cDNA Cloning, Overexpression, Purification and Pharmacologic Evaluation for Anticancer Activity of Ribosomal Protein L23A Gene (RPL23A) from the Giant Panda

Abstract
:1. Introduction
2. Results and Discussion
2.1. Analysis of the cDNA of RPL23A from the Giant Panda
2.2. Prediction and Analysis of Primary Structure, Protein Functional Sites and Advanced Structure in RPL23A Protein of the Giant Panda
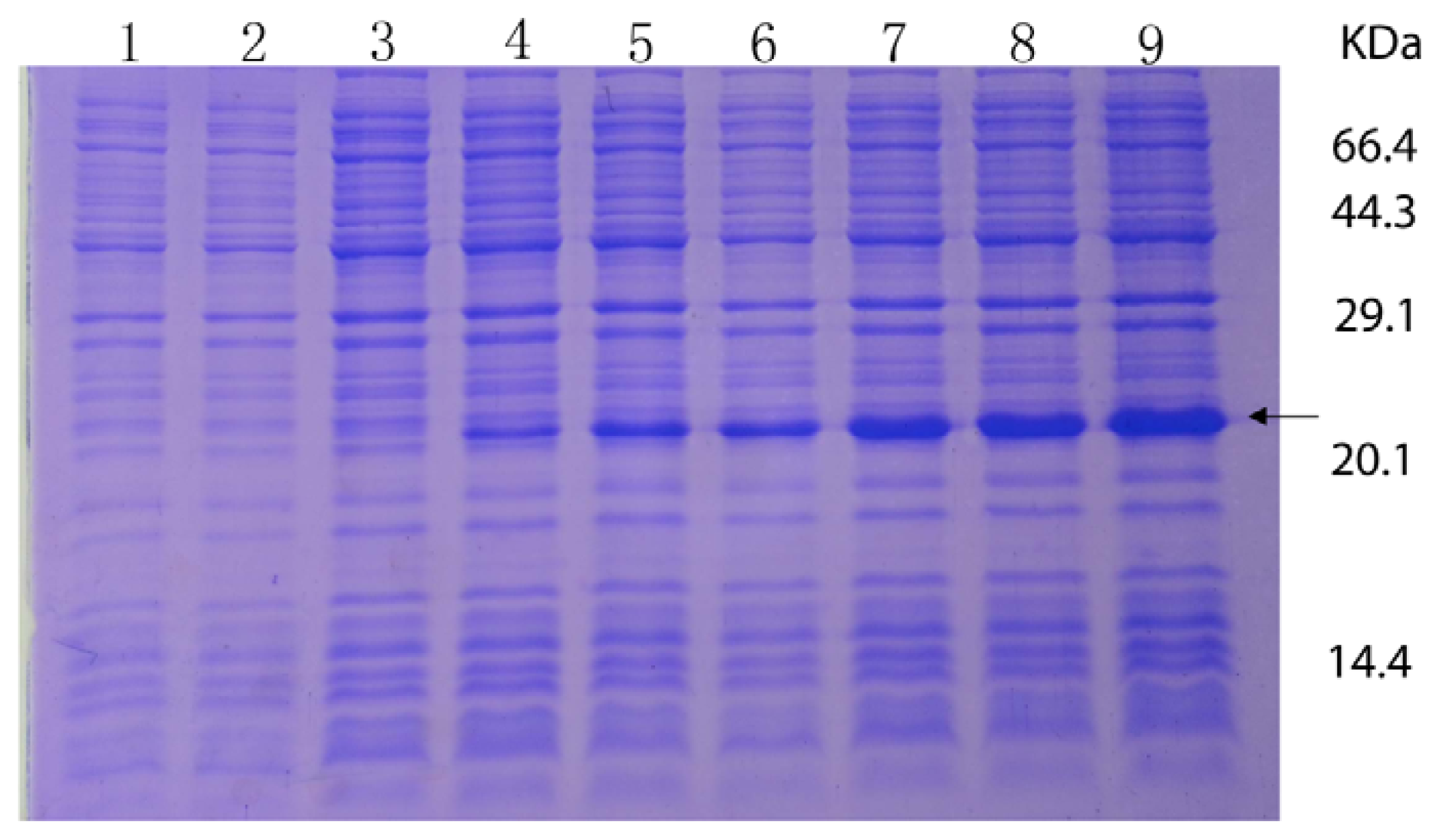
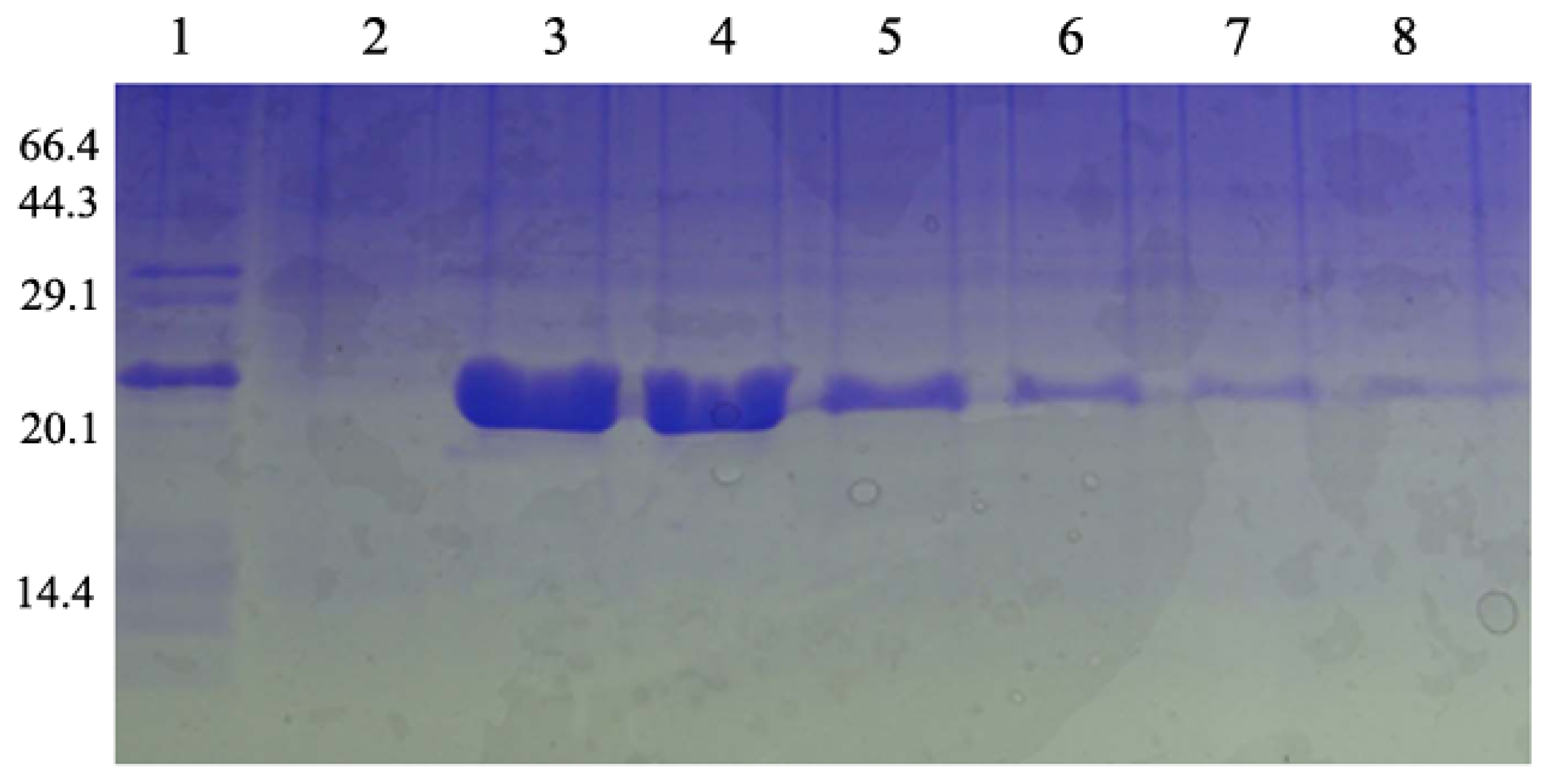
2.3. Overexpression and Purification of the RPL23A Gene in E. coli
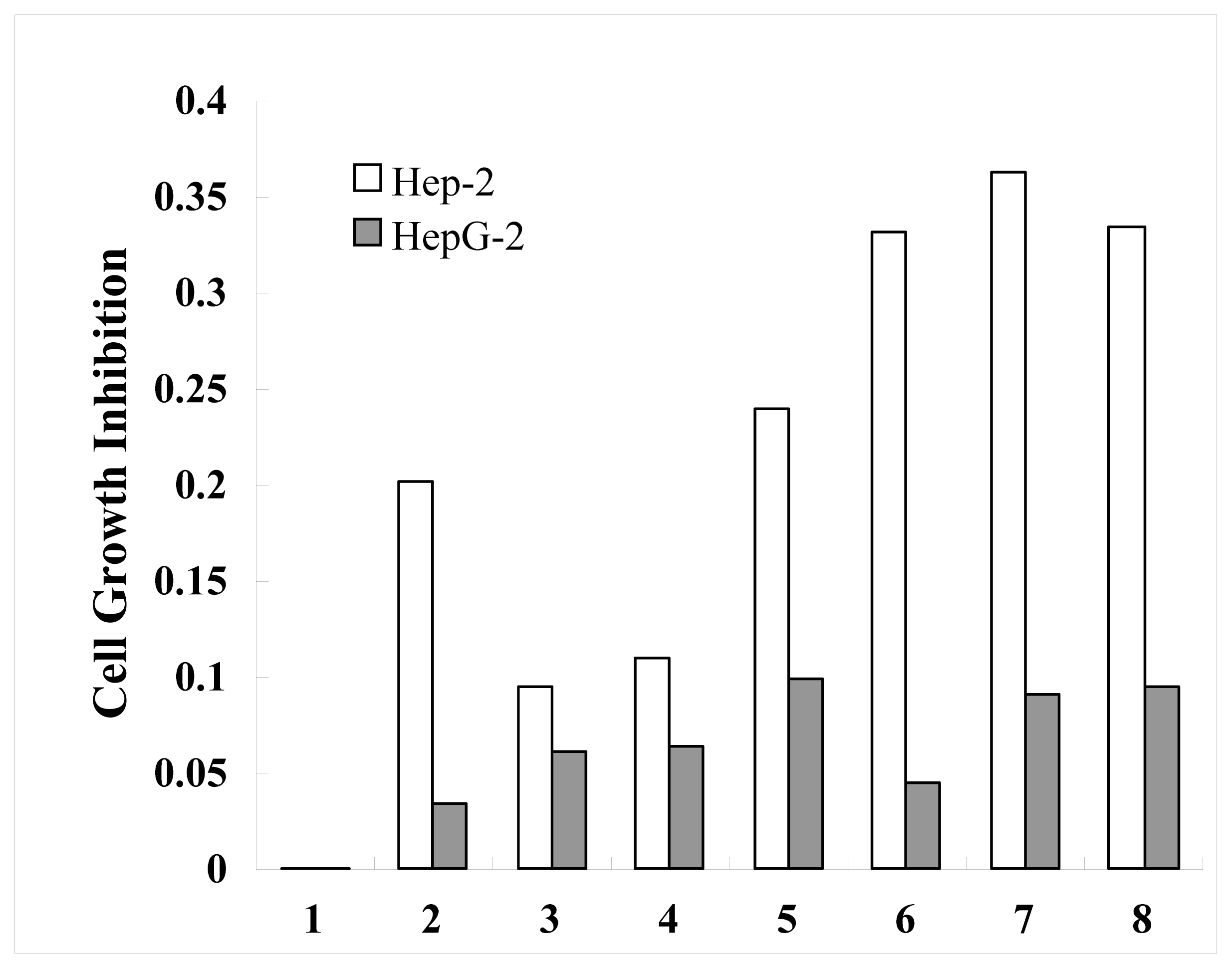
2.4. Anticancer Activity of Ribosomal Protein L23A (RPL23A) from the Giant Panda on Human Laryngeal Carcinoma Hep-2 cells and Human Hepatoma HepG2 Cells
2.5. The effect of Ribosomal Protein L23A (RPL23A) from the Giant Panda on Morphology of Human Laryngeal Carcinoma Hep-2 cells
3. Experimental Section
3.1. Materials
3.2. RNA Isolation
3.3. Primers Design, RT-PCR
Pd-RPL23A-F: 5′-CCTTTTCGACAAGATGGCG-3′;Pd-RPL23A-R: 5′-GAATTAGCCAGCTGGACTCA-3′.
3.4. Cloning and Identifying the cDNA Sequence
3.5. Construction of the Expression Vector and Overexpression of Recombinant RPL23A
3.6. Purification of Recombinant Protein RPL23A
3.7. Cell Lines Culture
3.8. Test the Cell Viability of Purified Recombinant Protein RPL23A on Human Laryngeal Carcinoma Hep-2 Cells and Human Hepatoma HepG-2 Cell Activity by MTT Assay
3.9. Data Analysis
4. Conclusions
Acknowledgements
References
- Wool, I.G.; Chan, Y.L.; Glück, A. Structure and evolution of mammalian ribosomal proteins. Biochem. Cell Biol 1996, 73, 933–947. [Google Scholar]
- Wool, I.G. Extraribosomal functions of ribosomal proteins. Trends Biochem. Sci 1996, 21, 164–165. [Google Scholar]
- Fan, W.; Christensen, M.; Eichler, E.; Zhang, X.; Lennon, G. Cloning, sequencing, gene organization, and localization of the human ribosomal protein RPL23A gene. Genomics 1998, 46, 234–239. [Google Scholar]
- Yang, F.; Liu, W.P. The progress of ribosomal protein genes and human diseases. J. Clin. Exp. Pathol 2005, 20, 354–356. [Google Scholar]
- Jiang, H.; Lin, J.J.; Tao, J.; Fisher, P.B. Suppression of human ribosomal protein L23A expression during cell growth inhibition by interferon-beta. Oncogene 1997, 14, 473–480. [Google Scholar]
- Rogan, S.; Heaphy, S. The vaccinia virus E3L protein interacts with SUMO-1 and ribosomal protein L23a in a yeast two hybrid assay. Virus Genes 2001, 21, 193–195. [Google Scholar]
- Jäkel, S.; Görlich, D. Importinβ, transportin, RanBP5 and RanBP7 mediate nuclear import of ribosomal proteins in mammalian cells. EMBO J 1998, 17, 4491–4502. [Google Scholar]
- Anand, P.; Kunnumakkara, A.B.; Sundaram, C.; Harikumar, K.B.; Tharakan, S.T.; Lai, O.S.; Sung, B.; Aggarwal, B.B. Cancer is a preventable disease that requires major lifestyle changes. Pharm. Res 2008, 25, 2097–2116. [Google Scholar]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin 2011, 61, 69–90. [Google Scholar]
- Croce, C.M. Oncogenes and cancer. N. Engl. J. Med 2008, 358, 502–511. [Google Scholar]
- Tolar, J.; Neglia, J.P. Transplacental and other routes of cancer transmission between individuals. J. Pediatr. Hematol. Oncol 2003, 25, 430–434. [Google Scholar]
- Uechi, T.; Tanaka, T.; Kenmochi, N. A complete map of the human ribosomal protein genes: Assignment of 80 genes to the cytogenetic map and implications for human disorders. Genomics 2001, 72, 223–230. [Google Scholar]
- Wu, G.F.; Hou, Y.L.; Hou, W.R.; Song, Y.; Zhang, T. Giant pandaribosomal protein s14: cDNA, genomic sequence cloning, sequence analysis, and overexpression. Genet. J. Mol. Res 2010, 9, 2004–2015. [Google Scholar]
- Du, Y.J.; Luo, X.Y.; Hao, Y.Z.; Zhang, T.; Hou, W.R. Cloning and overexpression of acidic ribosomal phosphoprotein P1 gene (RPLP1) from the Giant Panda. Inter. J. Bio. Sci 2007, 3, 428–433. [Google Scholar]
- Hou, W.R.; Chen, Y.; Peng, Z.S.; Wu, X.; Tang, Z.X. cDNA cloning and sequences analysis of ubiquinol-cytochrome c reductase complex ubiquinone-binding protein (QP-C) from giant panda. Acta Theriol. Sin 2007, 27, 190–194. [Google Scholar]
- Hou, Y.L.; Du, Y.J.; Hou, W.R.; Zhou, C.Q.; Hao, Y.Z.; Zhang, T. Cloning and sequence analysis of translocase of inner mitochondrial membrane 10 homolog (yeast) gene (TIMM10) from the Giant Panda. J. Cell. Anim. Biol 2009, 3, 9–14. [Google Scholar]
- Liao, M.J.; Zhu, M.Y.; Zhang, Z.H.; Zhang, A.J.; Li, G.H.; Sheng, F.J. Cloning and sequence analysis of FSH and LH in the Giant Panda (Ailuropoda melanoleuca). Anim. Reprod. Sci 2003, 77, 107–116. [Google Scholar]
- Wu, Z.A.; Liu, W.X.; Murphy, C.; Gall, J. Satellite DNA sequence from g enomic DNA of the giant panda. Nucleic Acids Res 1990, 18. [Google Scholar] [CrossRef]
- Zhang, T.; Hou, W.R.; Hou, Y.L. genomic sequence cloning and overexpression of ribosomal protein s20 gene (RPS20) from the Giant Panda (Ailuropoda melanoleuca). Afr. J. Biotechnol 2009, 8, 5627–5632. [Google Scholar]
- Su, X.L.; Hou, Y.L.; Yuan, S.B.; Tian, M.J.; Sun, B.; Li, J.; Wu, G.F.; Song, Y.; Hou, W.R. cDNA, Genomic sequence cloning and sequence analysis of ribosomal protein L18A gene(RPL18A) from the Giant Panda (Ailuropoda melanoleuca). Proceedings of the 2010 3rd International Conference on Biomedical Engineering and Informatics (BMEI), Yantai, China, 16–18 October 2010; pp. 2165–2169.
- Frederik, N.; Wuppermann, K.M.; Marion, J.; Christian, A.J.; Johannes, H.H. Chlamydia pneumoniae GroEL1 protein is cell surface associated and required for infection of HEp-2 cells. J. Bacteriol 2008, 190, 3757–3767. [Google Scholar]
- Yu, L.; Li, H.Z.; Lu, S.M.; Liu, W.W.; Li, J.F.; Wang, H.B.; Xu, W. Alteration in TWIST expression: Possible role in paclitaxel-induced apoptosis in human laryngeal carcinoma hep-2 cell line. Croat. Med. J 2009, 50, 536–542. [Google Scholar]
- Chen, C.C.; Chan, W.H. Inhibition of citrinin-induced apoptotic biochemical signaling in human hepatoma G2 cells by resveratrol. Int. J. Mol. Sci 2009, 10, 3338–3357. [Google Scholar]
- Wool, I.G. The structure and function of eukaryotic ribosomes. Annu. Rev. Biochem 1979, 48, 719–754. [Google Scholar]
- Nutting, C.M.; Robinson, M.; Birchall, M. Survival from laryngeal cancer in England and Wales up to 2001. Br. J. Cancer 2008, 99, 38–39. [Google Scholar]
- Barnard, G.F.; Staniunas, R.J.; Bao, S.; Mafune, K.; Steele, G.D., Jr; Gollan, J.L.; Chen, L.B. Increased expression of human ribosomal phosphoprotein P0 messenger RNA in hepatocellular carcinoma and colon carcinoma. Cancer Res. 1992, 52, 3067–3072. [Google Scholar]
- Chiao, P.J.; Shin, D.M.; Sacks, P.G.; Hong, W.K.; Tainsky, M.A. Elevated expression of the ribosomal protein S2 gene in human tumors. Mol. Carcinog 1992, 5, 219–231. [Google Scholar]
- Denis, M.G.; Chadeneau, C.; Lecabellec, M.T.; LeMoullac, B.; LeMevel, B.; Meflah, K.; Lustenberger, P. Over-expression of the S13 ribosomal protein in actively growing cells. Int. J. Cancer 1993, 55, 275–280. [Google Scholar]
- Ropolo, M.; Geroldi, A.; Rossi, O.; Degan, P.; Zupo, S.; Poggi, A.; Frosina, G. Expression of the Drosophila melanogaster S3 ribosomal/repair protein in T24 human bladder cells. Anticancer Res. 2004, 24, 3811–3818. [Google Scholar]
- Clark, D.E.; Errington, T.M.; Smith, J.A.; Frierson, H.F., Jr; Weber, M.J.; Lannigan, D.A. The serine/threonine protein kinase, p90 ribosomal S6 kinase, is an important regulator of prostate cancer cell proliferation. Cancer Res. 2005, 65, 3108–3116. [Google Scholar]


 : N-glycosylation site;
: N-glycosylation site;
 : Protein kinase C phosphorylation site; □: Tyrosine kinase phosphorylation site;
: Protein kinase C phosphorylation site; □: Tyrosine kinase phosphorylation site;
 : Ribosomal protein L23 signature;
: Ribosomal protein L23 signature;
 : polymorphism site. (Pa: A. melanoleuca; Ho: Homo sapiens; Mu: Mus musculus; Ra: Rattus norvegicus; Ma: Macaca mulatta).
: N-glycosylation site;
: Protein kinase C phosphorylation site; □: Tyrosine kinase phosphorylation site;
: Ribosomal protein L23 signature;
: polymorphism site. (Pa: A. melanoleuca; Ho: Homo sapiens; Mu: Mus musculus; Ra: Rattus norvegicus; Ma: Macaca mulatta).
: polymorphism site. (Pa: A. melanoleuca; Ho: Homo sapiens; Mu: Mus musculus; Ra: Rattus norvegicus; Ma: Macaca mulatta).
: N-glycosylation site;
: Protein kinase C phosphorylation site; □: Tyrosine kinase phosphorylation site;
: Ribosomal protein L23 signature;
: polymorphism site. (Pa: A. melanoleuca; Ho: Homo sapiens; Mu: Mus musculus; Ra: Rattus norvegicus; Ma: Macaca mulatta).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ailuropoda melanoleuca | Homo sapiens | Mus musculus | Rattus norvegicus | Macaca mulatta | |
|---|---|---|---|---|---|
| Molecular weight (kD) | 17.7179 | 17.6919 | 17.6919 | 17.6919 | 17.6919 |
| PI | 11.16 | 11.16 | 11.16 | 11.16 | 11.16 |
| Species | amino acid site | |||||
|---|---|---|---|---|---|---|
| 123 | 126 | 127 | 128 | 129 | 130 | |
| A. melanoleuca | H | H | H | H | H | H |
| Homo sapiens | H | H | H | H | H | H |
| Mus musculus | C | H | H | H | H | H |
| Rattus norvegicus | C | H | H | H | H | H |
| Bos Taurus | H | C | C | C | C | C |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sun, B.; Hou, Y.-L.; Hou, W.-R.; Zhang, S.-N.; Ding, X.; Su, X.-L. cDNA Cloning, Overexpression, Purification and Pharmacologic Evaluation for Anticancer Activity of Ribosomal Protein L23A Gene (RPL23A) from the Giant Panda. Int. J. Mol. Sci. 2012, 13, 2133-2147. https://doi.org/10.3390/ijms13022133
Sun B, Hou Y-L, Hou W-R, Zhang S-N, Ding X, Su X-L. cDNA Cloning, Overexpression, Purification and Pharmacologic Evaluation for Anticancer Activity of Ribosomal Protein L23A Gene (RPL23A) from the Giant Panda. International Journal of Molecular Sciences. 2012; 13(2):2133-2147. https://doi.org/10.3390/ijms13022133
Chicago/Turabian StyleSun, Bing, Yi-Ling Hou, Wan-Ru Hou, Si-Nan Zhang, Xiang Ding, and Xiu-Lan Su. 2012. "cDNA Cloning, Overexpression, Purification and Pharmacologic Evaluation for Anticancer Activity of Ribosomal Protein L23A Gene (RPL23A) from the Giant Panda" International Journal of Molecular Sciences 13, no. 2: 2133-2147. https://doi.org/10.3390/ijms13022133
APA StyleSun, B., Hou, Y. -L., Hou, W. -R., Zhang, S. -N., Ding, X., & Su, X. -L. (2012). cDNA Cloning, Overexpression, Purification and Pharmacologic Evaluation for Anticancer Activity of Ribosomal Protein L23A Gene (RPL23A) from the Giant Panda. International Journal of Molecular Sciences, 13(2), 2133-2147. https://doi.org/10.3390/ijms13022133