via the 2-C-methyl-D-erythritol-4-phosphate (MEP) pathway [
76], while yeast produce them via the mevalonate pathway [
75]. (β)-Myrcene, D-limonene (1-Methyl-4-(prop-1-en-2-yl) cyclohex-1-ene), ocimene, (β)-linalool (3,7-Dimethyl-1,6-octadien-3-ol), camphene, p-cymene, α-terpinolene, (α)-terpinene, 1,8-cineole, (ɤ)-terpinene, (α)-terpineol (2-(4-Methylcyclohex-3-en-1-yl)propan-2-ol), Citronellyl formate, (β)-damascenone ((E)-1-(2,6,6-Trimethyl-1-cyclohexa-1,3-dienyl) but-2-en-1-one), (β)-citronellol (3,7-Dimethyloct-6-en-1-ol), geraniol ((2E)-3,7-Dimethylocta-2,6-dien-1-ol), trans-(β)-ionone (4-(2,6,6-Trimethylcyclohex-1-en-1-yl) but-3-en-2-one), m-cymene, prenol, (α)-ionene, D-germacrene, valencene, cedrol, phytol, myrtenol, eugenol (2-Methoxy-4-(prop-2-en-1-yl) phenol), and β- phellandrene were the major terpene compounds detected in the reviewed papers.
Linalool, an important flavour terpene, was detected after fermentation of juices in 7 studies: (1) The linalool concentration of a distillate prepared using simple distillation from unfermented orange pomace was 1.6 µg/mL and after fermentation by
L. rhamnosus of the orange pomace, the concentration in the resulting distillate was 2.3 µg/mL. Similarly, in distillates prepared using vacuum distillation, the linalool concentration in the unfermented orange pomace distillate was lower than that in the distillate from
L. rhamnosus fermented orange pomace, at 0.1 µg/mL and 1.1 µg/mL, respectively. In the same study, in the unfermented melon by-product distillate prepared using simple distillation, the linalool concentration was 0.5 µg/mL and in the
L. rhamnosus fermented melon by-product distillate, the concentration was 4.4 µg/mL. However, linalool was not detected in the distillates of fermented and unfermented melon by-products prepared using vacuum distillation [
44]; (2) in elderberry juice fermented by either
L. plantarum or
L. rhamnosus, the linalool concentration was 328.5 and 339.7 ng/mL, respectively, compared to 79.3 and 189.3 ng/mL in the unfermented juices, respectively [
22]; (3) in cherry juice fermented by either
L. rhamnosus,
L. paracasei, or
L. plantarum, the linalool concentration was 39.6, 21.5, and 19.6–29 ng/mL, respectively, compared to 13.7–15.7 ng/mL in the unfermented juice [
29]; (4) in goji juice fermented by a bacterial mixture (
L. plantarum,
L. rhamnosus,
B. velezensis, and
B. licheniformis), the linalool concentration was 22.3 ng/g, where it was not detected in the unfermented juice [
25]; (5) in apple juice fermented by different LAB, the linalool concentration ranged from 3.0 to 4.5 ng/g, compared to 2.0 ng/g in the unfermented juice [
21]; (6) in yam juice fermented by
L. plantarum, the linalool concentration was 21 times higher compared to the concentration in the juice fermented by a combination of
L. plantarum and
S. thermophilus, and it was not detected in the unfermented yam juice [
33]; and (7) in mango slurry fermented by LAB, linalool was detected, where it was not detected in yeast fermented or unfermented slurry [
41]. Though the linalool concentration increased in these studies after fermentation, it was also reduced after fermentation in 2 studies: (1) In papaya juice fermented by either
L. acidophilus or
L. plantarum, the linalool concentration was 1.6 and 1.2 times lower, respectively, compared to the concentration in the unfermented juice [
23]; and (2) in
Momordica charantia juice fermented by
L. plantarum, the linalool concentration was reduced compared to the concentration in the unfermented juice [
18].
D-limonene, a significant flavour terpene compound, increased in 8 experiments after the fermentation of juices: (1) In Chinese wolfberry juice fermented by either
L. casei,
L. paracasei, L. acidophilus,
L. helveticus, or
B. lactis, the D-limonene concentration was 281.4, 218, 372.6, 271.8, and 273.5 µg/mL, respectively, compared to 143.5 µg/mL in the unfermented juice, where in fermented juice by
L. plantarum, the D-limonene concentration was 67.4 µg/mL [
50]; (2) the D-limonene concentration in distillates prepared using vacuum distillation from
L. rhamnosus fermented melon by-product or unfermented melon by-product was 140 ng/mL and 10 ng/mL, respectively. In the same study, distillates prepared using vacuum distillation had concentrations of 100 ng/mL and 20 ng/mL in the
L. rhamnosus fermented orange pomace distillate and the unfermented pomace distillate, respectively [
44]; (3) in cherry juice fermented by either
L. rhamnosus or
L. paracasei, the D-limonene concentration was 7.9 and 3.0 ng/mL, respectively, compared to 2.8 ng/mL in the unfermented juice, where in cherry juice fermented by four
L. plantarum strains, the D-limonene concentration was 1.8, 1.9, 3.1, and 4.0 ng/mL, compared to 2.3 ng/mL in the unfermented juice [
29]; (4) in jujube juice fermented by a mixture of
L. plantarum,
L. rhamnosus, and
S. thermophilus, the D-limonene concentration was 964 ng/g, where it was not detected in the unfermented juice [
47]; (5) in apple juice fermented by different LAB, the D-limonene concentration ranged from 2.5 to 3.1 ng/g, compared to 1.1 ng/g in the unfermented juice [
21]; (6) in mango slurry fermented by different LAB, the D-limonene concentration was 7.8–10 times higher compared to the concentration in the unfermented slurry, where in yeast fermentation, the D-limonene concentration was 2.2 to 4.5 times lower compared to the concentration in the unfermented juice [
41]; (7) in pomegranate juice fermented by
L. plantarum strains, the D-limonene concentration was increased compared to the concentration in the unfermented juice [
24]; and (8) in tomato juice fermented by
L. casei, D-limonene was detected, where it was not detected after fermentation by
L. plantarum or in the unfermented juice [
32]. However, the D-limonene concentration was reduced in 2 studies after fermentation: (1) The D-limonene concentration in a distillate prepared using simple distillation from
L. rhamnosus fermented orange pomace was 2.3 µg/mL, compared to 4.8 µg/mL in the unfermented pomace distillate [
44]; and (2) in blended apple, orange, carrot, and jujube juices fermented by a mixed starter culture (
L. plantarum,
B. breve, and
S. thermophilus), the D-limonene concentration was 170 ng/mL, compared to 2093 ng/mL in the unfermented blended juice [
35].
In 5 studies, α-terpineol was detected after LAB fermentation: (1) The α-terpineol concentration in a distillate prepared using vacuum distillation from
L. rhamnosus fermented orange pomace was 1.5 µg/mL, compared to 0.3 µg/mL in the unfermented pomace distillate, where it was not detected in distillates prepared using simple distillation. In the same study, using simple distillation, in a distillate of melon by-product fermented by
L. rhamnosus, the α-terpineol concentration was 2.0 µg/mL, compared to 1.0 µg/mL in the unfermented melon by-product distillate. Whereas using vacuum distillation, the α-terpineol concentration was slightly increased to 0.25 µg/mL in the distillate from the fermented melon by-product, compared to 0.18 µg/mL in the unfermented melon by-product distillate [
44]; (2) in kiwifruit juice (Hongyang and Xuxiang cultivars), Hongyang cultivar juice fermented by either
L. acidophilus,
L. helveticus, or
L. plantarum, the α-terpineol concentration was 84.2, 154.9, and 151.1 ng/mL, respectively, where it was not detected in the unfermented juice, and in addition, the α-terpineol was not detected in the unfermented and the fermented Xuxiang cultivar juices [
49]; (3) in goji juice fermented by a bacterial mixture (
L. rhamnosus,
L. reuteri, and
B. velezensis), the α-terpineol concentration was 38.7 ng/g, compared to 33.9 ng/g in the unfermented juice [
25]; (4) in non-pH-adjusted (2.65) bog bilberry juice fermented by two strains of
L. plantarum, the α-terpineol concentration was 5.2 and 5.9 ng/mL, compared to 4.3 ng/mL in the unfermented juice, where the pH of the juice was adjusted to pH 3.5, the initial α-terpineol concentration was 3 ng/mL, which was reduced after fermentation by two strains of
L. plantarum to 2.8 and 2.5 ng/mL [
34]; and (5) in mango slurry fermented by different LAB, α-terpineol was detected, where it was not detected after yeast fermentation or in the unfermented mango slurry [
41].
β-damascenone, another important flavour terpene compound, was detected after fermentation in 4 studies: (1) In Hetain varietal jujube juice fermented by either
L. acidophilus,
L. casei,
L. helveticus, or
L. plantarum, the β-damascenone concentration was 16.7, 22.4, 19.0, and 21.4 µg/mL, respectively, where it was not detected in the unfermented juice [
20]; (2) the β-damascenone concentration in a distillate prepared using simple distillation from a melon by-product fermented by
L. rhamnosus was 8 µg/mL, compared to 1.2 µg/mL in the unfermented melon by-product distillate, and when using vacuum distillation, the β-damascenone concentration was 0.03 µg/mL in unfermented melon by-product distillate and 0.12 µg/mL in the distillate of
L. rhamnosus fermented melon by-product [
44]; (3) in goji juice fermented by different combinations of mixed bacterial cultures, the β-damascenone concentration ranged from 39.6 to 52 ng/g, where it was not detected in the unfermented juice [
25]; and (4) in apple juice fermented by different LAB, the β-damascenone concentration ranged from 2.4 to 3.9 ng/mL, where it was not detected in the unfermented juice [
19]. However, Chen et al. [
21] reported in apple juice that the initial β-damascenone concentration was 12.8 ng/g and that after fermentation with LAB, it was not detected.
Eugenol is a volatile phenol that imparts clove, honey, and spicy notes to fermented fruit and vegetable juices. In the three papers, where it was mentioned, it was classified as a terpene: (1) In horse gram sprouts fermented by two
L. plantarum strains, the eugenol concentration was 5.2 and 5.3 µg/g, and it was not detected in raw seeds [
37]; (2) in cherry juice fermented by either
L. paracasei or
L. rhamnosus, the eugenol concentration was 3.9, and 26.8 ng/mL, respectively, compared to 2.4 ng/mL in the unfermented juice, and cherry juice fermented by four
L. plantarum strains, the eugenol concentration was 11.1, 13.0, 14.7, and 22.4 ng/mL, compared to 3.2 ng/mL in the unfermented juice [
29]; and (3) in apple juice fermented by either
L. plantarum,
L. rhamnosus,
L. casei, or
L. acidophilus, the eugenol concentration was 9.1, 6.2, 17.1, and 33.3 ng/g, respectively, compared to 5.4 ng/g in the unfermented juice [
21].
In addition, geraniol was detected in 3 studies after fermentation: (1) In watermelon juice fermented by either
L. plantarum,
L. brevis,
P. pentosaceus, L. casei, or
L. rhamnosus, the geraniol concentration was 5.6, 1.7, 1.7, 1.5, and 1.3 ng/mL, respectively, compared to 1.0 ng/mL in the unfermented juice [
48]; (2) in goji juice fermented by bacterial mixture, the geraniol concentration ranged from 119.3–246 ng/g, where it was not detected in the unfermented juice [
25]; and (3) in apple juice fermented by either
L. plantarum,
L. rhamnosus,
L. casei, or
L. acidophilus, the geraniol concentration was 15.9, 13.2, 14.4, and 14.3 ng/g, respectively, compared to 5.5 ng/g in the unfermented juice [
21].
Overall, a wide range of terpenes were found to increase after LAB fermentation of fruit and vegetable juices, with D-limonene being the most commonly reported (8 papers) terpene, followed by linalool (7 papers), α-terpineol (5 papers), β-damascenone (4 papers), eugenol (3 papers), and geraniol (3 papers). However, in some instances after fermentation, the concentration of linalool (2 papers), D-limonene (2 papers), and β-damascenone (1 paper) decreased.








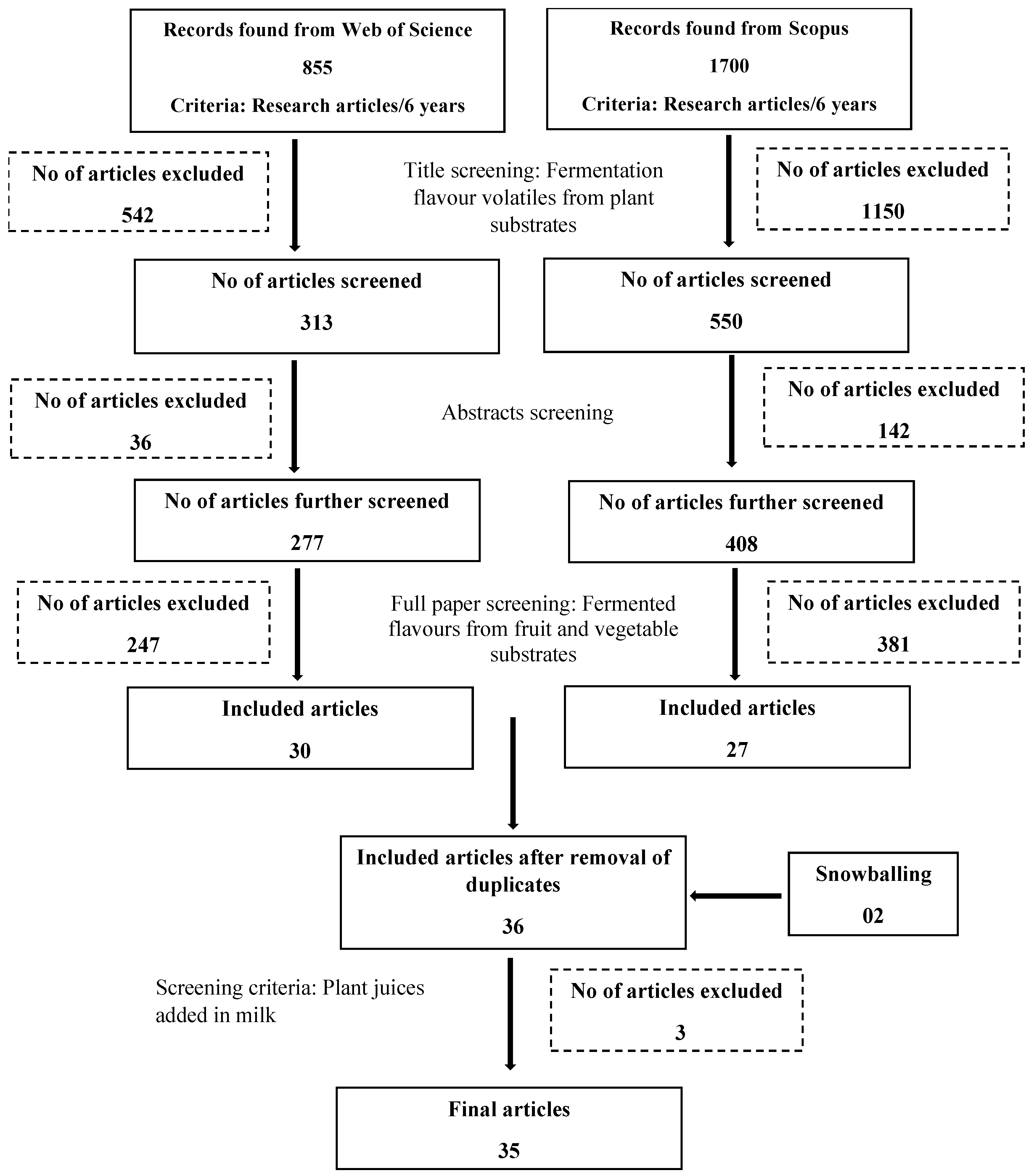{kind=link}
{kind=link}
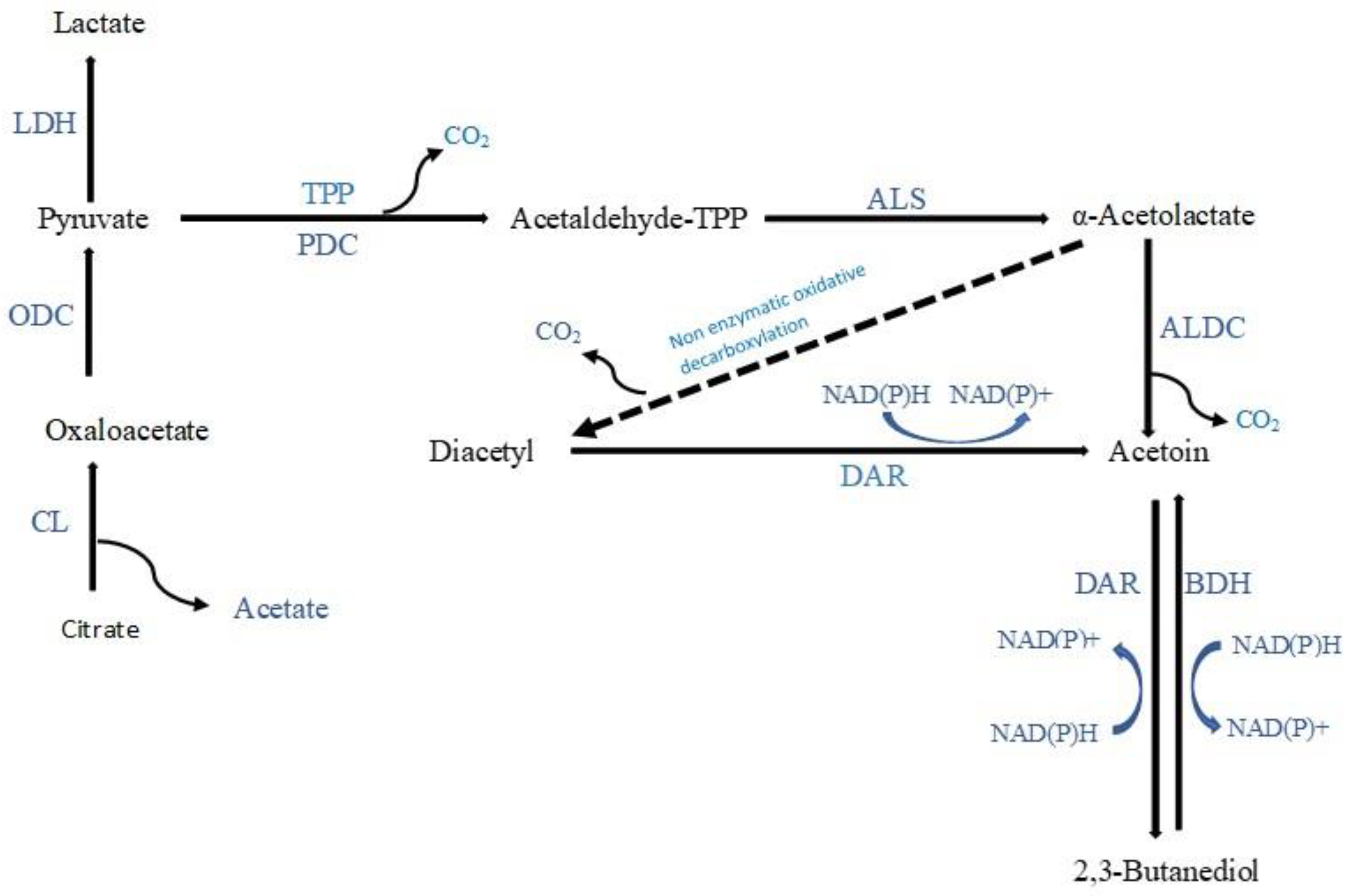{kind=link}
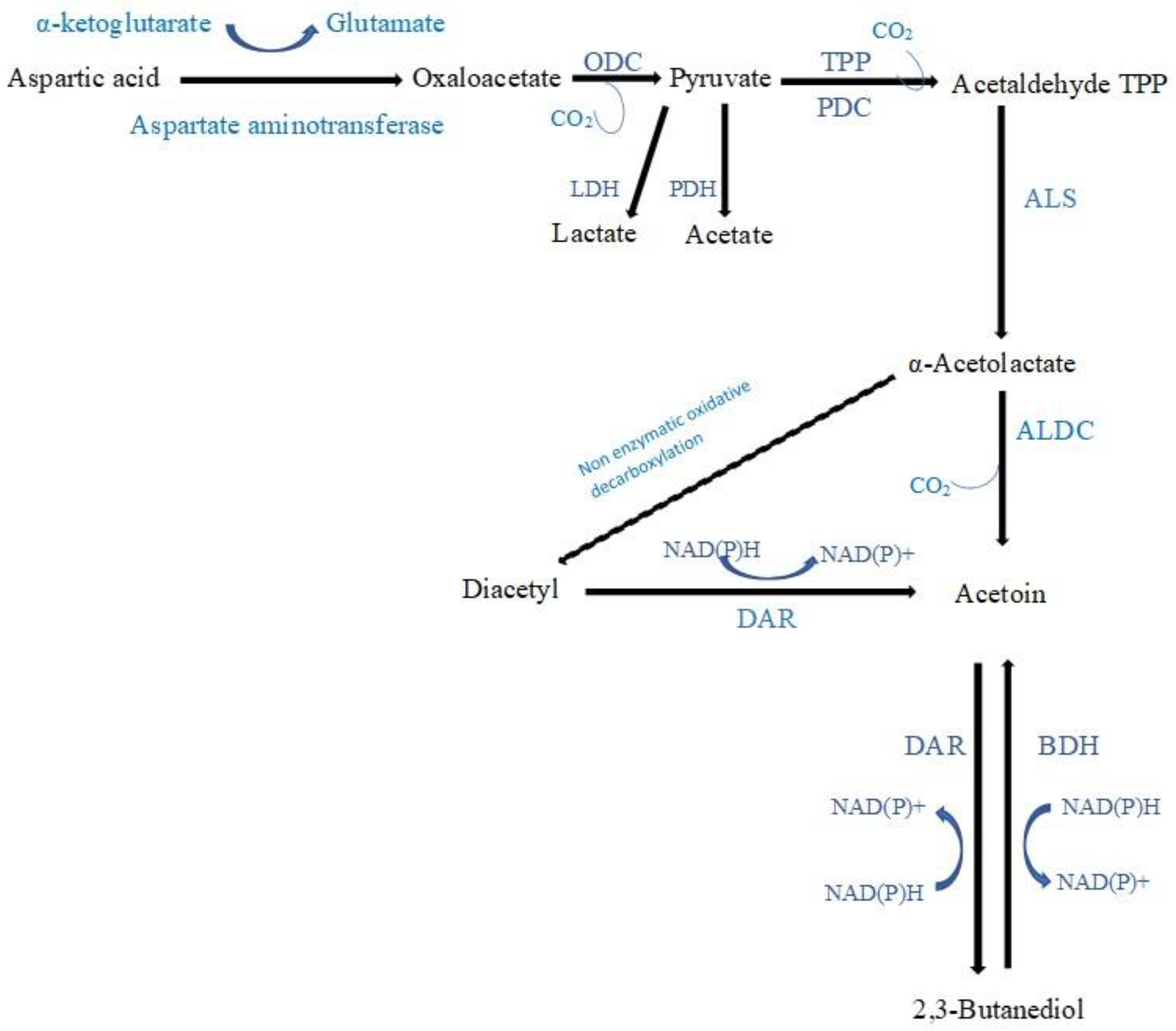{kind=link}