
Recent Advances in the Nutritional Value, Chemical Compositions, Pharmacological Activity, and Application Value of Orychophragmus violaceus: A Comprehensive Review

Abstract
:
1. Introduction
2. Research Methodology
3. Nutritional Value
3.1. Vitamins
3.2. Amino Acid
3.3. Mineral Element
3.4. Fatty Acids
3.5. Others
4. Chemical Compositions
4.1. Flavonoids
4.2. Alkaloids
4.3. Phenylpropanoids
4.4. Phenolic Acid
4.5. Terpenoids
4.6. Steroids
4.7. Anthocyanidin
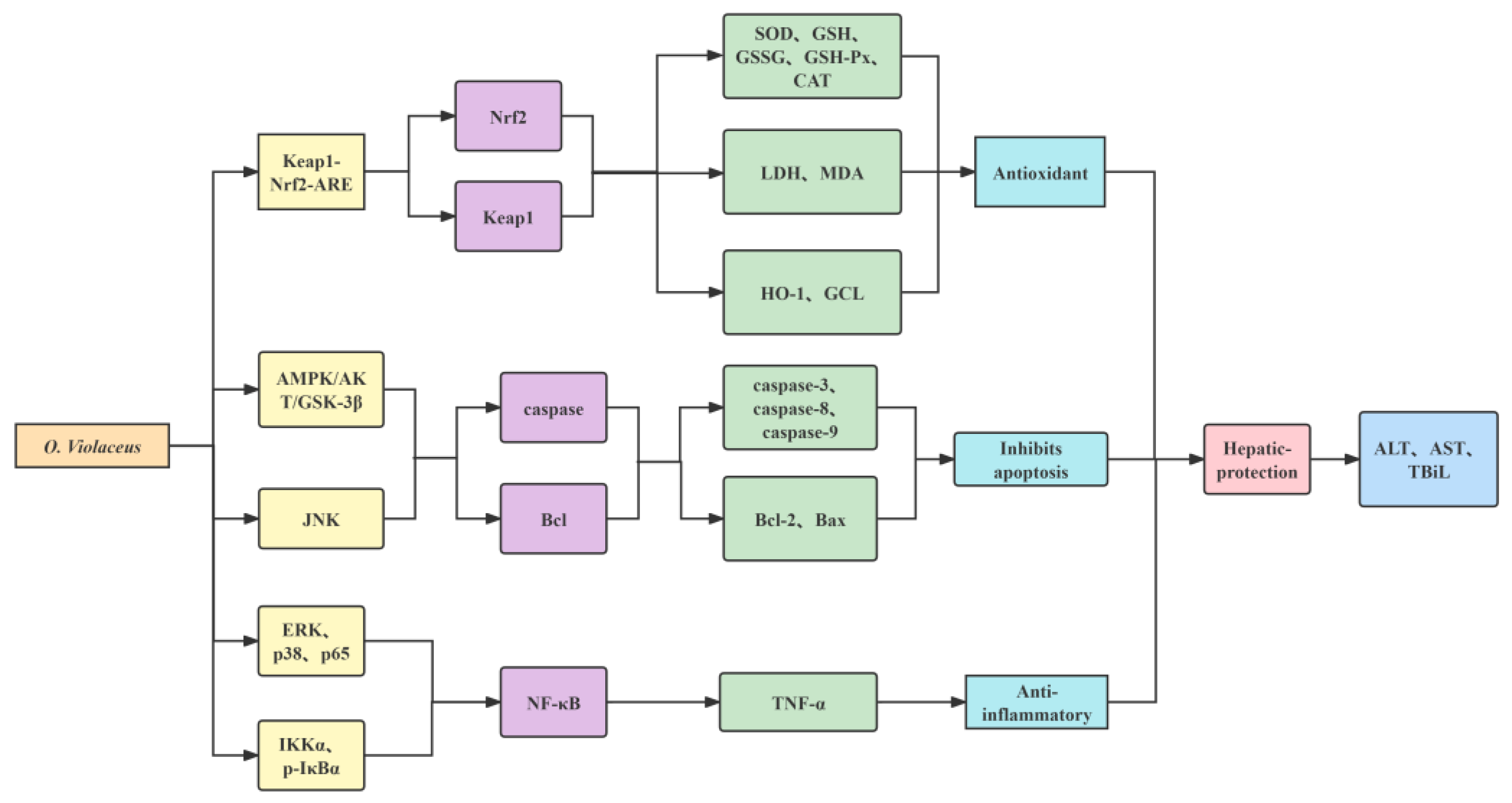
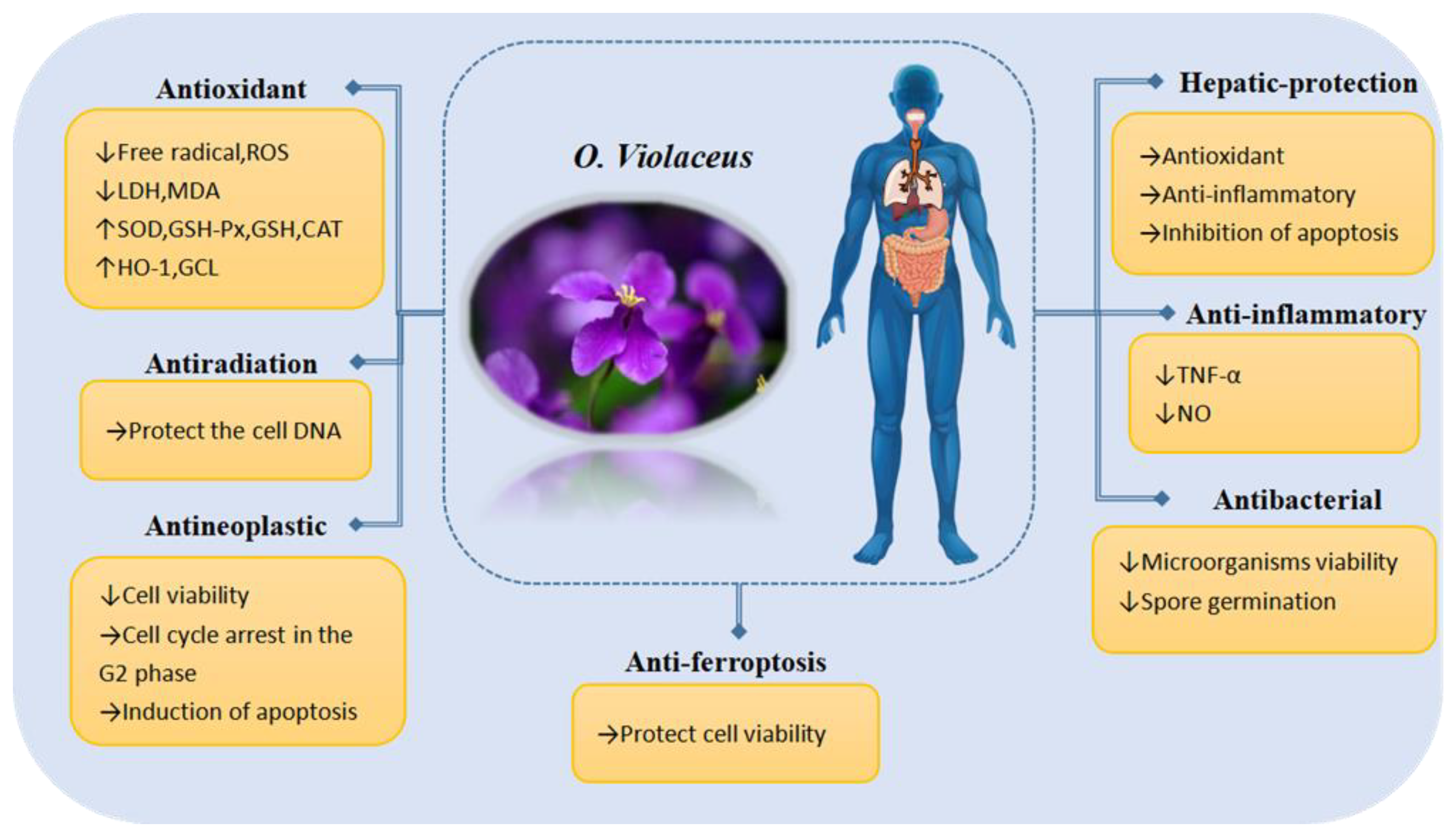
5. Pharmacological Activity
5.1. Antioxidant Properties
5.2. Antiradiation Properties
5.3. Antineoplastic Properties
5.4. Hepaticprotective Properties
5.5. Antiferroptosis Properties
5.6. Anti-Inflammatory Properties
5.7. Antibacterial Properties
6. Toxicity
7. Application Value
7.1. Ornamental Value
7.2. Ecological Value
7.3. Feeding Value
7.4. Edible Value
7.5. Medicinal Value
8. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, H.; Wang, Q.; Liu, J.-Q. Recent Advances and Future Prospects of the Important Resource Plant Orychophragmus. Plant Sci. J. 2014, 32, 189–198. [Google Scholar]
- Chen, Z.; Lu, Y.; Hu, J.; Zhang, Y.; Jin, X. Taxonomic study Orychophragmus Bunge (Brassicaceae), agenus endemic to China. J. Zhejiang Univ. Sci. Ed. 2017, 44, 201–205. [Google Scholar]
- Chen, B.; Ren, Q. Characteristics and application value of Orychophragmus violaceus. Mod. Agric. Sci. Technol. 2015, 9, 169. [Google Scholar]
- Chen, X.; Mao, L.; Pan, Y.; Wang, Y. Comparison on pollen morphological characteristics of some species of Brassicaceae. J. Plant Resour. Environ. 2022, 31, 13–20. [Google Scholar]
- Li, M.; Zhao, H. Pod Development Regularity of Orychophragmus violaceus. J. Anhui Sci. Technol. 2018, 32, 18–23. [Google Scholar]
- Zhang, L.Q. Study on the shade tolerance of Orychophragmus violaceus. Contemp. Hortic. 2006, 8, 42. [Google Scholar]
- Ni, S.; Cheng, Z.; Gong, J.; Luo, R.; Li, Z.; Cui, Y.; Zhao, G. Research Overview on Orychophragmus spp. Plant. J. Anhui Agric. Sci. 2009, 37, 6435–6460. [Google Scholar]
- Luo, P.; Li, Y.; Huang, B. A Desirable Genrtic Resource of Oil Plant-Orychophragmus violaceus. Chin. J. Oil Crop Sci. 1993, 4, 75–77. [Google Scholar]
- Zhang, T.-T.; Ma, G.-X.; Xu, F.-Q.; Wu, J.-N.; Zhang, X.; Yang, J.-S.; Zhang, B.-X.; Xu, X.-D. Water Soluble Constituents from of Seeds Orychophragmus violaceus. Chin. Pharm. J. 2014, 49, 2165–2167. [Google Scholar]
- Wang, X.; Li, X.; Wang, H. Study on the Extraction Method of Flavonoid Compounds in Orychophragmus violaceus and its Antioxidation. J. Anhui Agric. Sci. 2007, 35, 8667–8669. [Google Scholar]
- Liu, S.; Zhang, G.; Cui, H.; Qin, H.; Dong, J. Chemical constituents from the seeds of Orychophragmus violaceus. J. Int. Pharm. Res. 2018, 45, 238–243. [Google Scholar]
- Zhu, N.; Wu, H.; Xu, Z.; Liu, C.; Tian, Y.; Hu, M.; Sun, Z.H.; Li, P.; Ma, G.; Xu, X. New alkaloids with unusual spermidine moieties from the seeds of Orychophragmus violaceus and their cytoprotective properties. RSC Adv. 2017, 7, 41495–41498. [Google Scholar] [CrossRef]
- Zhang, G.J.; Li, B.; Chen, L.; Tian, Y.; Liu, S.J.; Cui, H.M.; Dong, J.X. Isocoumarin derivatives and monoterpene glycoside from the seeds of Orychophragmus violaceus. Fitoterapia 2018, 125, 111–116. [Google Scholar] [CrossRef]
- Xia, X.Y. Study on Ployphenols and Quality Control of the Seeds of Orychophragmus violaceus; Harbin University of Commerce: Harbin, China, 2019. [Google Scholar]
- Zhang, G.; Cui, H.; Liu, S.; Dong, J. A new triterpenoid saponin and a diterpene glucoside from the seeds of Orychophragmus violaceus. Nat. Prod. Res. 2019, 33, 407–413. [Google Scholar] [CrossRef]
- Luo, P.; Huang, B.; Ye, D.; Gao, H.; Chen, Y.; Mu, Z. Study on the forage plant resources of Orychophragmus violaceus. Acta Pratacult. Sin. 1998, 7, 54–58. [Google Scholar]
- Li, X.; He, S.; Ren, B.; Sheng, N. Nutrient constituents of Orychophragmus violaceus (L.) O. E. Schulz and the evaluation as a wild vegetable. J. Plant Resour. Environ. 1997, 6, 9–13. [Google Scholar]
- Luo, P.; Huang, B.; Lan, Z.; Yan, H. A Study on the Vegetable Resource Orychophragmus violaceus. J. Sichuan Univ. Nat. Sci. Ed. 1998, 35, 148–151. [Google Scholar]
- Schjoldager, J.G.; Paidi, M.D.; Lindblad, M.M.; Birck, M.M.; Kjærgaard, A.B.; Dantzer, V.; Lykkesfeldt, J.; Tveden-Nyborg, P. Maternal vitamin C deficiency during pregnancy results in transient fetal and placental growth retardation in guinea pigs. Eur. J. Nutr. 2015, 54, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Tveden-Nyborg, P. The Pharmacokinetics of Vitamin C. Nutrients 2019, 11, 2412. [Google Scholar] [CrossRef] [PubMed]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [PubMed]
- Weng, D.; Huang, X. Evaluation on the protein quality of Orychophragmus violaceus. Acta Bot. Boreali-Occident. Sin. 2001, 21, 673–677. [Google Scholar]
- Wang, X.; Xu, G. Study on Protein Extraction Technology from Iolet Orychophragmus. J. Anhui Agric. Sci. 2009, 37, 9128–9129. [Google Scholar]
- Cao, W.; Rui, Y.; Li, X. Determination of Forty-Six Elements in Different Organs of Orychophragmus violaceus in Agricultural Farm. Asian J. Chem. 2014, 26, 1038–1040. [Google Scholar] [CrossRef]
- Min, Z. Study on the Extraction of Orychophragmus violaceus Seed Oil and the Preparation of Its Microencapsulation; Guizhou University: Guiyang, China, 2018. [Google Scholar]
- Qi, P.; Li, Z. Genome and Genetics of the Crucifer Orychophragmus violaceus Native to China. J. Plant Genet. Resour. 2023, 24, 340–348. [Google Scholar]
- Zhao, R.; Yang, X.; Zhang, A.; Zhou, T.; Zhou, Y.; Yang, L. An efficient approach for simultaneously obtaining oil and epigoitrin from Orychophragmus violaceus seeds by microwave-mediated immiscible binary solvent extraction. Food Chem. 2022, 372, 131258. [Google Scholar] [CrossRef]
- Sun, X.; Peng, H.; Guo, J.; Peng, B.; Bai, M.; Hang, Y. Fatty Acid Analysis of the Seed Oil in a Germplasm Collection of 94 Species in 58 Genera of Brassicaceae. Chem. Ind. For. Prod. 2011, 31, 46–54. [Google Scholar]
- Xue, Y.; Wang, Z.; Hao, Y.; Deng, M.; Xue, J. Analysis of Fatty Acids of Different Growth Stages in Orychophragmus violaceus Seeds. J. Shanxi Agric. Sci. 2020, 48, 715–718. [Google Scholar]
- Lv, Z. Analysis and utilization evaluation of oil and fat from seeds of Orychophragmus violaceus. Chin. Wild Plant Resour. 1992, 1, 1–5. [Google Scholar]
- Liu, S.; Xu, Z.; Zhang, G.; Dong, J. Lignan derivatives and a jasmonic acid derivative from the seeds of Orychophragmus violaceus. Nat. Prod. Res. 2022, 36, 3779–3785. [Google Scholar]
- Hu, Q.; Hansen, N.; Laursen, J.; Dixelius, C.; Andersen, B. Intergeneric hybrids between Brassica napus and Orychophragmus violaceus containing traits of agronomic importance for oilseed rape breeding. Theor. Appl. Genet. 2002, 105, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Chen, P.; Tang, X.; Zhong, T.; Yang, T.; Nwafor, C.C.; Yang, C.; Ge, X.; An, H.; Li, Z.; et al. Genome assembly of the Brassicaceae diploid Orychophragmus violaceus reveals complex whole-genome duplication and evolution of dihydroxy fatty acid metabolism. Plant Commun. 2023, 4, 100432. [Google Scholar] [CrossRef] [PubMed]
- Romsdahl, T.; Shirani, A.; Minto, R.E.; Zhang, C.; Cahoon, E.B.; Chapman, K.D.; Berman, D. Nature-Guided Synthesis of Advanced Bio-Lubricants. Sci. Rep. 2019, 9, 11711. [Google Scholar] [CrossRef] [PubMed]
- Hao, J. Study on the Difference of Di-hydroxy Fatty Acid Composition in the Seeds of Orychophragmus Limprichtianus and Orychophragmus violaceus; Huazhong Agricultural University: Wuhan, China, 2019. [Google Scholar]
- Petrussa, E.; Braidot, E.; Zancani, M.; Peresson, C.; Bertolini, A.; Patui, S.; Vianello, A. Plant Flavonoids-Biosynthesis, Transport and Involvement in Stress Responses. Int. J. Mol. Sci. 2013, 14, 14950–14973. [Google Scholar] [CrossRef] [PubMed]
- Weng, D.; Wang, H.; Weng, J. Study on flavonoids in leaves and stalks of Orychophragmus violaceus. Chin. Wild Plant Resour. 2000, 19, 13–15. [Google Scholar]
- Wang, X.; Xia, X.; Xu, G. Research on Extraction of Total Isoflavones in Oryehophragmus violaceus by Microwave-assisted. J. Anhui Agric. Sci. 2007, 35, 6705–6706. [Google Scholar]
- Xin, W. Total Flavonoids Extraction, Purification and Oxidation Resistance of Orychophragmus; Northwest Agriculture and Forestry University: Xi’an, China, 2008. [Google Scholar]
- Sai, C.; Cui, L.; Yang, M.; Zhou, M.; Jin, X.; Wang, J. Optimization of ultrasound assisted extraction technology of total flavonoids from orychophragmus violaceus by response surface method and study on the antioxidant activity. J. Jining Med. Univ. 2018, 41, 53–58. [Google Scholar]
- Zhang, G. Studies on the Chemical Constituments and Bioactivities from the Seeds of Orychophragmus violaceus; Academy of Military Sciences: Beijing, China, 2018. [Google Scholar]
- Xu, Z. Study on the Chemical Compositions and Bioactivities of Orychophragmus violaceus. Master’s Thesis, Guangdong Pharmaceutical University, Guangzhou, China, 2022. [Google Scholar]
- Huo, X.; Liu, C.; Gao, L.; Xu, X.; Zhu, N.; Cao, L. Hepatoprotective Effect of Aqueous Extract from the Seeds of Orychophragmus violaceus against Liver Injury in Mice and HepG2 Cells. Int. J. Mol. Sci. 2017, 18, 1197. [Google Scholar] [CrossRef] [PubMed]
- Chen, L. Studies on Antiviral Effect and Antibacterial Effect of Rdix Isatidis Alkaloid In Vitro; Henan Agricultural University: Zhengzhou, China, 2010. [Google Scholar]
- Zhang, H. Neurotoxicity and Neuroprotection as well as Metabolic Differences of Gelsemium Alkaloids; Northwest Agriculture and Forestry University: Xi’an, China, 2022. [Google Scholar]
- Liu, C.-Q.; Zhu, N.-L.; Yang, S.-X.; Dai, Y.-H.; Li, L.-Y.; Xu, X.-D.; Cao, L. Protective Effect of Orychophragmuspine I Against Oxidative Damage in HepG2 Cells Induced by Hydrogen Peroxide. Mod. Food Sci. Technol. 2017, 33, 19–25. [Google Scholar]
- Zhang, G.J.; Li, B.; Cui, H.M.; Chen, L.; Tian, Y.; Liu, S.J.; Li, B.W.; Li, M.; Xia, Z.M.; Chen, X.X.; et al. Orychophragines A-C, Three Biologically Active Alkaloids from Orychophragmus violaceus. Org. Lett. 2018, 20, 656–659. [Google Scholar] [CrossRef]
- Zhang, G.; Dong, J.; Liu, S.; Li, B.; Tian, Y.; Li, M. Application of Orychophragines D in the Preparation of Drugs against Radiation Damage. China Patent CN113262229B, 24 February 2023. [Google Scholar]
- Xia, X.; Zhu, N.; Ji, Y.; Xu, X. Determination of epigoitrin in seeds of Orychophragmus violaceus by UPLC. Sci. Technol. Innov. 2018, 31, 10–11. [Google Scholar]
- Xu, Z.X.; Li, B.; Tian, Y.; Li, M.; Dong, J.X.; Zhang, G.J. Pentacyclic spermidine alkaloids with radioprotective and anti-inflammatory activities from Orychophragmus violaceus. Org. Biomol. Chem. 2021, 19, 9844–9848. [Google Scholar] [CrossRef]
- Xu, Z.; Bin, L.; Ying, T.; Min, L.; Dong, J.; Zhang, G. Chemical constituents from the aerial parts of Orychophragmus violaceus. Nat. Prod. Res. 2022, 38, 278–286. [Google Scholar] [CrossRef]
- Zhang, G.; Sang, T.; Chen, X.; Ge, C.; Li, B.; Tian, Y.; Li, M.; Liu, S.; Xia, Z.; Li, H.; et al. Orychophragine D:A new 2-piperazinone fused 5-azacytosine type alkaloid with radioprotective activity from the seeds of Orychophragmus violaceus. Fitoterapia 2023, 168, 105544. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Yang, W.; Xu, Z.; Liu, C.; Wu, H.; Ma, G.; Xu, X. Study on the alkaloidal components of Orychophragmus violaceus. In Proceedings of the 2016 First China Traditional Chinese Medicine Resources Conference, Xi’an, China, 26 October 2016; pp. 362–363. [Google Scholar]
- Kong, L.-L.; Hu, J.-F.; Chen, N.-H. Advances in pharmacology and toxicology of coumarins. Chin. Pharmacol. Bull. 2012, 28, 165–168. [Google Scholar]
- Ye, C.; Chen, J.; Wu, L.; Liu, C. Advances in the study of the anti-inflammatory activity of coumarins. J. Mod. Med. Health 2022, 38, 4063–4065+4069. [Google Scholar]
- Yang, Q.; Chen, M. Research progress in the prevention and treatment of cardiovascular diseases by regulating intestinal flora with phenolic acids. Chin. Tradit. Pat. Med. 2022, 44, 3920–3926. [Google Scholar]
- Wang, J.; Ding, H. Research progress on bacteriostatic effects of phenolic acids. Chin. Tradit. Pat. Med. 2022, 44, 1906–1911. [Google Scholar]
- Wan, X.; Wang, Y.; Zhou, C.; Guo, H.; Ma, S. Research progress on chemical constituents and pharmacological effects of Salvia miltiorrhiza. Chin. Tradit. Herb. Drugs 2020, 51, 788–798. [Google Scholar]
- Wei, C.; Cui, P.; Xiao, J.; Luo, X.; Jiang, S. Research progress of natural diterpenoids with antibacterial activity. Chin. Tradit. Pat. Med. 2022, 44, 2240–2249. [Google Scholar]
- Zhou, Q. Research on the Vasorelaxant Substances, Novel Terpenoids from the Aerial Parts of Pogostemon cablin and Leonurus japonicus; Chengdu University of Traditional Chinese Medicine: Chengdu, China, 2017. [Google Scholar]
- Chen, L.; Wang, L.; Yang, J. Research Progress on Diterpenoids and Their Biological Activities in Rubus Plants. Mod. Chin. Med. 2022, 24, 2040–2047. [Google Scholar]
- Wang, B.; Gao, L.; Wang, J.; Zhang, J.; Sun, X. Establishment of a liquid chromatographic method for determination of the content of steroidal saponin 25-epi-officinalisninⅡin Asparagi Radix. J. Int. Pharm. Res. 2018, 45, 799–803. [Google Scholar]
- Liu, L. Study on STeroidal AAlkaloids and Antitumor Effect of Solanum nigrum and Solanum lyratum; China Academy of Chinese Medical Sciences: Beijing, China, 2022. [Google Scholar]
- Zhao, Q.; Xie, B.; Yan, J.; Zhao, F.; Xiao, J.; Yao, L.; Zhao, B.; Huang, Y. In Vitro antioxidant and antitumor activities of polysaccharides extracted from Asparagus officinalis. Carbohydr. Polym. 2012, 87, 392–396. [Google Scholar] [CrossRef]
- Honda, T.; Tatsuzawa, F.; Kobayashi, N.; Kasai, H.; Nagumo, S.; Shigihara, A.; Saito, N. Acylated anthocyanins from the violet-blue flowers of Orychophragonus violaceus. Phytochemistry 2005, 66, 1844–1851. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Liu, W.; Wang, J.; Gu, C.; Wan, Y.; Wang, J. Tracing carbonation in cementitious slurry using anthocyanin extracted from fresh or stale petals. RSC Adv. 2022, 12, 32557–32566. [Google Scholar] [CrossRef]
- Jia, X.; Deng, Y.; Sun, X.; Liang, L.; Xiao, Z. Anthocyanin Profiles in Petals of Orychophragonus violaceus. Chin. Agric. Sci. Bull. 2018, 34, 60–64. [Google Scholar]
- Demirci-Çekiç, S.; Özkan, G.; Avan, A.N.; Uzunboy, S.; Çapanoğlu, E.; Apak, R. Biomarkers of Oxidative Stress and Antioxidant Defense. J. Pharm. Biomed. Anal. 2022, 209, 114477. [Google Scholar] [CrossRef] [PubMed]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-inflammatory effects of flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef]
- Hui, G. Molecular Mechanism of Quercetin and Catechin in Protecting Cell against Oxidative Stress by Targeting FOXO3 to Synergistically Inhibit CHUK Transcription; Shandong Agriculutural University: Taian, China, 2022. [Google Scholar]
- Li, G.; Yang, S. Extraction of Codonopsis Pilosula Polysaccharide and Its Effect of Antiactive Oxygen Free Radicals. Chem. World 2001, 8, 421–422+434. [Google Scholar]
- Calvo-Asensio, I.; Barthlott, T.; von Muenchow, L.; Lowndes, N.F.; Ceredig, R. Differential Response of Mouse Thymic Epithelial Cell Types to Ionizing Radiation-Induced DNA Damage. Front. Immunol. 2017, 8, 418. [Google Scholar] [CrossRef]
- Arora, R.; Gupta, D.; Chawla, R.; Sagar, R.; Sharma, A.; Kumar, R.; Prasad, J.; Singh, S.; Samanta, N.; Sharma, R.K. Radioprotection by plant products: Present status and future prospects. Phytother. Res. PTR 2005, 19, 1–22. [Google Scholar] [CrossRef]
- Liu, L.; Chen, M.; Zhao, H.; Yao, L. Recent Advances in Understanding the Radioprotective Effect of Natural Products. Food Sci. 2018, 39, 269–274. [Google Scholar]
- Zhou, A.; Wang, D.; Yue, X.; Zhou, C.; Huang, L. Anti-tumor Effect of Chinese Herbal Polysaccharides: A Review. Chin. J. Exp. Tradit. Med. Formulae 2022, 28, 236–246. [Google Scholar]
- Niu, C.; Zhang, L.; Huang, J. Research Progress on Antitumor Mechanism of Traditional Chinese Medicine. J. Liaoning Univ. Tradit. 2023, 25, 132–136. [Google Scholar]
- Xia, X.; Cole, S.P.C.; Cai, T.; Cai, Y. Effect of traditional Chinese medicine components on multidrug resistance in tumors mediated by P-glycoprotein. Oncol. Lett. 2017, 13, 3989–3996. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, L. Expert consensus on liver inflammation and its prevention and treatment. Chin. J. Pract. Intern. Med. 2014, 34, 152–162. [Google Scholar]
- Hamdy, N.; El-Demerdash, E. New therapeutic aspect for carvedilol: Antifibrotic effects of carvedilol in chronic carbon tetrachloride-induced liver damage. Toxicol. Appl. Pharmacol. 2012, 261, 292–299. [Google Scholar] [CrossRef]
- Gu, X.; Manautou, J.E. Molecular mechanisms underlying chemical liver injury. Expert Rev. Mol. Med. 2012, 14, e4. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.; Wang, F.-f.; Yang, S.; Li, L.; Cao, L. Protective effect of water extracts of Orychophragmus violaceus seeds on TAA-induced acute liver injury in mice. China J. Chin. Mater. Med. 2020, 45, 1399–1405. [Google Scholar]
- da Rocha, B.A.; Ritter, A.M.; Ames, F.Q.; Gonçalves, O.H.; Leimann, F.V.; Bracht, L.; Natali, M.R.; Cuman, R.K.; Bersani-Amado, C.A. Acetaminophen-induced hepatotoxicity: Preventive effect of trans anethole. Biomed. Pharmacother. 2017, 86, 213–220. [Google Scholar] [CrossRef]
- Pang, M.; Shan, Y.; Wang, F.-f.; Yang, S.; Li, L.; Cao, L. Protective Effect of Water Extracts of Orychophragmus violaceus Seeds on Acetaminophen-induced Acute Liver Injury in Mice: Roles of Activating Hepatic AMPK/AKT/GSK-3β Signaling. Mod. Food Sci. Technol. 2020, 36, 34–41. [Google Scholar]
- Xie, J.; Han, Z.; Yun, W. Protective effect of salvianolate on ConA-induced immune liver injury in mice. J. Chin. Med. Mater. 2017, 40, 2686–2688. [Google Scholar]
- Gong, F.; Shen, Y.; Zhang, C.; Xu, J.; Wu, X.; Hua, Z.; Xu, Q. Dregea volubilis ameliorates concanavalin A-induced liver injury by facilitating apoptosis of activated T cells. Exp. Biol. Med. 2008, 233, 1124–1132. [Google Scholar] [CrossRef] [PubMed]
- Min, P. The Molecular Mechanism of Water Extracts from Orychophragmus violaceus Seed on Liver Injury; Harbin University of Commerce: Harbin, China, 2020. [Google Scholar]
- Hagymási, K.; Blázovics, A.; Lengyel, G.; Kocsis, I.; Fehér, J. Oxidative damage in alcoholic liver disease. Eur. J. Gastroenterol. Hepatol. 2001, 13, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Bataller, R. Alcoholic liver disease: Pathogenesis and new therapeutic targets. Gastroenterology 2011, 141, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-Q.; Gao, L.; Huo, X.-w.; Zhu, N.; Dai, Y.; Yang, S.; Li, L.; Cao, L. Protective Effect of Water Extracts of Orychophragmus violaceus Seeds on Alcohol-induced Acute Liver Injury in Mice. Mod. Food Sci. Technol. 2017, 33, 7–11+79. [Google Scholar]
- Wang, L.; Li, Z.; Li, L.; Li, Y.; Yu, M.; Zhou, Y.; Lv, X.; Arai, H.; Xu, Y. Acute and sub-chronic oral toxicity profiles of the aqueous extract of Cortex Dictamni in mice and rats. J. Ethnopharmacol. 2014, 158 Pt A, 207–215. [Google Scholar] [CrossRef]
- Zhan, Y.-w.; Xu, Z.-Q.; Guo, X.-h.; Li, R.; Shen, J.; Xu, X.; Zhang, B. Protective effect of Orychophragmus violaceus seed against acute liver injury induced by Cortex Dictamni in mice. Chin. J. Pharmacol. Toxicol. 2016, 30, 101–106. [Google Scholar]
- Sun, W.; Lu, Y. Progress incocaine toxicology research. Chin. Pharmacol. Bull. 2004, 11, 1212–1214. [Google Scholar]
- Ndikum-Moffor, F.M.; Roberts, S.M. Cocaine-protein targets in mouse liver. Biochem. Pharmacol. 2003, 66, 105–113. [Google Scholar] [CrossRef]
- Xu, Z.; Li, R.; Shen, J.; Zhu, N.; Zhang, B. Protective effect of Orychophragmus violaceus on cocaine-induced acute liver injury in mice. J. Toxicol. 2017, 31, 51–54+59. [Google Scholar]
- Hang, L. The Role of Nrf2 in CCl4 Induced Liver Fibrosis and Related Mechanisms. Ph.D. Thesis, China Medical University, Shenyang, China, 2021. [Google Scholar]
- Schuppan, D.; Kim, Y.O. Evolving therapies for liver fibrosis. J. Clin. Investig. 2013, 123, 1887–1901. [Google Scholar] [CrossRef]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Zhang, G.; Du, L.; Lu, Y.; Chen, X.; Dong, J.; Wang, L. Application of Orychophragine D in the Preparation of Drugs to Inhibit Ferroptosis in IEC6 Cells. China Patent CN114028404A, 11 February 2022. [Google Scholar]
- Li, J.; Li, Y.; Lu, R.; Wei, J.; Liao, G. Research Progress on Anti-Inflammatory Alkaloids. Guangzhou Chem. Ind. 2020, 48, 14–19. [Google Scholar]
- Zhang, W.; Yuan, S.; Wang, C.; Ye, X. Extraction of antifungal protein from Orychophragmus violaceus seeds and its antifungal activity. J. Fujian Agric. For. Univ. Nat. Sci. Ed. 2019, 48, 296–302. [Google Scholar]
- Zhang, M.; Jia, F.; Zhang, B. Studies on acute and subacute toxicity of Orychophragmus violaceus. In Proceedings of the 4th Annual Conference of Chinese Society of Toxicology on Toxicology and Safety Evaluation of Traditional Chinese Medicine and Natural Medicine, Haikou, China, 15 November 2019; p. 171. [Google Scholar]
- Ma, M.; Mei, Y. Research Status and Development Prospect of Orychophragmus violaceus. J. Anhui Agric. 2012, 40, 5109–5111+5113. [Google Scholar]
- Hongmei, J. Discussion on cultivation technique and landscape application of Orychophragmus violaceus. Agric. Technol. 2018, 38, 218. [Google Scholar]
- Shi, Y.; Liang, F.; Ji, Y.; Ji, Y.; Ma, N.; Ye, S. The application value of Orychophragmus violaceus in garden. Sci. Technol. Tianjin Agric. 2011, 2, 18–20. [Google Scholar]
- Zhang, Z.; An, J.; Han, Y.; Feng, L.; Li, X.; Xiong, S.; Xing, F.; Xin, M.; Li, Y.; Wang, Z. Advantages of an Orychophragmus violaceus-maize rotation in reducing greenhouse gas emissions and reactive nitrogen losses and increasing net ecosystem economic benefits on the North China Plain. J. Clean. Prod. 2021, 317, 128426. [Google Scholar] [CrossRef]
- Wu, D. Studies on Bio-Ecology of Landscape Cover Green Plant Orychophragmus violaceus; Hunan Agricultural University: Changsha, China, 2010. [Google Scholar]
- Jia, L. Study on Nutrition Charracteristics and Green Manure Effect of Orychophragmus violaceus; Chinese Academy of Agricultural Sciences: Beijing, China, 2010. [Google Scholar]
- Daohong, Z. Application value and intensive planting management technology of Orychophragmus violaceus. Mod. Agric. Sci. Technol. 2018, 24, 74–75. [Google Scholar]
- Cui, W.; Ma, H.; Kong, L. Research progress on Nrf2/ARE pathway and mechanism of antioxidation. J. Jilin Univ. Med. Ed. 2011, 37, 187–190. [Google Scholar]
- Lu, C.; Xu, W.; Zheng, S. Nrf2 activation is required for curcumin to induce lipocyte phenotype in hepatic stellate cells. Biomed. Pharmacother. 2017, 95, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.-f.; Wang, Y.; Ren, R.-j.; Huo, H.; Sun, J. Anti-oxidative stress actions and regulation mechanisms of Keap1-Nrf2/ARE signal pathway. J. Int. Pharm. Res. 2016, 43, 146–152+166. [Google Scholar]
- Tang, L. The Research on the Mechanism of Kuijieling to the NF-κB Activation of IκBα Phosphorylation/Ubiquitin-Proteasome System; Guangzhou University of Chinese Medicine: Guangzhou, China, 2012. [Google Scholar]
- Tang, J.; Xu, L.; Zeng, Y.; Gong, F. Effect of gut microbiota on LPS-induced acute lung injury by regulating the TLR4/NF-kB signaling pathway. Int. Immunopharmacol. 2021, 91, 107272. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Wang, W.; He, Z.; Chen, X.; Liu, Z.; Tao, X.; Zhao, P.; Yang, Z.; Zhao, H. Anti-inflammatory Protective Effect of Lingguizhugan Decoction via NF-κB Pathway on LPS-induced Acute Lung Injury in Mice. Chin. J. Exp. Tradit. Med. Formulae 2023, 29, 14–21. [Google Scholar]
- Yue, S.; Hu, B.; Wang, Z.; Yue, Z.; Wang, F.; Zhao, Y.; Yang, Z.; Shen, M. Salvia miltiorrhiza compounds protect the liver from acute injury by regulation of p38 and NFκB signaling in Kupffer cells. Pharm. Biol. 2014, 52, 1278–1285. [Google Scholar] [CrossRef] [PubMed]
- Saito, C.; Zwingmann, C.; Jaeschke, H. Novel mechanisms of protection against acetaminophen hepatotoxicity in mice by glutathione and N-acetylcysteine. Hepatology 2010, 51, 246–254. [Google Scholar] [CrossRef]
- Wang, Z.; Hu, J.N.; Yan, M.H.; Xing, J.J.; Liu, W.C.; Li, W. Caspase-Mediated Anti-Apoptotic Effect of Ginsenoside Rg5, a Main Rare Ginsenoside, on Acetaminophen-Induced Hepatotoxicity in Mice. J. Agric. Food Chem. 2017, 65, 9226–9236. [Google Scholar] [CrossRef]
- Che, Y. Berberine Decreases Apoptosis by Regulating Bcl-2 Family Proteins and Activating Caspase Pathway in Hypoxia/Reoxygenation-Induced H9C2 Cardiomyocytes; Fourth Military Medical University: Xi’an, China, 2014. [Google Scholar]
- Yu, H.; Guo, P.; Xie, X.; Wang, Y.; Chen, G. Ferroptosis, a new form of cell death, and its relationships with tumourous diseases. J. Cell. Mol. Med. 2017, 21, 648–657. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, P.; Zhai, B.; Zhang, M.; Xiang, Y.; Fang, J.; Xu, S.; Gao, Y.; Chen, X.; Sui, X.; et al. The emerging role of ferroptosis in inflammation. Biomed. Pharmacother. 2020, 127, 110108. [Google Scholar] [CrossRef]
- Lei, G.; Zhang, Y.; Koppula, P.; Liu, X.; Zhang, J.; Lin, S.H.; Ajani, J.A.; Xiao, Q.; Liao, Z.; Wang, H.; et al. The role of ferroptosis in ionizing radiation-induced cell death and tumor suppression. Cell Res. 2020, 30, 146–162. [Google Scholar] [CrossRef] [PubMed]
- Dong, T. Study on the Mechanism of Nrf2 Signaling Pathway Alleviating PM2.5 Induced Lung Injury by Regulating Ferroptosis; Jilin University: Changchun, China, 2022. [Google Scholar]
- Ning, J. Important Roles of Nrf2 in Protection of Transient Controlled Hypothermia for Lung Injury after Limb’s Explosion Injury in Rats; Third Military Medical University: Chongqing, China, 2008. [Google Scholar]
- Min, L. Nrf2, HO-1 in Cerebral Ischemia-Reperfusion in Rats and the Neuroprotective Effect of Oxymatrine; Hebei Medical University: Shijiazhuang, China, 2009. [Google Scholar]
- Sun, X.; Ou, Z.; Chen, R.; Niu, X.; Chen, D.; Kang, R.; Tang, D. Activation of the p62-Keap1-NRF2 pathway protects against ferroptosis in hepatocellular carcinoma cells. Hepatol. Baltim. Md. 2016, 63, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y. Erastin Induced Ferroptosis of Colon Cancer Cells through Regulation of SPINK4/VIL1 and NF-κB Pathway; Guilin Medical University: Guilin, China, 2020. [Google Scholar]
- Li, X.; Teitgen, A.M.; Shirani, A.; Ling, J.; Busta, L.; Cahoon, R.E.; Zhang, W.; Li, Z.; Chapman, K.D.; Berman, D.; et al. Discontinuous fatty acid elongation yields hydroxylated seed oil with improved function. Nat. Plants 2018, 4, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Haro-González, J.N.; Castillo-Herrera, G.A.; Martínez-Velázquez, M.; Espinosa-Andrews, H. Clove Essential Oil (Syzygium aromaticum L. Myrtaceae): Extraction, Chemical Composition, Food Applications, and Essential Bioactivity for Human Health. Molecules 2021, 26, 6387. [Google Scholar] [CrossRef]
- Jing, F.; Li, F.; Zhang, T.; Lv, C.; Zhao, T. Research Progress of Chemical Constituents and Pharmacological Activities of Forsythia. J. Chin. Med. Mater. 2023, 46, 242–251. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | Sample | VC * | VB1 * | VB2 * | VE * | VK # | VB6 # | β-Carotene # |
|---|---|---|---|---|---|---|---|---|
| Seedling | O. violaceus | 132.38 | 0.06 | 0.13 | 0.08 | 0.06 | 1.14 | 1.09 |
| B.chinensis | 29.70 | 0.04 | 0.11 | 0.05 | 0.08 | 0.78 | 0.57 | |
| Moss | O. violaceus | 101.62 | 0.05 | 0.11 | 0.11 | 0.06 | 0.84 | 1.31 |
| B. chinensis | 44.57 | 0.04 | 0.09 | 0.10 | 0.07 | 0.67 | 1.01 |
| Sample | β-Carotene @ | VB1 * | VB2 * | VC * |
|---|---|---|---|---|
| O. violaceus | 1620 | 0.10 | 0.05 | 27 |
| Brassica oleracea var. albiflora Kuntze | 3450 | 0.20 | 0.00 | 78 |
| Brassica campestris var. purpuraria | 1255 | 0.10 | 0.06 | 21 |
| Brassica caulorapa Pasq. | 20 | 0.04 | 0.82 | 41 |
| Baby Bok Choy | 1680 | 0.02 | 0.00 | 26 |
| Brassica oleracea var. botrytis Linnaeus | 20 | 0.08 | 0.08 | 81 |
| Brassica rapa var. glabra Regel | 40 | 0.08 | 0.04 | 28 |
| Brassica juncea (L.) Czern. et Coss. Var. foliosa L. H. Bailey | 1700 | 0.02 | 0.10 | 72 |
| Brassicajuncea (L.) Czern. et Coss. Var. multiceps Tsen et Lee | 310 | 0.03 | 0.11 | 31 |
| Amino Acid | Content | |
|---|---|---|
| Seedling | Moss | |
| * Ile | 42.30 | 43.99 |
| * Leu | 79.27 | 84.03 |
| * Phe | 38.66 | 40.21 |
| * Lys | 58.49 | 61.99 |
| * Thr | 41.92 | 44.85 |
| * Val | 63.16 | 63.79 |
| * Met | 6.66 | 6.56 |
| * Trp | 13.54 | 12.32 |
| Ser | 42.18 | 43.02 |
| Tyr | 27.51 | 28.89 |
| Ala | 60.62 | 62.44 |
| Glu | 112.33 | 117.95 |
| His | 20.09 | 20.09 |
| Arg | 41.71 | 44.21 |
| Pro | 114.88 | 92.05 |
| Cys | 6.74 | 6.94 |
| Asp | 78.42 | 83.13 |
| Gly | 51.29 | 52.83 |
| Elements | Flower | Root | Leaf | |
|---|---|---|---|---|
| Macroelement | B | 19.69 | 12.01 | 22.05 |
| Na | 210.53 | 199.25 | 240.61 | |
| Mg | 1204.99 | 380.79 | 1271.30 | |
| Si | 322.58 | 239.72 | 207.08 | |
| P | 3864.03 | 1747.57 | 2613.93 | |
| K | 12,911.21 | 8075.82 | 10,043.72 | |
| Ca | 4750.79 | 3121.87 | 12,834.32 | |
| Secondary element | Mn | 32.64 | 16.31 | 86.86 |
| Fe | 234.52 | 123.06 | 1776.91 | |
| Cu | 8.56 | 3.34 | 4.98 | |
| Zn | 44.10 | 16.65 | 28.75 | |
| Br | 2.69 | 2.02 | 7.86 | |
| Sr | 23.13 | 35.13 | 88.83 | |
| I | 0.95 | 1.90 | 1.30 | |
| Heavy metal element | Al | 170.03 | 131.51 | 95.07 |
| Ti | 15.38 | 12.30 | 43.33 | |
| Cr | 2.14 | 2.22 | 221.68 | |
| Ni | 1.66 | 0.90 | 18.70 | |
| Pb | 1.357 | 0.184 | 0.679 | |
| Trace beneficial element | Li | 0.301 | 0.198 | 0.414 |
| Co | 0.374 | 0.174 | 0.578 | |
| Mo | 0.885 | 0.576 | 1.476 | |
| Rare earth element | La | 0.248 | 0.135 | 0.114 |
| Ce | 0.717 | 0.420 | 0.343 | |
| Nd | 0.185 | 0.102 | 0.087 | |
| Sc | 0..233 | 0.158 | 0.179 | |
| Compounds | Molecular Formula | Relative Area Percentage (%) | |
|---|---|---|---|
| MIBSE | SE | ||
| Palmitic acid | C16H32O2(C16:0) | 8.84 | 9.11 |
| Linoleic acid | C18H32O2(C18:2) | 55.62 | 54.93 |
| Oleic acid | C18H34O2(C18:1) | 15.85 | 15.58 |
| Stearic acid | C18H36O2(C18:0) | 3.67 | 4.19 |
| Linolenic acid | C18H30O2(C18:3) | 4.02 | 3.78 |
| cis-11,14-Eicosenoic acid | C20H36O2(C20:2) | 0.51 | 0.56 |
| 11-Eicosenoic acid | C20H38O2(C20:1) | 3.87 | 4.11 |
| Arachidic acid | C20H40O2(C20:0) | 2.94 | 3.11 |
| Behenic acid | C22H44O2(C22:0) | 1.18 | 1.31 |
| Nervonic acid | C24H46O2(C24:1) | 2.40 | 2.32 |
| Tetracosenoic acid | C24H48O2(C24:0) | 1.10 | 1.01 |
| Plant | Oil Content (%) | Crude Protein Content (%) | Glucosinolate Content (μg/g) |
|---|---|---|---|
| O. violaceus | 50.29 | 52.32 | 35.24 |
| Ning-You-7-Hao | 41.71 | 34.20 | 69.67 |
| Westar | 42.90 | 37.58 | 6.77 |
| No. | Compound | Type | Formula | Mol. wt. | References |
|---|---|---|---|---|---|
| 1 | Quercetin | flavonols | C15H10O7 | 302.02 | [37] |
| 2 | Kaempferol | flavonols | C15H10O6 | 286.05 | [37] |
| 3 | Isorhamnetin | flavonols | C16H12O7 | 316.06 | [37] |
| 4 | Isorhamnetin-3-O-β-d-glucopyranoside | flavonols | C22H22O12 | 478.11 | [41] |
| 5 | 3,4′,6,8-tetyahydroxy-flavone-7-C-glucoside | flavonols | C21H20O11 | 448.10 | [42] |
| 6 | kaeMpferol 3-O-β-d-galactosyl-7-O-α-L-rhamnoside | flavonols | C27H30O15 | 594.16 | [42] |
| 7 | kaeMpferol 3-O-β-d-arabinosyl-7-O-α-L-rhamnoside | flavonols | C26H28O14 | 564.15 | [42] |
| 8 | kaeMpferol 3-O-β-d-glucopyranosyl-7-O-α-L-rhamnoside | flavonols | C27H30O15 | 594.16 | [42] |
| 9 | Quercetin 3-O-β-d-glucopyranosyl-7-O-α-L-rhamnoside | flavonols | C27H30O16 | 610.15 | [42] |
| 10 | Kaempferol-7-O-α-L-rhamnoside | flavonols | C21H20O10 | 432.11 | [42] |
| 11 | kaempferol-3-O-β-d-glucoside | flavonols | C21H20O11 | 448.10 | [42] |
| 12 | Quercetin-7-O-α-L-rhamnoside | flavonols | C21H20O11 | 448.10 | [42] |
| 13 | 5,7,3′,5′-tetyahydroxy-flavone-7-O-β-d-glucoside | flavanones | C21H22O11 | 450.12 | [41] |
| 14 | aromadendrin-7-O-β-d-glucopyranoside | flavanones | C21H22O10 | 434.12 | [41] |
| 15 | hesperetin-7-O-β-d-glucopyranoside | flavanones | C22H24O11 | 464.12 | [41] |
| 16 | 3′,5′,5,7-tetyahydroxy-dihydroflavone | flavanones | C15H12O6 | 288.06 | [9] |
| 17 | Geristein | isoflavones | C15H10O5 | 270.05 | [11] |
| 18 | acactin-7-O-β-d-glucoside | flavones | C22H22O10 | 446.12 | [11] |
| 19 | Genkwanin | flavones | C16H12O5 | 284.07 | [41] |
| 20 | acacetin-7-O-neohesperidoside | flavones | C28H32O14 | 592.18 | [41] |
| 21 | apigenin-7-O-β-d-glucoside | flavones | C21H20O10 | 432.11 | [41] |
| 22 | chrysoeriol-7-O-β-d-glucoside | flavones | C22H22O11 | 462.12 | [41] |
| 23 | Catechin | flavan-3-ols | C15H12O6 | 288.06 | [43] |
| No. | Compound | Formula | Mol. wt. | References |
|---|---|---|---|---|
| 24 | orychophragmuspine A | C26H33N3O5 | 467.27 | [12] |
| 25 | orychophragmuspine B | C26H33N3O5 | 467.24 | [12] |
| 26 | orychophragmuspine C | C26H33N3O5 | 467.24 | [12] |
| 27 | orychophragmuspine D | C28H35N3O6 | 509.25 | [12] |
| 28 | orychophragmuspine E | C28H35N3O6 | 509.25 | [12] |
| 29 | orychophragmuspine F | C27H35N3O5 | 481.26 | [12] |
| 30 | orychophragmuspine G | C27H35N3O5 | 481.26 | [12] |
| 31 | orychophragmuspine H | C25H33N3O4 | 439.25 | [12] |
| 32 | orychophragmuspine I | C26H35N3O5 | 469.26 | [46] |
| 33 | orychophragine A | C14H17N3O4 | 291.12 | [47] |
| 34 | orychophragine B | C16H19N5O5 | 361.14 | [47] |
| 35 | orychophragine C | C16H19N5O5 | 361.14 | [47] |
| 36 | orychophragine D | C16H18N6O4 | 358.14 | [48] |
| 37 | (+)-orychoviolines A | C25H33O4N3 | 439.25 | [50] |
| 38 | (−)-orychoviolines A | C25H33O4N3 | 439.25 | [50] |
| 39 | (+)-orychoviolines B | C27H39O5N3 | 485.29 | [50] |
| 40 | (−)-orychoviolines B | C27H39O5N3 | 485.29 | [50] |
| 41 | demethylorychophragine A | C13H15N3O4 | 277.11 | [51] |
| 42 | 4-hydroxy-5-methoxy-3-quinolinecarbonitrile | C11H8N2O2 | 200.06 | [42] |
| 43 | Cappariloside A | C16H18N2O6 | 334.12 | [42] |
| 44 | 2-Phenylacetamide | C8H9NO | 135.07 | [42] |
| 45 | Uracil | C4H4N2O2 | 112.03 | [42] |
| 46 | Uridine | C9H12N2O6 | 244.07 | [42] |
| 47 | isolumichrome | C12H10N4O2 | 242.08 | [42] |
| 48 | 1-ribosyl-2-3-diketo-1,2,3,4-tetrahydro-6,7-dimethyl-quinoxaline | C15H20N2O6 | 324.13 | [42] |
| 49 | 3,6-dibenzyl-2,5-dioxopiperazine | C16H14N2O2 | 266.11 | [42] |
| 50 | perlolyrine | C16H12N2O2 | 264.09 | [42] |
| 51 | DL-isoleucyl-leucyl anhydride | C12H22N2O2 | 226.17 | [42] |
| 52 | 4-oxo-1,4-dihydroquinoline-3-carbonitrile | C10H6N2O | 170.05 | [42] |
| 53 | epigoitrin | C5H7NOS | 129.02 | [50] |
| 54 | Adenosine | C10H13N5O4 | 267.10 | [9] |
| 55 | 2′-Dideoxyadenosine | C10H13N5O3 | 251.10 | [9] |
| 56 | 7,8-Dimethylalloxazine | C12H10N4O2 | 242.08 | [52] |
| 57 | N-2-hydroxy-3-butenyl-Benzamide | C11H13NO2 | 191.09 | [53] |
| 58 | 2-Amino-5-hydroxybenzoic acid | C7H7NO3 | 153.04 | [53] |
| 59 | N-benzoyl-epigoitri | C12H11NO2S | 233.05 | [53] |
| 60 | α-[(2-carboxyacetyl)amino]-Benzeneacetic acid | C11H11NO5 | 237.06 | [53] |
| 61 | 3-phenyl-6-vinylmorpholin-2-one | C12H13NO2 | 203.09 | [53] |
| No. | Compound | Type | Formula | Mol. wt. | References |
|---|---|---|---|---|---|
| 62 | orychophramarin A | coumarin | C15H10O5 | 270.05 | [13] |
| 63 | orychophramarin B | coumarin | C15H10O5 | 270.05 | [13] |
| 64 | orychophramarin C | coumarin | C16H12O6 | 300.06 | [13] |
| 65 | orychophramarin D | coumarin | C21H22O10 | 434.12 | [13] |
| 66 | 7-hydroxy-6-methoxy coumarin | coumarin | C10H8O4 | 192.04 | [13] |
| 67 | lariciresinol | lignans | C20H24O6 | 360.16 | [13] |
| 68 | 11-deoxykompasinol A | lignans | C25H24O7 | 436.15 | [31] |
| 69 | (+)-lariciresinol-9-O-β-d-glucopyranoside | lignans | C26H34O11 | 522 | [31] |
| 70 | (+)-isolariciresinol-9′-O-β-d-glucopyranoside | lignans | C26H34O11 | 522 | [31] |
| 71 | (+)-isolariciresinol-9-O-β-d-glucopyranoside | lignans | C26H34O11 | 522 | [31] |
| 72 | (−)-isolariciresinol-9′-O-β-d-glucopyranoside | lignans | C26H34O11 | 522 | [31] |
| 73 | (−)-5′-methoxyisolariciresinol-9′-O-β-d-glucopyranoside | lignans | C27H36O11 | 434.16 | [31] |
| 74 | p-hudroxycoumaric | phenylpropanoic acids | C9H8O3 | 164.05 | [42] |
| 75 | caffeic acid | phenylpropanoic acids | C9H8O4 | 180.04 | [14] |
| 76 | trans-Ferulic acid | phenylpropanoic acids | C10H10O4 | 194.06 | [42] |
| 77 | 3,4-dimethoxy cinnamic acid | phenylpropanoic acids | C11H12O4 | 208.07 | [42] |
| No. | Compound | Type | Formula | Mol. wt. | References |
|---|---|---|---|---|---|
| 78 | salicylic acid | benzoic acid | C7H6O3 | 138.03 | [14] |
| 79 | vanillic acid | benzoic acid | C8H8O4 | 168.04 | [14] |
| 80 | protocatechuic acid | benzoic acid | C7H6O4 | 154.03 | [14] |
| 81 | syringate | benzoic acid | C9H10O5 | 198.05 | [14] |
| 82 | benzonic acid | benzoic acid | C7H6O2 | 122.04 | [41] |
| 83 | 2-hydroxybenzoic acid | benzoic acid | C7H6O3 | 138.03 | [41] |
| 84 | 4-hydroxybenzoic acid | benzoic acid | C7H6O3 | 138.03 | [41] |
| 85 | 4-hydroxy-3-methoxy-benzoic acid | benzoic acid | C8H8O4 | 168.04 | [41] |
| 86 | Eudesmic acid | benzoic acid | C10H12O5 | 212.07 | [41] |
| 87 | Isovanillic acid | benzoic acid | C8H8O4 | 168.04 | [42] |
| 88 | 2,3-dihydroxybenzoic acid | benzoic acid | C7H6O4 | 154.03 | [42] |
| No. | Compound | Type | Formula | Mol. wt. | References |
|---|---|---|---|---|---|
| 89 | orychovioside A | monoterpene glycoside | C22H38O10 | 462.25 | [46] |
| 90 | orychoside A | triterpene saponin | C54H88O22 | 1088.58 | [15] |
| 91 | orychoside B | Diterpene glycosides | C26H40O10 | 512.26 | [15] |
| 92 | rubusoside | Diterpene glycosides | C32H50O13 | 642.33 | [15] |
| 93 | mimengoside C | triterpene saponin | C54H88O22 | 1088.58 | [15] |
| 94 | mimengoside D | triterpene saponin | C47H76O17 | 912.51 | [15] |
| 95 | mimengoside E | triterpene saponin | C54H88O22 | 1088.58 | [15] |
| 96 | mimengoside F | triterpene saponin | C47H76O17 | 912.51 | [15] |
| 97 | buddlejasaponin I | triterpene saponin | C54H88O22 | 1088.58 | [15] |
| 98 | buddlejasaponin III | triterpene saponin | C47H76O17 | 912.51 | [15] |
| 99 | buddlejasaponin Ia | triterpene saponin | C55H92O23 | 1120.60 | [15] |
| 100 | buddlejasaponin IIIa | triterpene saponin | C48H80O18 | 944.53 | [15] |
| 101 | 3-O-α-L-rhamnose-(1→4)-β-d-glucose(1→3)-[β-d-glucose-(1→2)]-β-d-fucose-23,28-dihydroxysaikosaponin-11,13(18)-diene | triterpene saponin | C54H88O21 | 1072.58 | [15] |
| 102 | 3β-O-(β-d-glucose-(1→4)-β-d-glucose(1→3)-[β-d-glucose-(1→2)]-β-d-fucose-23,28-dihydroxysaikosaponin-11,13(18)-diene | triterpene saponin | C53H86O21 | 1058.57 | [15] |
| No. | Compound | Formula | Mol. wt. | References |
|---|---|---|---|---|
| 103 | dioscin | C45H72O16 | 868.48 | [11] |
| 104 | gracillin | C45H72O17 | 884.48 | [11] |
| 105 | β-Sitosterol | C29H50O | 414.39 | [41] |
| 106 | Daucosterol | C35H60O6 | 576.44 | [41] |
| 107 | β-Sitosterol-3-O-β-d-glucopyranoside-20-O-palmitate | C51H90O7 | 814.67 | [41] |
| No. | Compound | Formula | Mol. wt. | References |
|---|---|---|---|---|
| 108 | Centaureidin 3-rutinoside | C28H33O14+ | 593.19 | [67] |
| 109 | Malvidin 3,5-diglucoside | C29H35O17+ | 655.19 | [67] |
| 110 | Centaureidin 3-coumaroylglucoside-5-glucoside | C38H41O20+ | 817.22 | [67] |
| 111 | Centaureidin 3-p-coumaroyl diglucoside-5-glucoside | C43H49O25+ | 965.26 | [67] |
| 112 | Centaureidin 3-feruloylsophoroside-5-malonyl glucoside | C47H53O26+ | 1033.28 | [67] |
| 113 | Centaureidin 3-coumaroyl locust glycoside-5-malonyl glucoside | C48H55O27+ | 1063.29 | [67] |
| Compounds | IC50 (μM) | References | ||||
|---|---|---|---|---|---|---|
| Hela | HepG2 | A549 | HCT-116 | SH-SY5Y | ||
| 11-deoxykompasinol A | 9.43 ± 0.62 | 18.26 ± 1.31 | / | / | / | [31] |
| (+)-lariciresinol-9-O-β-d-glucopyranoside | 17.82 ± 1.17 | 36.78 ± 1.21 | / | / | / | [31] |
| β-d-glucopyranosyl-12-hydroxyjasmonate | 22.20 ± 2.24 | 29.10 ± 2.45 | / | / | / | [31] |
| orychophragine A | 11.91 ± 0.46 | 7.73 ± 0.55 | 10.79 ± 0.86 | 9.93 ± 0.71 | / | [47] |
| orychoside A | / | 7.13 ± 0.10 | 8.77 ± 0.31 | / | / | [41] |
| mimengoside C | / | 7.82 ± 0.17 | 6.78 ± 0.21 | / | / | [41] |
| mimengoside D | / | 9.62 ± 0.68 | 11.63 ± 0.50 | / | / | [41] |
| mimengoside E | / | 16.82 ± 1.46 | 14.03 ± 0.49 | / | / | [41] |
| mimengoside F | / | 22.20 ± 1.24 | 19.10 ± 0.45 | / | / | [41] |
| buddlejasaponin I | / | 0.78 ± 0.21 | 0.48 ± 0.04 | / | / | [41] |
| buddlejasaponin III | / | 1.06 ± 0.09 | 0.96 ± 0.12 | / | / | [41] |
| orychophramarin A | 8.91 ± 0.65 | / | / | 5.10 ± 0.46 | / | [13] |
| orychophramarin B | 23.47 ± 1.37 | / | / | 28.30 ± 0.91 | / | [13] |
| orychophramarin C | 14.24 ± 0.67 | / | / | 10.12 ± 0.54 | / | [13] |
| orychophramarin D | >50 | / | / | 44.71 ± 1.93 | / | [13] |
| perlolyrine | 51.3 ± 1.32 | / | / | / | 23.4 ± 2.88 | [42] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Zhang, G.; Cui, W.; Ge, C.; Li, B.; Li, M.; Liu, S.; Wang, L. Recent Advances in the Nutritional Value, Chemical Compositions, Pharmacological Activity, and Application Value of Orychophragmus violaceus: A Comprehensive Review. Molecules 2024, 29, 1314. https://doi.org/10.3390/molecules29061314
Chen X, Zhang G, Cui W, Ge C, Li B, Li M, Liu S, Wang L. Recent Advances in the Nutritional Value, Chemical Compositions, Pharmacological Activity, and Application Value of Orychophragmus violaceus: A Comprehensive Review. Molecules. 2024; 29(6):1314. https://doi.org/10.3390/molecules29061314
Chicago/Turabian StyleChen, Xiaolan, Guangjie Zhang, Wenjin Cui, Chunbo Ge, Bin Li, Min Li, Shuchen Liu, and Lin Wang. 2024. "Recent Advances in the Nutritional Value, Chemical Compositions, Pharmacological Activity, and Application Value of Orychophragmus violaceus: A Comprehensive Review" Molecules 29, no. 6: 1314. https://doi.org/10.3390/molecules29061314
APA StyleChen, X., Zhang, G., Cui, W., Ge, C., Li, B., Li, M., Liu, S., & Wang, L. (2024). Recent Advances in the Nutritional Value, Chemical Compositions, Pharmacological Activity, and Application Value of Orychophragmus violaceus: A Comprehensive Review. Molecules, 29(6), 1314. https://doi.org/10.3390/molecules29061314