Blocking the Function of Inflammatory Cytokines and Mediators by Using IL-10 and TGF-β: A Potential Biological Immunotherapy for Intervertebral Disc Degeneration in a Beagle Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
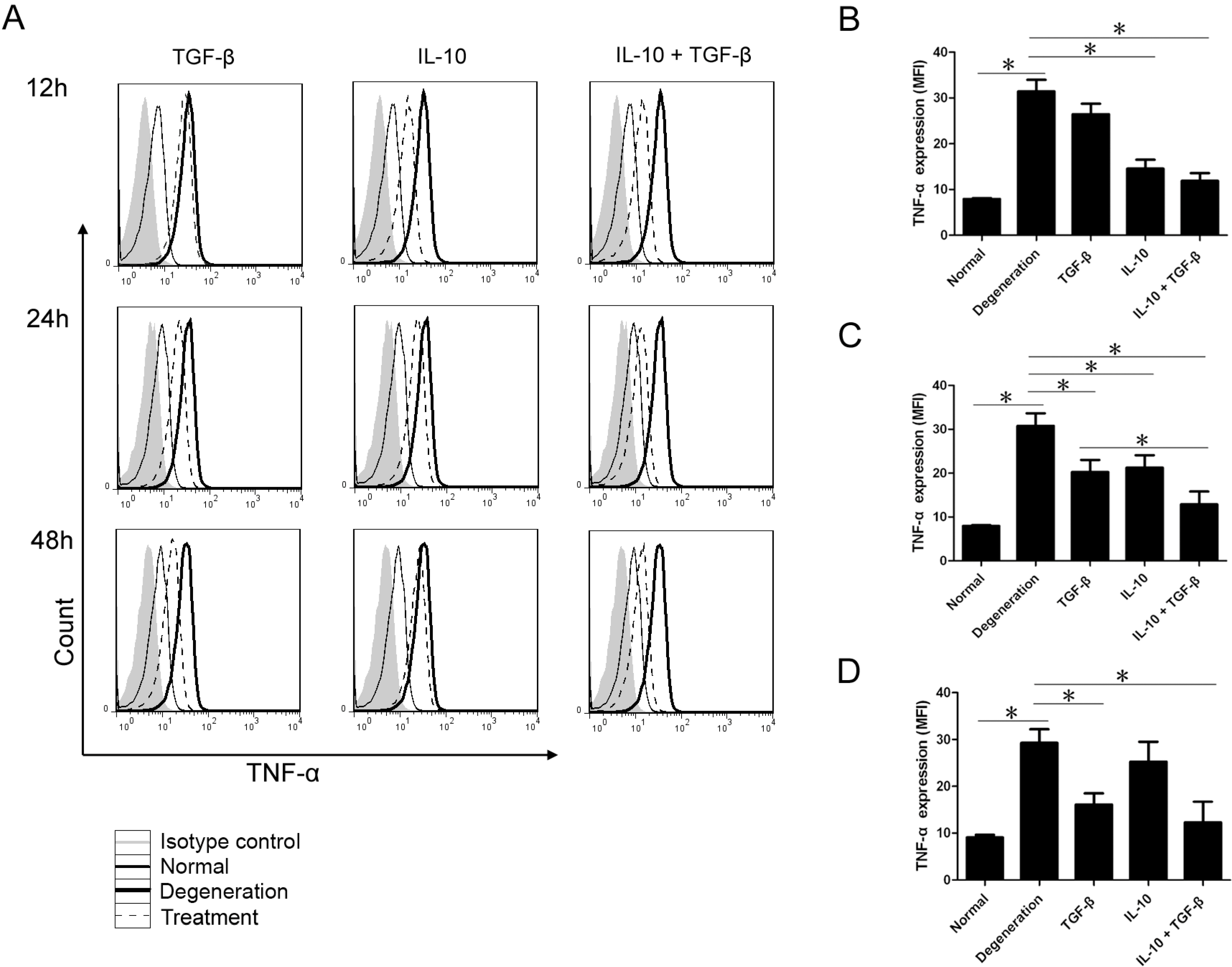
2.1. Intracellular Inflammatory Cytokine Analysis with Flow Cytometry

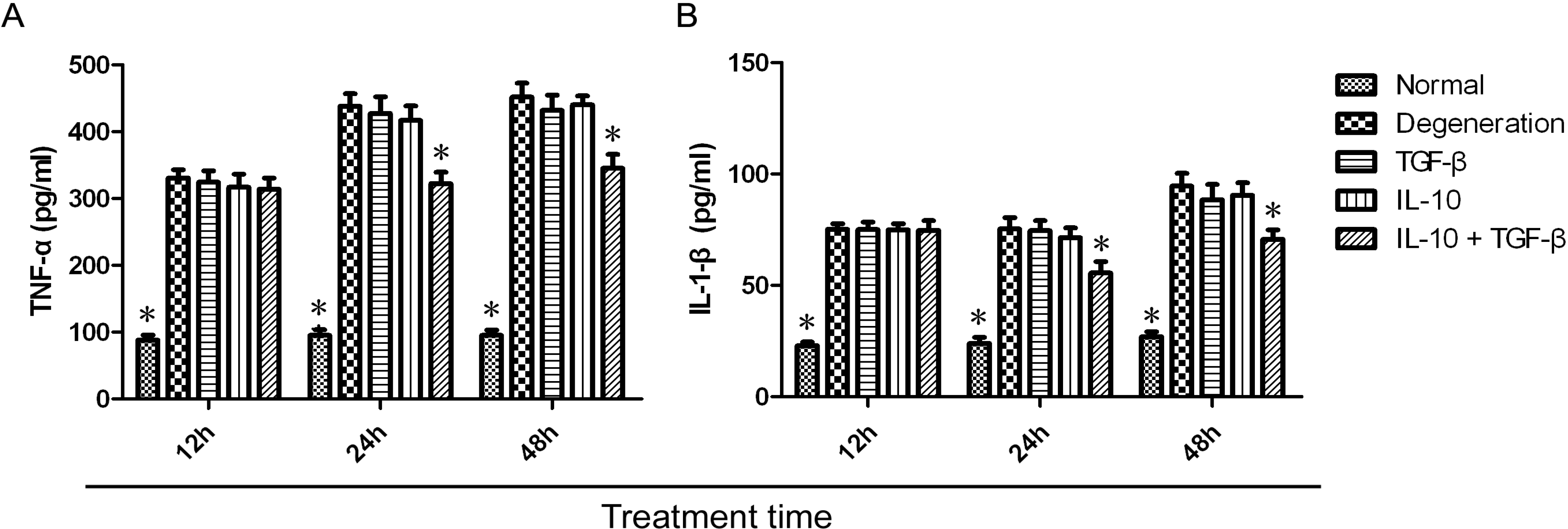
2.2. Quantitative Assay of Cytokines in the Supernatant

2.3. Quantitative Analysis of mRNA Expression

3. Discussion
4. Experimental Section
4.1. Animals
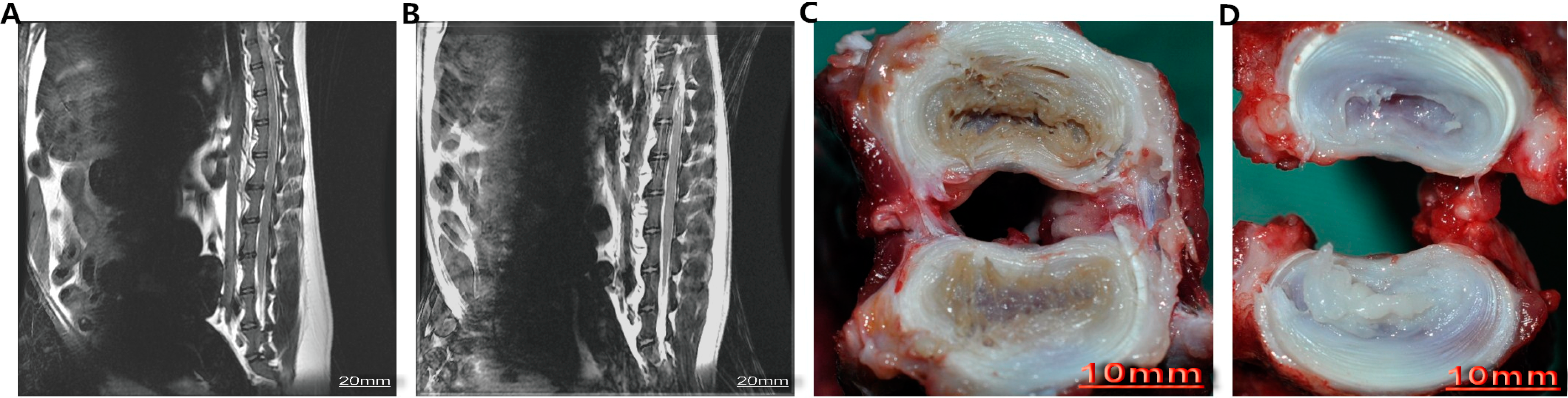
4.2. Animal Model of Intervertebral Disk Degeneration (IDD)

4.3. Isolation and Culture of Nucleus Pulposus (NP) Cells
4.4. Cytokine Treatments and Groups
4.5. Assay of Inflammatory Cytokine-Producing Cells
4.6. Quantitative Assay of Cytokines in the Supernatant
4.7. Real-Time PCR Analysis
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- McBeth, J.; Jones, K. Epidemiology of chronic musculoskeletal pain. Best Pract. Res. Clin. Rheumatol. 2007, 21, 403–425. [Google Scholar]
- Kandel, R.; Roberts, S.; Urban, J.P. Tissue engineering and the intervertebral disc: The challenges. Eur. Spine J. 2008, 17, S480–S491. [Google Scholar]
- Asche, C.V.; Kirkness, C.S.; McAdam-Marx, C.; Fritz, J.M. The societal costs of low back pain: Data published between 2001 and 2007. J. Pain Palliat. Care Pharmacother. 2007, 21, 25–33. [Google Scholar]
- Freemont, A.J. The cellular pathobiology of the degenerate intervertebral disc and discogenic back pain. Rheumatology 2009, 48, 5–10. [Google Scholar]
- Smith, L.J.; Nerurkar, N.L.; Choi, K.S.; Harfe, B.D.; Elliott, D.M. Degeneration and regeneration of the intervertebral disc: Lessons from development. Dis. Model Mech. 2011, 4, 31–41. [Google Scholar]
- Zhao, J.G.; Wang, J.; Xin, Q.; Zhang, P.; Zhang, S.F.; Qi, F.; Mao, D.; Zhang, Z.C. Calcium pentosan polysulfate and sodium pentosan polysulfate may be used to treat intervertebral disc degeneration. Med. Hypotheses 2011, 76, 610–613. [Google Scholar]
- Sakai, D. Future perspectives of cell-based therapy for intervertebral disc disease. Eur. Spine J. 2008, 17, S452–S458. [Google Scholar]
- Podichetty, V.K. The aging spine: The role of inflammatory mediators in intervertebral disc degeneration. Cell Mol. Biol. 2007, 53, 4–18. [Google Scholar]
- Rannou, F.; Richette, P.; Benallaoua, M.; Francois, M.; Genries, V.; Korwin-Zmijowska, C.; Revel, M.; Corvol, M.; Poiraudeau, S. Cyclic tensile stretch modulates proteoglycan production by intervertebral disc annulus fibrosus cells through production of nitrite oxide. J. Cell Biochem. 2003, 90, 148–157. [Google Scholar]
- Ulrich, J.A.; Liebenberg, E.C.; Thuillier, D.U.; Lotz, J.C. Repeated disc injury causes persistent inflammation. Spine 2007, 32, 2812–2819. [Google Scholar]
- Takahashi, H.; Suguro, T.; Okazima, Y.; Moteqi, M.; Okada, Y.; Kakiuchi, T. Inflammatory cytokines in the herniated disc of the lumbar spine. Spine 1996, 21, 218–224. [Google Scholar]
- Seguin, C.A.; Bojarski, M.; Pilliar, R.M.; Roughley, P.J.; Kandel, R.A. Differential regulation of matrix degrading enzymes in a TNFα-induced model of nucleus pulposus tissue degeneration. Matrix Biol. 2006, 25, 409–418. [Google Scholar]
- Wang, J.; Markova, D.; Anderson, D.G.; Zheng, Z.; Shapiro, I.M.; Risbud, M.V. TNF-α and IL-1β promote a disintegrin-like and metalloprotease with thrombospondin type I motif-5-mediated aggrecan degradation through syndecan-4 in intervertebral disc. J. Biol. Chem. 2011, 286, 39738–39749. [Google Scholar]
- Purmessur, D.; Walter, B.A.; Roughley, P.J.; Laudier, D.M.; Hecht, A.C.; Iatridis, J. A role for TNF-α in intervertebral disc degeneration: A non-recoverable catabolic shift. Biochem. Biophys. Res. Commun. 2013, 433, 151–156. [Google Scholar]
- Seguin, C.A.; Pilliar, R.M.; Roughley, P.J.; Kandel, R.A. Tumor necrosis factor-α modulates matrix production and catabolism in nucleus pulposus tissue. Spine 2005, 30, 1940–1948. [Google Scholar]
- Weiler, C.; Nerlich, A.G.; Zipperer, J.; Bachmeier, B.E.; Boos, N. Expression of major matrix metalloproteinases is associated with intervertebral disc degradation and resorption. Eur. Spine J. 2002, 11, 308–320. [Google Scholar]
- Maeda, S.; Kokubun, S. Changes with age in proteoglycan synthesis in cells cultured in vitro from the inner and outer rabbit annulus fibrosus. Responses to interleukin-1 and interleukin-1 receptor antagonist protein. Spine 2000, 25, 166–169. [Google Scholar]
- Zhao, C.Q.; Liu, D.; Li, H.; Jiang, L.S.; Dai, L.Y. Interleukin-1β enhances the effect of serum deprivation on rat annular cell apoptosis. Apoptosis 2007, 12, 2155–2161. [Google Scholar]
- Millward-Sadler, S.J.; Costello, P.W.; Freemont, A.J.; Hoyland, J.A. Regulation of catabolic gene expression in normal and degenerate human intervertebral disc cells: Implications for the pathogenesis of intervertebral disc degeneration. Arthritis Res. Ther. 2009, 11, R65. [Google Scholar]
- Hoyland, J.A.; Le Maitre, C.; Freemont, A.J. Investigation of the role of IL-1 and TNF in matrix degradation in the intervertebral disc. Rheumatology 2008, 47, 809–814. [Google Scholar]
- Risbud, M.V.; Shapiro, I.M. Role of cytokines in intervertebral disc degeneration: Pain and disc content. Nat. Rev. Rheumatol. 2014, 10, 44–56. [Google Scholar]
- De Waal Malefyt, R.; Abrams, J.; Bennett, B.; Figdor, C.G.; de Vries, J.E. Interleukin 10 (IL-10) inhibits cytokine synthesis by human monocytes: An autoregulatory role of IL-10 produced by monocytes. J. Exp. Med. 1991, 174, 1209–1220. [Google Scholar]
- Kühn, R.; Löhler, J.; Rennick, D.; Rajewsky, K.; Müller, W. Interleukin-10-deficient mice develop chronic enterocolitis. Cell 1993, 75, 263–274. [Google Scholar]
- Wise, L.; McCaughan, C.; Tan, C.K.; Mercer, A.A.; Fleming, S.B. Orf virus interleukin-10 inhibits cytokine synthesis in activated human THP-1 monocytes, but onlypartially impairs their proliferation. J. Gen. Virol. 2007, 88, 1677–1682. [Google Scholar]
- Illien-Jünger, S.; Lu, Y.; Purmessur, D.; Mayer, J.E.; Walter, B.A.; Roughley, P.J.; Qureshi, S.A.; Hecht, A.C.; Iatridis, J.C. Dtrimental effects of discectomy on intervertebral disc biology can be decelerated by growth factor treatment during surgery: A large animal organ culture mode. Spine J. 2014. [Google Scholar] [CrossRef]
- Cho, H.; Lee, S.; Park, S.H.; Huang, J.; Hasty, K.A.; Kim, S.J. Synergistic effect of combined growth factors in porcine intervertebral disc degeneration. Connect. Tissue Res. 2013, 54, 181–186. [Google Scholar]
- Abbott, R.D.; Purmessur, D.; Monsey, R.D.; Brigstock, D.R.; Laudier, D.M.; Iatridis, J.C. Degenerative grade affects the responses of human nucleus pulposus cells to link-N, CTGF, and TGFβ3. J. Spinal Disord. Tech. 2013, 26, E86–E94. [Google Scholar]
- Zhang, R.; Ruan, D.; Zhang, C. Effects of TGF-β1 and IGF-1 on proliferation of human nucleus pulposus cells in medium with different serum concentrations. J. Orthop. Surg. 2006, 1, 9. [Google Scholar]
- Nishida, K.; Kang, J.D.; Gilbertson, L.G.; Moon, S.H.; Suh, J.K.; Voqt, M.T.; Robbins, P.D.; Evans, C.H. Modulation of the biologic activity of the rabbit intervertebral disc by gene therapy: An in vivo study of adenovirus-mediated transfer of the human transforming growth factor β1 encoding gene. Spine 1999, 24, 2419–2425. [Google Scholar]
- Maltman, J.; Pragnell, I.B.; Graham, G.J. Specificity and reciprocity in the interactions between TGF-β and macrophage inflammatory protein-1α. J. Immunol. 1996, 156, 1566–1571. [Google Scholar]
- Song, X.Y.; Gu, M.; Jin, W.W.; Klinman, D.M.; Wahl, S.M. Plasmid DNA encoding transforming growth factor-β1 suppresses chronic disease in a streptococcal cell wall-induced arthritis model. J. Clin. Investig. 1998, 101, 2615–2621. [Google Scholar]
- Maeda, H.; Kuwahara, H.; Ichimura, Y.; Ohtsuki, M.; Kurakata, S.; Shiraishi, A. TGF-β enhances macrophage ability to produce IL-10 in normal and tumor-bearing mice. J. Immunol. 1995, 155, 4926–4932. [Google Scholar]
- Wang, S.C.; Ohata, M.; Schrum, L.; Rippe, R.A.; Tsukamoto, H. Expression of interleukin-10 by in vitro and in vivo activated hepatic stellate cells. J. Biol. Chem. 1998, 273, 302–308. [Google Scholar]
- Li, X.; Phillips, F.M.; An, H.S.; Ellman, M.; Thonar, E.J.; Wu, W.; Park, D.; Im, H.J. The action of resveratrol, a phytoestrogen found in grapes, on the intervertebral disc. Spine 2008, 33, 2586–2595. [Google Scholar]
- Moon, H.J.; Kim, J.H.; Lee, H.S.; Chotai, S.; Kang, J.D.; Suh, J.K.; Park, Y.K. Annulus fibrosus cells interact with neuron-like cells to modulate production of growth factors and cytokines in symptomatic disc degeneration. Spine 2012, 37, 2–9. [Google Scholar]
- Tian, Y.; Yuan, W.; Fujita, N.; Wang, J.; Wang, H.; Shapiro, I.M.; Risbud, M.V. Inflammatory cytokines associated with degenerative disc disease control aggrecanase-1 (ADAMTS-4) expression in mucleus pulposus cells through MAPK and NF-κB. Am. J. Pathol. 2013, 182, 2310–2321. [Google Scholar]
- Bogdan, C.; Paik, J.; Vodovotz, Y.; Nathan, C. Contrasting mechanisms for suppression of macrophage cytokine release by transforming growth factor-β and Interleukin-10. J. Biol. Chem. 1992, 267, 23301–23308. [Google Scholar]
- Elliott, D.M.; Yerramalli, C.S.; Beckstein, J.C.; Boxberqer, J.I.; Johannessen, W.; Vresilovic, E.J. The effect of relative needle diameter in puncture and sham injection animal models of degeneration. Spine 2008, 33, 588–596. [Google Scholar]
- Chelberg, M.K.; Banks, G.M.; Geiger, D.F.; Oegema, T.R., Jr. Identification of heterogenous cell populations in normal human intervertebral disc. J. Anat. 1995, 186, 43–53. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Liu, T.; Wu, L.; Chen, C.; Jia, Z.; Bai, X.; Ruan, D. Blocking the Function of Inflammatory Cytokines and Mediators by Using IL-10 and TGF-β: A Potential Biological Immunotherapy for Intervertebral Disc Degeneration in a Beagle Model. Int. J. Mol. Sci. 2014, 15, 17270-17283. https://doi.org/10.3390/ijms151017270
Li W, Liu T, Wu L, Chen C, Jia Z, Bai X, Ruan D. Blocking the Function of Inflammatory Cytokines and Mediators by Using IL-10 and TGF-β: A Potential Biological Immunotherapy for Intervertebral Disc Degeneration in a Beagle Model. International Journal of Molecular Sciences. 2014; 15(10):17270-17283. https://doi.org/10.3390/ijms151017270
Chicago/Turabian StyleLi, Wei, Tianyi Liu, Liangliang Wu, Chun Chen, Zhiwei Jia, Xuedong Bai, and Dike Ruan. 2014. "Blocking the Function of Inflammatory Cytokines and Mediators by Using IL-10 and TGF-β: A Potential Biological Immunotherapy for Intervertebral Disc Degeneration in a Beagle Model" International Journal of Molecular Sciences 15, no. 10: 17270-17283. https://doi.org/10.3390/ijms151017270
APA StyleLi, W., Liu, T., Wu, L., Chen, C., Jia, Z., Bai, X., & Ruan, D. (2014). Blocking the Function of Inflammatory Cytokines and Mediators by Using IL-10 and TGF-β: A Potential Biological Immunotherapy for Intervertebral Disc Degeneration in a Beagle Model. International Journal of Molecular Sciences, 15(10), 17270-17283. https://doi.org/10.3390/ijms151017270