Plant Flavonoids—Biosynthesis, Transport and Involvement in Stress Responses

 , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
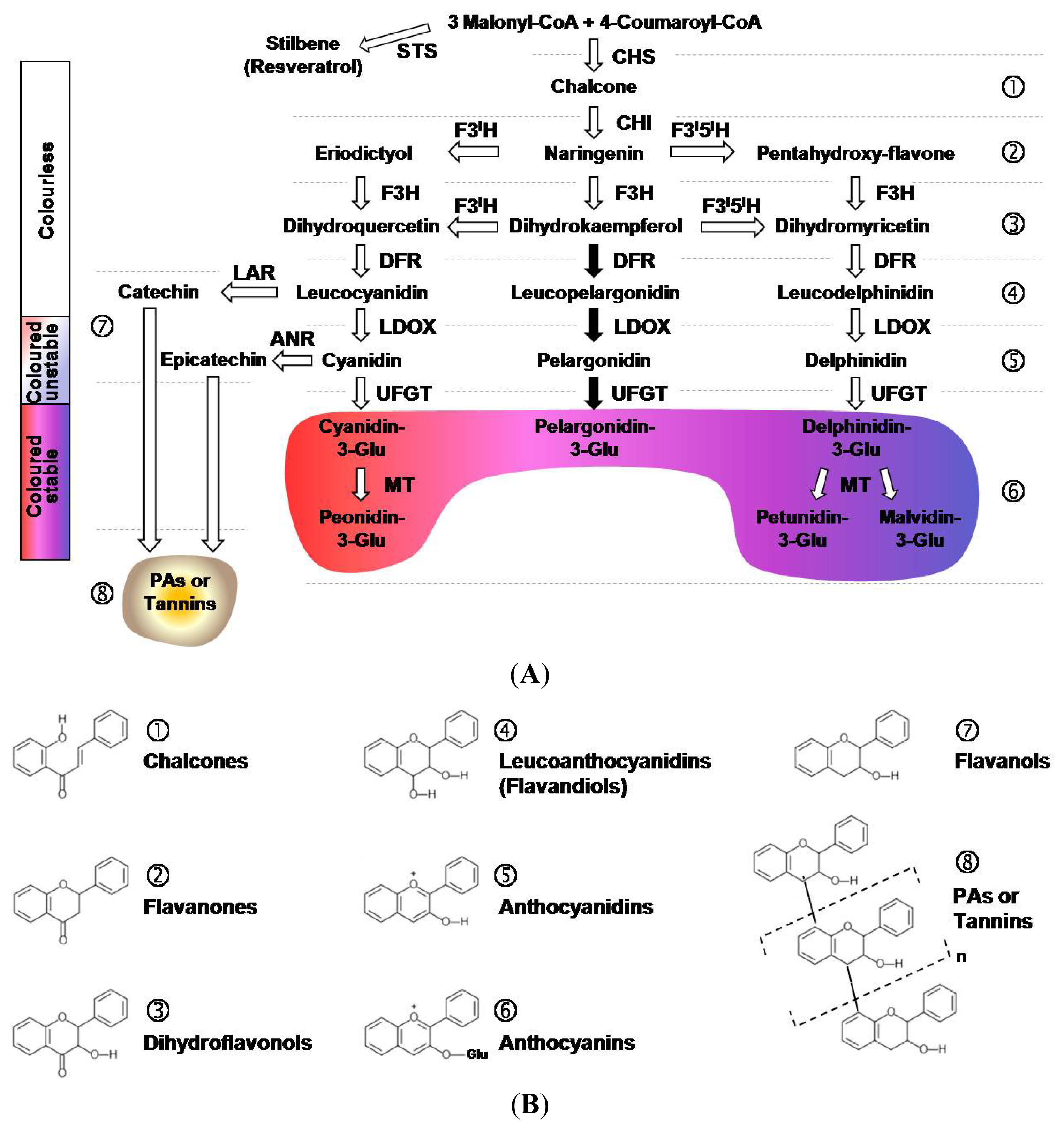
2. Biosynthetic Pathway of Flavonoids in Plant Cells
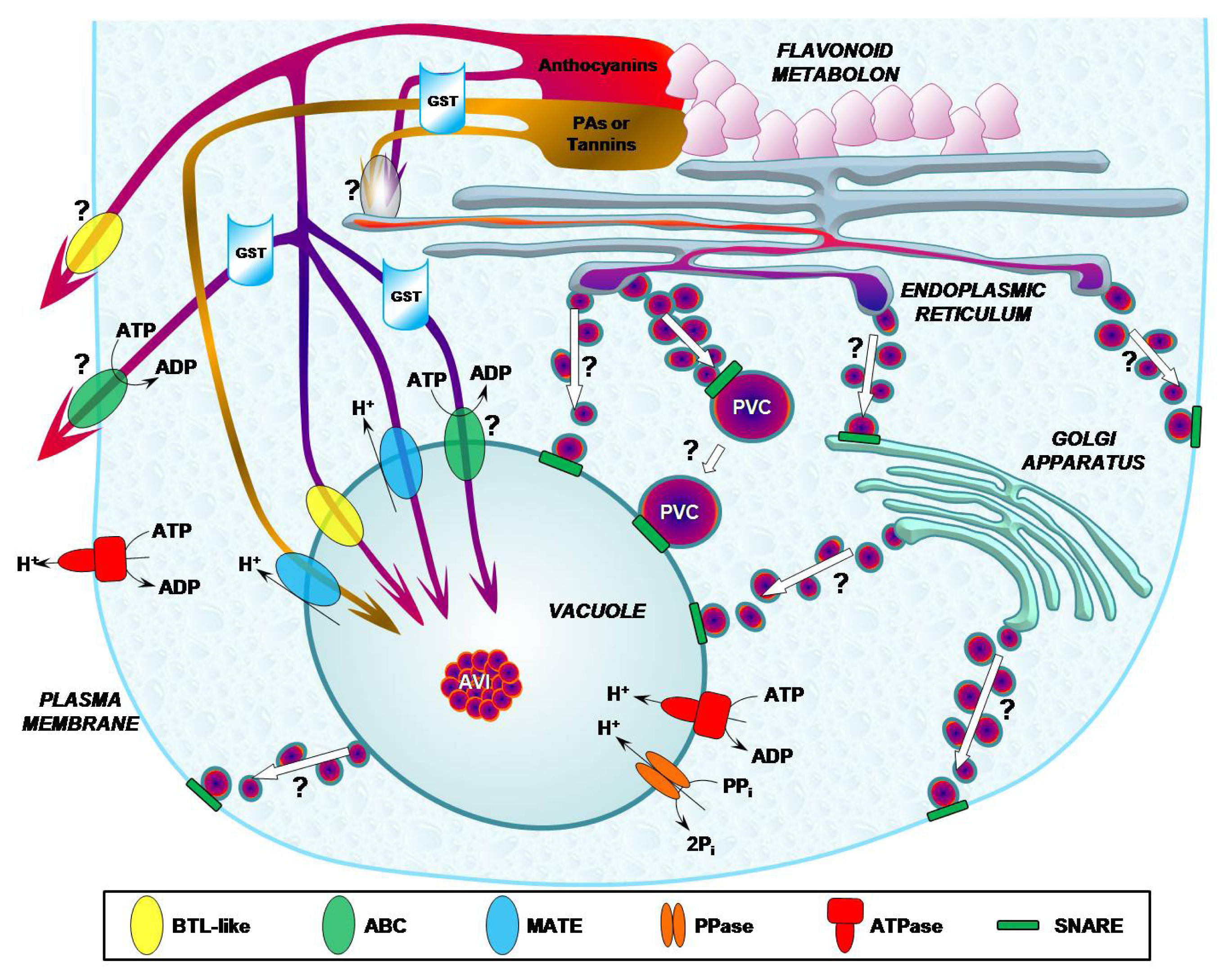
3. Mechanisms of Flavonoid Transport in Plant Cells
4. Genetic Regulation of Flavonoid Transport in Plant Cells
5. Transport Mediated by Vesicle Trafficking in Plant Cells
6. Long Distance Transport of Flavonoids in Plants
7. Mechanism(s) of Flavonoid Transport and Regulation in Grapevine
8. Involvement of Flavonoids during Stress Response in Grape
8.1. Biotic Stress
8.2. Abiotic Stress
8.2.1. Light and UV Stress
8.2.2. Temperature
8.2.3. Water Deficit
9. Conclusions
Acknowledgements
Conflict of Interest
References
- Debeaujon, I.; Peeters, A.J.M.; Leon-Kloosterziel, K.M.; Koornneef, M. The TRANSPARENT TESTA12 gene of Arabidopsis encodes a multidrug secondary transporter-like protein required for flavonoid sequestration in vacuoles of the seed coat endothelium. Plant Cell 2001, 13, 853–871. [Google Scholar]
- Kitamura, S. Transport of Flavonoids: From Cytosolic Synthesis to Vacuolar Accumulation. In Science of Flavonoids; Grotewold, E., Ed.; Springer: Berlin, Germany, 2006; pp. 123–146. [Google Scholar]
- Castellarin, S.D.; Di Gaspero, G.; Marconi, R.; Nonis, A.; Peterlunger, E.; Paillard, S.; Adam-Blondon, A.F.; Testolin, R. Colour variation in red grapevines (Vitis vinifera L.): Genomic organisation, expression of flavonoid 3′-hydroxylase, flavonoid 3′,5′-hydroxylase genes and related metabolite profiling of red cyanidin-/blue delphinidin-based anthocyanins in berry skin. BMC Genom. 2006, 7. [Google Scholar] [CrossRef]
- Conde, C.; Silva, P.; Fontes, N.; Dias, A.C.P.; Tavares, R.M.; Sousa, M.J.; Agasse, A.; Delrot, S.; Geros, H. Biochemical changes throughout grape berry development and fruit and wine quality. Food 2007, 1, 1–22. [Google Scholar]
- Bogs, J.; Ebadi, A.; McDavid, D.; Robinson, S.P. Identification of the flavonoid hydroxylases from grapevine and their regulation during fruit development. Plant Physiol 2006, 140, 279–291. [Google Scholar]
- Springob, K.; Nakajima, J.; Yamazaki, M.; Saito, K. Recent advances in the biosynthesis and accumulation of anthocyanins. Nat. Prod. Rep 2003, 20, 288–303. [Google Scholar]
- Zhao, J.; Huhman, D.; Shadle, G.; He, X.Z.; Sumner, L.W.; Tang, Y.H.; Dixon, R.A. MATE2 mediates vacuolar sequestration of flavonoid glycosides and glycoside malonates in Medicago truncatula. Plant Cell 2011, 23, 1536–1555. [Google Scholar]
- Boss, P.K.; Davies, C.; Robinson, S.P. Analysis of the expression of anthocyanin pathway genes in developing Vitis vinifera L. cv shiraz grape berries and the implications for pathway regulation. Plant Physiol 1996, 111, 1059–1066. [Google Scholar]
- Famiani, F.; Walker, R.P.; Tecsi, L.; Chen, Z.H.; Proietti, P.; Leegood, R.C. An immunohistochemical study of the compartmentation of metabolism during the development of grape (Vitis vinifera L.) berries. J. Exp. Bot 2000, 51, 675–683. [Google Scholar]
- Yazaki, K. Transporters of secondary metabolites. Curr. Opin. Plant Biol 2005, 8, 301–307. [Google Scholar]
- Markham, K.R.; Gould, K.S.; Winefield, C.S.; Mitchell, K.A.; Bloor, S.J.; Boase, M.R. Anthocyanic vacuolar inclusions—Their nature and significance in flower colouration. Phytochemistry 2000, 55, 327–336. [Google Scholar]
- Lecas, M.; Brillouet, J.M. Cell-wall composition of grape berry skins. Phytochemistry 1994, 35, 1241–1243. [Google Scholar]
- Gagne, S.; Saucier, C.; Geny, L. Composition and cellular localization of tannins in cabernet sauvignon skins during growth. J. Agric. Food Chem 2006, 54, 9465–9471. [Google Scholar]
- Grotewold, E. The challenges of moving chemicals within and out of cells: Insights into the transport of plant natural products. Planta 2004, 219, 906–909. [Google Scholar]
- Martinoia, E.; Maeshima, M.; Neuhaus, H.E. Vacuolar transporters and their essential role in plant metabolism. J. Exp. Bot 2007, 58, 83–102. [Google Scholar]
- He, F.; Mu, L.; Yan, G.L.; Liang, N.N.; Pan, Q.H.; Wang, J.; Reeves, M.J.; Duan, C.Q. Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules 2010, 15, 9057–9091. [Google Scholar]
- Conn, S.; Franco, C.; Zhang, W. Characterization of anthocyanic vacuolar inclusions in Vitis vinifera L. cell suspension cultures. Planta 2010, 231, 1343–1360. [Google Scholar]
- Terrier, N.; Glissant, D.; Grimplet, J.; Barrieu, F.; Abbal, P.; Couture, C.; Ageorges, A.; Atanassova, R.; Leon, C.; Renaudin, J.P.; et al. Isogene specific oligo arrays reveal multifaceted changes in gene expression during grape berry (Vitis vinifera L.) development. Planta 2005, 222, 832–847. [Google Scholar]
- Ageorges, A.; Fernandez, L.; Vialet, S.; Merdinoglu, D.; Terrier, N.; Romieu, C. Four specific isogenes of the anthocyanin metabolic pathway are systematically co-expressed with the red colour of grape berries. Plant Sci 2006, 170, 372–383. [Google Scholar]
- Stafford, H.A. Possible multienzyme complexes regulating the formation of C6-C3 phenolic compounds and lignins in higher plants. Recent Adv. Phytochem 1974, 8, 53–79. [Google Scholar]
- Winkel, B.S.J. Metabolic channeling in plants. Annu. Rev. Plant Biol 2004, 55, 85–107. [Google Scholar]
- Kuhn, B.M.; Geisler, M.; Bigler, L.; Ringli, C. Flavonols accumulate asymmetrically and affect auxin transport in Arabidopsis. Plant Physiol 2011, 156, 585–595. [Google Scholar]
- Ono, E.; Fukuchi-Mizutani, M.; Nakamura, N.; Fukui, Y.; Yonekura-Sakakibara, K.; Yamaguchi, M.; Nakayama, T.; Tanaka, T.; Kusumi, T.; Tanaka, Y. Yellow flowers generated by expression of the aurone biosynthetic pathway. Proc. Natl. Acad. Sci. USA 2006, 103, 11075–11080. [Google Scholar]
- Saslowsky, D.E.; Warek, U.; Winkel, B.S.J. Nuclear localization of flavonoid enzymes in Arabidopsis. J. Biol. Chem 2005, 280, 23735–23740. [Google Scholar]
- Toda, K.; Kuroiwa, H.; Senthil, K.; Shimada, N.; Aoki, T.; Ayabe, S.; Shimada, S.; Sakuta, M.; Miyazaki, Y.; Takahashi, R. The soybean F3′H protein is localized to the tonoplast in the seed coat hilum. Planta 2012, 236, 79–89. [Google Scholar]
- Wang, H.L.; Wang, W.; Zhang, P.; Pan, Q.H.; Zhan, J.C.; Huang, W.D. Gene transcript accumulation, tissue and subcellular localization of anthocyanidin synthase (ANS) in developing grape berries. Plant Sci 2010, 179, 103–113. [Google Scholar]
- Tian, L.; Wan, S.B.; Pan, Q.H.; Zheng, Y.J.; Huang, W.D. A novel plastid localization of chalcone synthase in developing grape berry. Plant Sci 2008, 175, 431–436. [Google Scholar]
- Wang, H.L.; Wang, W.; Li, H.; Zhang, P.; Zhan, J.C.; Huang, W.D. Expression and tissue and subcellular localization of anthocyanidin synthase (ANS) in grapevine. Protoplasma 2011, 248, 267–279. [Google Scholar]
- Castellarin, S.D.; di Gaspero, G. Transcriptional control of anthocyanin biosynthetic genes in extreme phenotypes for berry pigmentation of naturally occurring grapevines. BMC Plant Biol. 2007, 7. [Google Scholar] [CrossRef]
- Bogs, J.; Downey, M.O.; Harvey, J.S.; Ashton, A.R.; Tanner, G.J.; Robinson, S.P. Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grape berries and grapevine leaves. Plant Physiol. Biochem 2005, 139, 652–663. [Google Scholar]
- Zhao, J.; Pang, Y.Z.; Dixon, R.A. The mysteries of proanthocyanidin transport and polymerization. Plant Physiol 2010, 153, 437–443. [Google Scholar]
- Pourcel, L.; Routaboul, J.M.; Kerhoas, L.; Caboche, M.; Lepiniec, L.; Debeaujon, I. TRANSPARENT TESTA10 encodes a laccase-like enzyme involved in oxidative polymerization of flavonoids in Arabidopsis seed coat. Plant Cell 2005, 17, 2966–2980. [Google Scholar]
- Gomez, C.; Terrier, N.; Torregrosa, L.; Vialet, S.; Fournier-Level, A.; Verries, C.; Souquet, J.M.; Mazauric, J.P.; Klein, M.; Cheynier, V.; et al. Grapevine MATE-type proteins act as vacuolar H+-dependent acylated anthocyanin transporters. Plant Physiol 2009, 150, 402–415. [Google Scholar]
- Nozue, M.; Yamada, K.; Nakamura, T.; Kubo, H.; Kondo, M.; Nishimura, M. Expression of a vacuolar protein (VP24) in anthocyanin-producing cells of sweet potato in suspension culture. Plant Physiol 1997, 115, 1065–1072. [Google Scholar]
- Martinoia, E.; Klein, M.; Geisler, M.; Bovet, L.; Forestier, C.; Kolukisaoglu, U.; Muller-Rober, B.; Schulz, B. Multifunctionality of plant ABC transporters—More than just detoxifiers. Planta 2002, 214, 345–355. [Google Scholar]
- Klein, M.; Burla, B.; Martinoia, E. The multidrug resistance-associated protein (MRP/ABCC) subfamily of ATP-binding cassette transporters in plants. FEBS Lett 2006, 580, 1112–1122. [Google Scholar]
- Zhao, J.; Dixon, R.A. The ‘ins’ and ‘outs’ of flavonoid transport. Trends Plant Sci 2010, 15, 72–80. [Google Scholar]
- Martinoia, E.; Meyer, S.; de Angeli, A.; Nagy, R. Vacuolar transporters in their physiological context. Annu. Rev. Plant Biol 2012, 63, 183–213. [Google Scholar]
- Kanga, J.; Parka, J.; Choia, H.; Burlab, B.; Kretzschmarb, T.; Leea, Y.; Martinoia, E. Plant ABC transporters. Arabidopsis B. 2011, 9. [Google Scholar] [CrossRef] [Green Version]
- Rea, P.A. Plant ATP-binding cassette transporters. Annu. Rev. Plant Biol 2007, 58, 347–375. [Google Scholar]
- Klein, M.; Martinoia, E.; Hoffmann-Thoma, G.; Weissenbock, G. A membrane-potential dependent ABC-like transporter mediates the vacuolar uptake of rye flavone glucuronides: Regulation of glucuronide uptake by glutathione and its conjugates. Plant J 2000, 21, 289–304. [Google Scholar]
- Goodman, C.D.; Casati, P.; Walbot, V. A multidrug resistance-associated protein involved in anthocyanin transport in Zea mays. Plant Cell 2004, 16, 1812–1826. [Google Scholar]
- Mueller, L.A.; Goodman, C.D.; Silady, R.A.; Walbot, V. AN9, a petunia glutathione S-transferase required for anthocyanin sequestration, is a flavonoid-binding protein. Plant Physiol 2000, 123, 1561–1570. [Google Scholar]
- Marrs, K.A.; Alfenito, M.R.; Lloyd, A.M.; Walbot, V. A glutathione-S-transferase involved in vacuolar transfer encoded by the maize gene Bronze-2. Nature 1995, 375, 397–400. [Google Scholar]
- Alfenito, M.R.; Souer, E.; Goodman, C.D.; Buell, R.; Mol, J.; Koes, R.; Walbot, V. Functional complementation of anthocyanin sequestration in the vacuole by widely divergent glutathione S-transferases. Plant Cell 1998, 10, 1135–1149. [Google Scholar]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci 2005, 10, 236–242. [Google Scholar]
- Larsen, E.S.; Alfenito, M.R.; Briggs, W.R.; Walbot, V. A carnation anthocyanin mutant is complemented by the glutathione S-transferases encoded by maize BZ2 and petunia AN9. Plant Cell Rep 2003, 21, 900–904. [Google Scholar]
- Kitamura, S.; Shikazono, N.; Tanaka, A. TRANSPARENT TESTA 19 is involved in the accumulation of both anthocyanins and proanthocyanidins in arabidopsis. Plant J 2004, 37, 104–114. [Google Scholar]
- Francisco, R.M.; Regalado, A.; Ageorges, A.S.; Burla, B.J.; Bassin, B.; Eisenach, C.; Zarrouk, O.; Vialet, S.; Marlin, T.R.S.; Chaves, M.M.; et al. ABCC1, an ATP binding cassette protein from grape berry, transports anthocyanidin 3-O-glucosides. Plant Cell 2013, 25, 1840–1854. [Google Scholar]
- Zhao, J.; Dixon, R.A. Mate transporters facilitate vacuolar uptake of epicatechin 3′-O-glucoside for proanthocyanidin biosynthesis in Medicago truncatula and Arabidopsis. Plant Cell 2009, 21, 2323–2340. [Google Scholar]
- Mathews, H.; Clendennen, S.K.; Caldwell, C.G.; Liu, X.L.; Connors, K.; Matheis, N.; Schuster, D.K.; Menasco, D.J.; Wagoner, W.; Lightner, J.; et al. Activation tagging in tomato identifies a transcriptional regulator of anthocyanin biosynthesis, modification, and transport. Plant Cell 2003, 15, 1689–1703. [Google Scholar]
- Marinova, K.; Kleinschmidt, K.; Weissenbock, G.; Klein, M. Flavonoid biosynthesis in barley primary leaves requires the presence of the vacuole and controls the activity of vacuolar flavonoid transport. Plant Physiol 2007, 144, 432–444. [Google Scholar]
- Frangne, N.; Eggmann, T.; Koblischke, C.; Weissenbock, G.; Martinoia, E.; Klein, M. Flavone glucoside uptake into barley mesophyll and Arabidopsis cell culture vacuoles. Energization occurs by H+-antiport and ATP-binding cassette-type mechanisms. Plant Physiol 2002, 128, 726–733. [Google Scholar]
- Passamonti, S.; Cocolo, A.; Braidot, E.; Petrussa, E.; Peresson, C.; Medic, N.; Macri, F.; Vianello, A. Characterization of electrogenic bromosulfophthalein transport in carnation petal microsomes and its inhibition by antibodies against bilitranslocase. FEBS J 2005, 272, 3282–3296. [Google Scholar]
- Nicolin, V.; Grill, V.; Micali, F.; Narducci, P.; Passamonti, S. Immunolocalisation of bilitranslocase in mucosecretory and parietal cells of the rat gastric mucosa. J. Mol. Histol 2005, 36, 45–50. [Google Scholar]
- Passamonti, S.; Terdoslavich, M.; Margon, A.; Cocolo, A.; Medic, N.; Micali, F.; Decorti, G.; Franko, M. Uptake of bilirubin into HEPG2 cells assayed by thermal lens spectroscopy—Bilitranslocase. FEBS J 2005, 272, 5522–5535. [Google Scholar]
- Jones, S. An overview of the basic helix-loop-helix proteins. Genome Biol. 2004, 5. [Google Scholar] [CrossRef] [Green Version]
- Heim, M.A.; Jakoby, M.; Werber, M.; Martin, C.; Weisshaar, B.; Bailey, P.C. The basic helix-loop-helix transcription factor family in plants: A genome-wide study of protein structure and functional diversity. Mol. Biol. Evol 2003, 20, 735–747. [Google Scholar]
- Hichri, I.; Barrieu, F.; Bogs, J.; Kappel, C.; Delrot, S.; Lauvergeat, V. Recent advances in the transcriptional regulation of the flavonoid biosynthetic pathway. J. Exp. Bot 2011, 62, 2465–2483. [Google Scholar]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB gene family in Arabidopsis thaliana. Curr. Opin. Plant Biol 2001, 4, 447–456. [Google Scholar]
- Czemmel, S.; Heppel, S.C.; Bogs, J. R2R3 MYB transcription factors: Key regulators of the flavonoid biosynthetic pathway in grapevine. Protoplasma 2012, 249, 109–118. [Google Scholar]
- DeVetten, N.; Quattrocchio, F.; Mol, J.; Koes, R. The AN11 locus controlling flower pigmentation in petunia encodes a novel WD-repeat protein conserved in yeast, plants, and animals. Gen. Dev 1997, 11, 1422–1434. [Google Scholar]
- Quattrocchio, F.; Bauldry, A.; Lepiniec, L.; Grotewold, E. The Regulation of Flavonoid Biosynthesis. In The Science of Flavonoids; Grotewold, E., Ed.; Springer: New York, NY, USA, 2006; pp. 97–122. [Google Scholar]
- Nesi, N.; Jond, C.; Debeaujon, I.; Caboche, M.; Lepiniec, L. The Arabidopsis TT2 gene encodes an R2R3 MYB domain protein that acts as a key determinant for proanthocyanidin accumulation in developing seed. Plant Cell 2001, 13, 2099–2114. [Google Scholar]
- Poustka, F.; Irani, N.G.; Feller, A.; Lu, Y.; Pourcel, L.; Frame, K.; Grotewold, E. A trafficking pathway for anthocyanins overlaps with the endoplasmic reticulum-to-vacuole protein-sorting route in Arabidopsis and contributes to the formation of vacuolar inclusions. Plant Physiol 2007, 145, 1323–1335. [Google Scholar]
- Sun, Y.; Li, H.; Huang, J.R. Arabidopsis TT19 functions as a carrier to transport anthocyanin from the cytosol to tonoplasts. Mol. Plant 2012, 5, 387–400. [Google Scholar]
- Zhang, H.B.; Wang, L.; Deroles, S.; Bennett, R.; Davies, K. New insight into the structures and formation of anthocyanic vacuolar inclusions in flower petals. BMC Plant Biol. 2006, 6. [Google Scholar] [CrossRef]
- Grotewold, E.; Chamberlin, M.; Snook, M.; Siame, B.; Butler, L.; Swenson, J.; Maddock, S.; Clair, G.S.; Bowen, B. Engineering secondary metabolism in maize cells by ectopic expression of transcription factors. Plant Cell 1998, 10, 721–740. [Google Scholar]
- Nozue, M.; Yasuda, H. Occurrence of anthocyanoplasts in cell-suspension cultures of sweet-potato. Plant Cell Rep 1985, 4, 252–255. [Google Scholar]
- Grotewold, E.; Davies, K. Trafficking and sequestration of anthocyanins. Nat. Prod. Commun 2008, 3, 1251–1258. [Google Scholar]
- Snyder, B.A.; Nicholson, R.L. Synthesis of phytoalexins in sorghum as a site-specific response to fungal ingress. Science 1990, 248, 1637–1639. [Google Scholar]
- Mueller, L.A.; Walbot, V. Models for Vacuolar Sequestration of Anthocyanins. In Regulation of Phytochemicals by Molecular Techniques; Romeo, J.T., Saunders, J.A., Matthews, B.F., Eds.; Pergamon: New York, NY, USA, 2001; pp. 297–312. [Google Scholar]
- Lin, Y.; Irani, N.G.; Grotewold, E. Sub-cellular trafficking of phytochemicals explored using auto-fluorescent compounds in maize cells. BMC Plant Biol 2003, 19, 10–12. [Google Scholar]
- Pourcel, L.; Irani, N.G.; Lu, Y.H.; Riedl, K.; Schwartz, S.; Grotewold, E. The formation of anthocyanic vacuolar inclusions in Arabidopsis thaliana and implications for the sequestration of anthocyanin pigments. Mol. Plant 2010, 3, 78–90. [Google Scholar]
- Pecket, R.C.; Small, C.J. Occurrence, location and development of anthocyanoplasts. Phytochemistry 1980, 19, 2571–2576. [Google Scholar]
- Dinter, A.; Berger, E.G. Golgi-disturbing agents. Histochem. Cell Biol 1998, 109, 571–590. [Google Scholar]
- Abrahams, S.; Lee, E.; Walker, A.R.; Tanner, G.J.; Larkin, P.J.; Ashton, A.R. The Arabidopsis TDS4 gene encodes leucoanthocyanidin dioxygenase (LDOX) and is essential for proanthocyanidin synthesis and vacuole development. Plant J 2003, 35, 624–636. [Google Scholar]
- Malsam, J.; Kreye, S.; Sollner, T.H. Membrane fusion: Snares and regulation. Cell. Mol. Life Sci 2008, 65, 2814–2832. [Google Scholar]
- Lipka, V.; Kwon, C.; Panstruga, R. SNARE-ware: The role of SNARE-domain proteins in plant biology. Annu. Rev. Cell Dev. Biol 2007, 23, 147–174. [Google Scholar]
- Meyer, D.; Pajonk, S.; Micali, C.; O’Connell, R.; Schulze-Lefert, P. Extracellular transport and integration of plant secretory proteins into pathogen-induced cell wall compartments. Plant J 2009, 57, 986–999. [Google Scholar]
- Saslowsky, D.; Winkel-Shirley, B. Localization of flavonoid enzymes in Arabidopsis roots. Plant J 2001, 27, 37–48. [Google Scholar]
- Buer, C.S.; Muday, G.K. The TRANSPARENT TESTA4 mutation prevents flavonoid synthesis and alters auxin transport and the response of Arabidopsis roots to gravity and light. Plant Cell 2004, 16, 1191–1205. [Google Scholar]
- Kaltenbach, M.; Schroder, G.; Schmelzer, E.; Lutz, V.; Schroder, J. Flavonoid hydroxylase from Catharanthus roseus: cDNA, heterologous expression, enzyme properties and cell-type specific expression in plants. Plant J 1999, 19, 183–193. [Google Scholar]
- Brown, D.E.; Rashotte, A.M.; Murphy, A.S.; Normanly, J.; Tague, B.W.; Peer, W.A.; Taiz, L.; Muday, G.K. Flavonoids act as negative regulators of auxin transport in vivo in Arabidopsis. Plant Physiol 2001, 126, 524–535. [Google Scholar]
- Subramanian, S.; Stacey, G.; Yu, O. Endogenous isoflavones are essential for the establishment of symbiosis between soybean and Bradyrhizobium japonicum. Plant J 2006, 48, 261–273. [Google Scholar]
- Akiyama, K.; Matsuzaki, K.; Hayashi, H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature 2005, 435, 824–827. [Google Scholar]
- Juszczuk, I.M.; Wiktorowska, A.; Malusa, E.; Rychter, A.M. Changes in the concentration of phenolic compounds and exudation induced by phosphate deficiency in bean plants (Phaseolus vulgaris L.). Plant Soil 2004, 267, 41–49. [Google Scholar]
- Besseau, S.; Hoffmann, L.; Geoffroy, P.; Lapierre, C.; Pollet, B.; Legrand, M. Flavonoid accumulation in Arabidopsis repressed in lignin synthesis affects auxin transport and plant growth. Plant Cell 2007, 19, 148–162. [Google Scholar]
- Peer, W.A.; Murphy, A.S. Flavonoids and auxin transport: Modulators or regulators? Trends Plant Sci 2007, 12, 556–563. [Google Scholar]
- Buer, C.S.; Muday, G.K.; Djordjevic, M.A. Flavonoids are differentially taken up and transported long distances in Arabidopsis. Plant Physiol 2007, 145, 478–490. [Google Scholar]
- Buer, C.S.; Muday, G.K.; Djordjevic, M.A. Implications of long-distance flavonoid movement in Arabidopsis thaliana. Plant Signal. Behav 2008, 3, 415–417. [Google Scholar]
- Lewis, D.R.; Wu, G.S.; Ljung, K.; Spalding, E.P. Auxin transport into cotyledons and cotyledon growth depend similarly on the ABCB19 multidrug resistance-like transporter. Plant J 2009, 60, 91–101. [Google Scholar]
- Gomez, C.; Conejero, G.; Torregrosa, L.; Cheynier, V.; Terrier, N.; Ageorges, A. In vivo grapevine anthocyanin transport involves vesicle-mediated trafficking and the contribution of ANTHOMATE transporters and GST. Plant J 2011, 67, 960–970. [Google Scholar]
- Zhang, W.; Conn, S.; Franco, C. Characterisation of anthocyanin transport and storage in Vitis vinifera L. cv. Gamay freaux cell suspension cultures. J. Biotechnol 2007, 131, S208–S208. [Google Scholar]
- Conn, S.; Curtin, C.; Bezier, A.; Franco, C.; Zhang, W. Purification, molecular cloning, and characterization of glutathione S-transferases (GSTs) from pigmented Vitis vinifera L. cell suspension cultures as putative anthocyanin transport proteins. J. Exp. Bot 2008, 59, 3621–3634. [Google Scholar]
- Cutanda-Perez, M.C.; Ageorges, A.; Gomez, C.; Vialet, S.; Terrier, N.; Romieu, C.; Torregrosa, L. Ectopic expression of VLMYBA1 in grapevine activates a narrow set of genes involved in anthocyanin synthesis and transport. Plant Mol. Biol 2009, 69, 633–648. [Google Scholar]
- Zhang, J.W.; Ma, H.Q.; Feng, J.D.; Zeng, L.; Wang, Z.; Chen, S.W. Grape berry plasma membrane proteome analysis and its differential expression during ripening. J. Exp. Bot 2008, 59, 2979–2990. [Google Scholar]
- Conn, S.; Zhang, W.; Franco, C. Anthocyanic vacuolar inclusions (AVIs) selectively bind acylated anthocyanins in Vitis vinifera L. (grapevine) suspension culture. Biotechnol. Lett 2003, 25, 835–839. [Google Scholar]
- Braidot, E.; Petrussa, E.; Bertolini, A.; Peresson, C.; Ermacora, P.; Loi, N.; Terdoslavich, M.; Passamonti, S.; Macri, F.; Vianello, A. Evidence for a putative flavonoid translocator similar to mammalian bilitranslocase in grape berries (Vitis vinifera L.) during ripening. Planta 2008, 228, 203–213. [Google Scholar]
- Grimplet, J.; Deluc, L.G.; Tillett, R.L.; Wheatley, M.D.; Schlauch, K.A.; Cramer, G.R.; Cushman, J.C. Tissue-specific mRNA expression profiling in grape berry tissues. BMC Genom. 2007, 8. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, A.; Peresson, C.; Petrussa, E.; Braidot, E.; Passamonti, S.; Macri, F.; Vianello, A. Identification and localization of the bilitranslocase homologue in white grape berries (Vitis vinifera L.) during ripening. J. Exp. Bot 2009, 60, 3861–3871. [Google Scholar]
- Zhang, X.Y.; Wang, X.L.; Wang, X.F.; Xia, G.H.; Pan, Q.H.; Fan, R.C.; Wu, F.Q.; Yu, X.C.; Zhang, D.P. A shift of phloem unloading from symplasmic to apoplasmic pathway is involved in developmental onset of ripening in grape berry. Plant Physiol 2006, 142, 220–232. [Google Scholar]
- Klein, M.; Weissenbock, G.; Dufaud, A.; Gaillard, C.; Kreuz, K.; Martinoia, E. Different energization mechanisms drive the vacuolar uptake of a flavonoid glucoside and a herbicide glucoside. J. Biol. Chem 1996, 271, 29666–29671. [Google Scholar]
- Deluc, L.; Barrieu, F.; Marchive, C.; Lauvergeat, V.; Decendit, A.; Richard, T.; Carde, J.P.; Merillon, J.M.; Hamdi, S. Characterization of a grapevine R2R3-MYB transcription factor that regulates the phenylpropanoid pathway. Plant Physiol 2006, 140, 499–511. [Google Scholar]
- Terrier, N.; Torregrosa, L.; Ageorges, A.; Vialet, S.; Verries, C.; Cheynier, V.; Romieu, C. Ectopic expression of vvmybpa2 promotes proanthocyanidin biosynthesis in grapevine and suggests additional targets in the pathway. Plant Physiol 2009, 149, 1028–1041. [Google Scholar]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol 2002, 5, 218–223. [Google Scholar]
- Belhadj, A.; Telef, N.; Saigne, C.; Cluzet, S.; Barrieu, F.; Hamdi, S.; Merillon, J.M. Effect of methyl jasmonate in combination with carbohydrates on gene expression of pr proteins, stilbene and anthocyanin accumulation in grapevine cell cultures. Plant Physiol. Biochem 2008, 46, 493–499. [Google Scholar]
- Zhang, W.; Curtin, C.; Kikuchi, M.; Franco, C. Integration of jasmonic acid and light irradiation for enhancement of anthocyanin biosynthesis in vitis vinifera suspension cultures. Plant Sci 2002, 162, 459–468. [Google Scholar]
- Repka, V.; Fischerova, I.; Silharova, K. Methyl jasmonate is a potent elicitor of multiple defense responses in grapevine leaves and cell-suspension cultures. Biol. Plant 2004, 48, 273–283. [Google Scholar]
- Rozema, J.; vandeStaaij, J.; Bjorn, L.O.; Caldwell, M. UV-b as an environmental factor in plant life: Stress and regulation. Trends Ecol. Evol 1997, 12, 22–28. [Google Scholar]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. The effect of bunch shading on berry development and flavonoid accumulation in Shiraz grapes. Austr. J. Grape Wine Res 2004, 10, 55–73. [Google Scholar]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci 2012, 196, 67–76. [Google Scholar]
- Cortell, J.M.; Kennedy, J.A. Effect of shading on accumulation of flavonoid compounds in (Vitis vinifera L.) pinot noir fruit and extraction in a model system. J. Agric. Food Chem 2006, 54, 8510–8520. [Google Scholar]
- Matus, J.T.; Loyola, R.; Vega, A.; Pena-Neira, A.; Bordeu, E.; Arce-Johnson, P.; Alcalde, J.A. Post-veraison sunlight exposure induces MYB-mediated transcriptional regulation of anthocyanin and flavonol synthesis in berry skins of vitis vinifera. J. Exp. Bot 2009, 60, 853–867. [Google Scholar]
- Koyama, K.; Ikeda, H.; Poudel, P.R.; Goto-Yamamoto, N. Light quality affects flavonoid biosynthesis in young berries of cabernet sauvignon grape. Phytochemistry 2012, 78, 54–64. [Google Scholar]
- Pastore, C.; Zenoni, S.; Fasoli, M.; Pezzotti, M.; Tornielli, G.B.; Filippetti, I. Selective defoliation affects plant growth, fruit transcriptional ripening program and flavonoid metabolism in grapevine. BMC Plant Biol. 2013, 13. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, J.H.; Xin, H.P.; Wang, N.; Guan, L.; Wu, B.H.; Li, S.H. Anthocyanin profile and gene expression in berry skin of two red Vitis vinifera grape cultivars that are sunlight dependent versus sunlight independent. Austr. J. Grape Wine Res 2013, 19, 238–248. [Google Scholar]
- Azuma, A.; Yakushiji, H.; Koshita, Y.; Kobayashi, S. Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 2012, 236, 1067–1080. [Google Scholar]
- Chalker-Scott, L. Environmental significance of anthocyanins in plant stress responses. Photochem. Photobiol 1999, 70, 1–9. [Google Scholar]
- Choi, S.; Kwon, Y.R.; Hossain, M.A.; Hong, S.W.; Lee, B.H.; Lee, H. A mutation in ELA1, an age-dependent negative regulator of PAP1/MYB75, causes UV- and cold stress-tolerance in Arabidopsis thaliana seedlings. Plant Sci 2009, 176, 678–686. [Google Scholar]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J.C. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vit 2002, 53, 171–182. [Google Scholar]
- Mori, K.; Sugaya, S.; Gemma, H. Decreased anthocyanin biosynthesis in grape berries grown under elevated night temperature condition. Sci. Hortic 2005, 105, 319–330. [Google Scholar]
- Yamane, T.; Jeong, S.T.; Goto-Yamamoto, N.; Koshita, Y.; Kobayashi, S. Effects of temperature on anthocyanin biosynthesis in grape berry skins. Am. J. Enol. Vit 2006, 57, 54–59. [Google Scholar]
- Mori, K.; Goto-Yamamoto, N.; Kitayama, M.; Hashizume, K. Loss of anthocyanins in red-wine grape under high temperature. J. Exp. Bot 2007, 58, 1935–1945. [Google Scholar]
- Matthews, M.A.; Anderson, M.M. Fruit ripening in Vitis vinifera L.—Responses to seasonal water deficits. Am. J. Enol. Vit 1988, 39, 313–320. [Google Scholar]
- Ojeda, H.; Andary, C.; Kraeva, E.; Carbonneau, A.; Deloire, A. Influence of pre- and postveraison water deficit on synthesis and concentration of skin phenolic compounds during berry growth of Vitis vinifera cv. Shiraz. Am. J. Enol. Vit 2002, 53, 261–267. [Google Scholar]
- Roby, G.; Harbertson, J.F.; Adams, D.A.; Matthews, M.A. Berry size and vine water deficits as factors in winegrape composition: Anthocyanins and tannins. Austr. J. Grape Wine Res 2004, 10, 100–107. [Google Scholar]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A.L. Effect of maturity and vine water status on grape skin and wine flavonoids. Am. J. Enol. Vit 2002, 53, 268–274. [Google Scholar]
- Castellarin, S.D.; Matthews, M.A.; di Gaspero, G.; Gambetta, G.A. Water deficits accelerate ripening and induce changes in gene expression regulating flavonoid biosynthesis in grape berries. Planta 2007, 227, 101–112. [Google Scholar]


© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Petrussa, E.; Braidot, E.; Zancani, M.; Peresson, C.; Bertolini, A.; Patui, S.; Vianello, A. Plant Flavonoids—Biosynthesis, Transport and Involvement in Stress Responses. Int. J. Mol. Sci. 2013, 14, 14950-14973. https://doi.org/10.3390/ijms140714950
Petrussa E, Braidot E, Zancani M, Peresson C, Bertolini A, Patui S, Vianello A. Plant Flavonoids—Biosynthesis, Transport and Involvement in Stress Responses. International Journal of Molecular Sciences. 2013; 14(7):14950-14973. https://doi.org/10.3390/ijms140714950
Chicago/Turabian StylePetrussa, Elisa, Enrico Braidot, Marco Zancani, Carlo Peresson, Alberto Bertolini, Sonia Patui, and Angelo Vianello. 2013. "Plant Flavonoids—Biosynthesis, Transport and Involvement in Stress Responses" International Journal of Molecular Sciences 14, no. 7: 14950-14973. https://doi.org/10.3390/ijms140714950
APA StylePetrussa, E., Braidot, E., Zancani, M., Peresson, C., Bertolini, A., Patui, S., & Vianello, A. (2013). Plant Flavonoids—Biosynthesis, Transport and Involvement in Stress Responses. International Journal of Molecular Sciences, 14(7), 14950-14973. https://doi.org/10.3390/ijms140714950