LIM Mineralization Protein-1 Inhibits the Malignant Phenotypes of Human Osteosarcoma Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
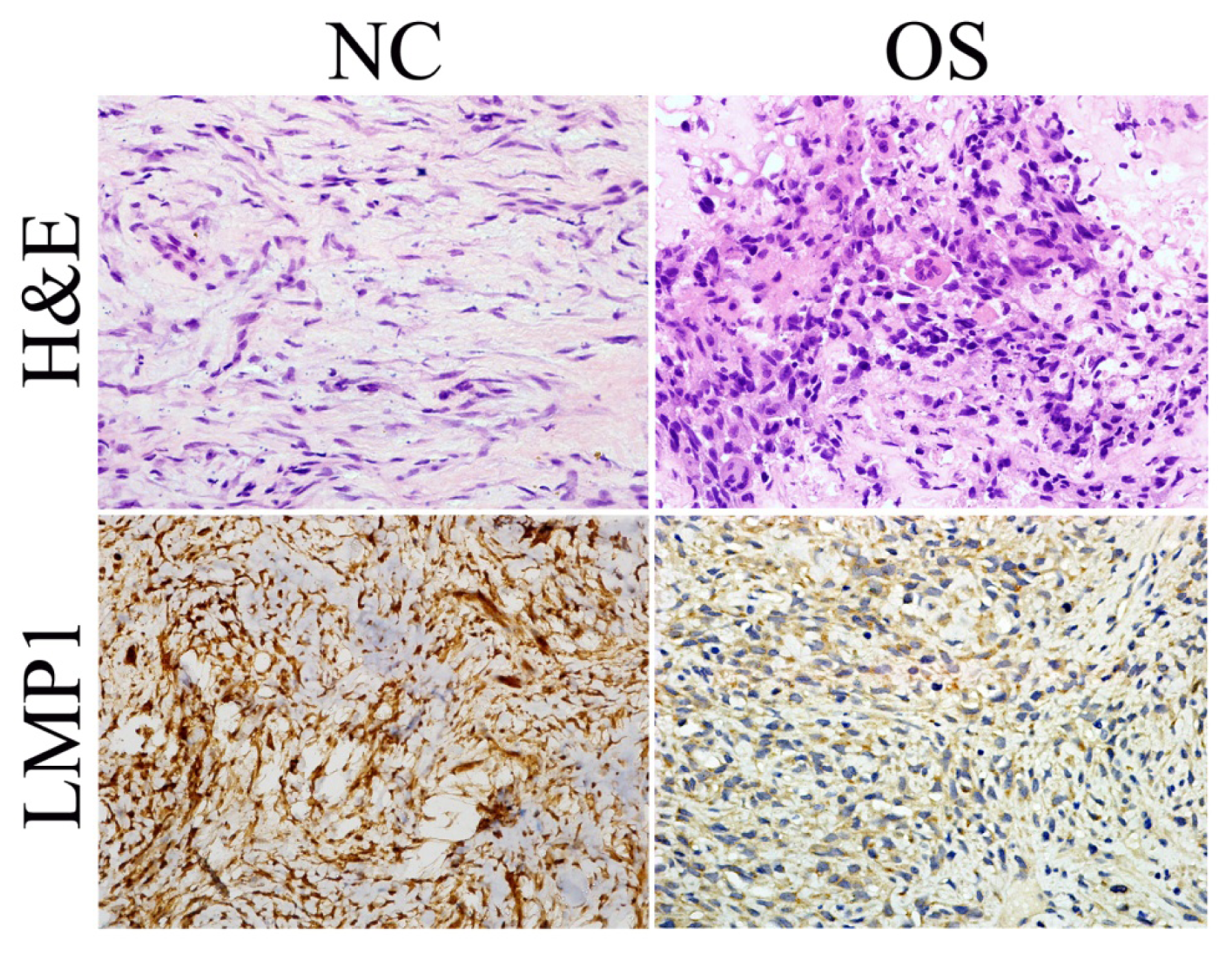
2.1. LMP-1 Expression Is Dysregulated in OS Tissues
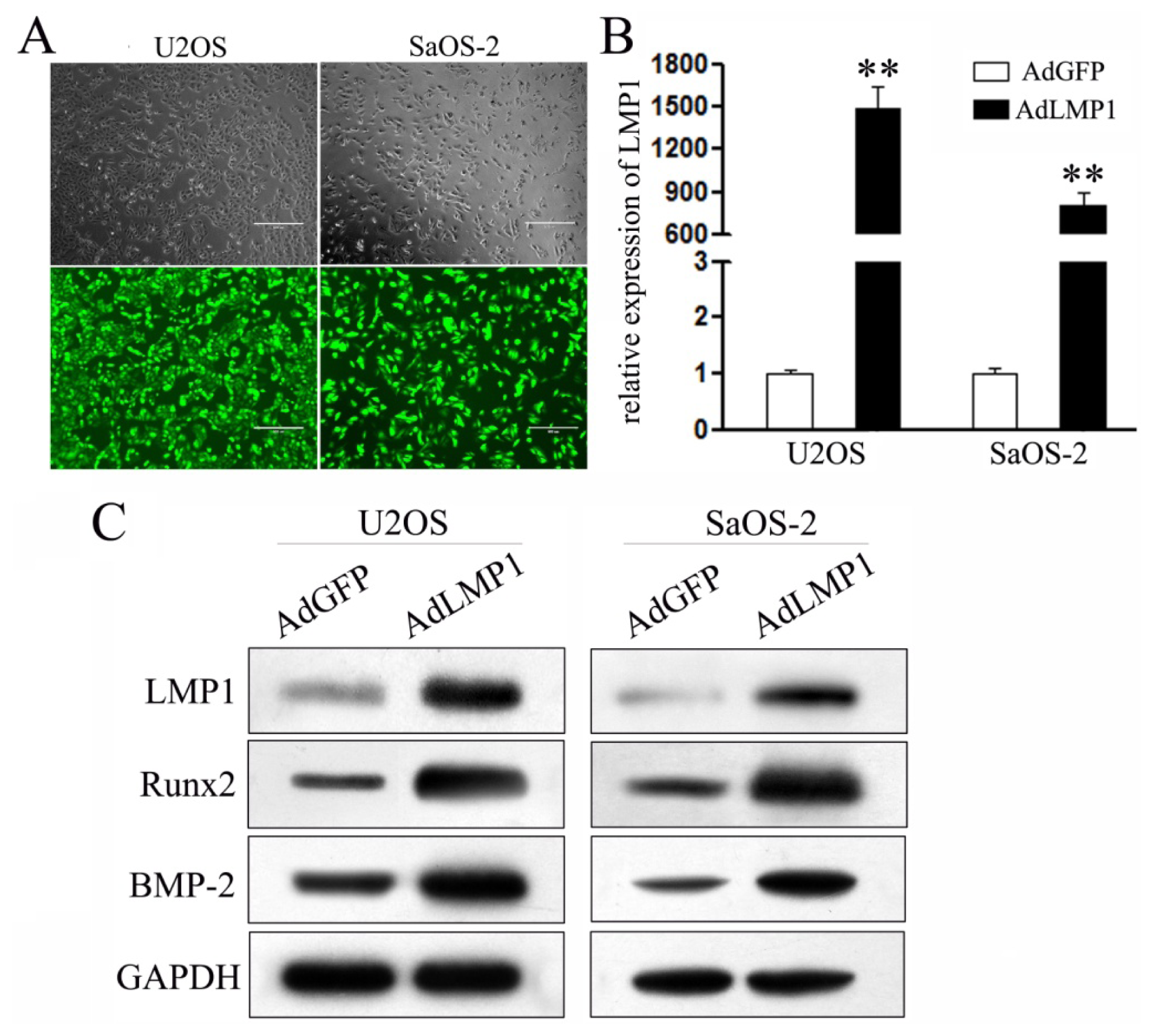
2.2. Restored LMP-1 Expression through Recombinant Adenovirus Infection
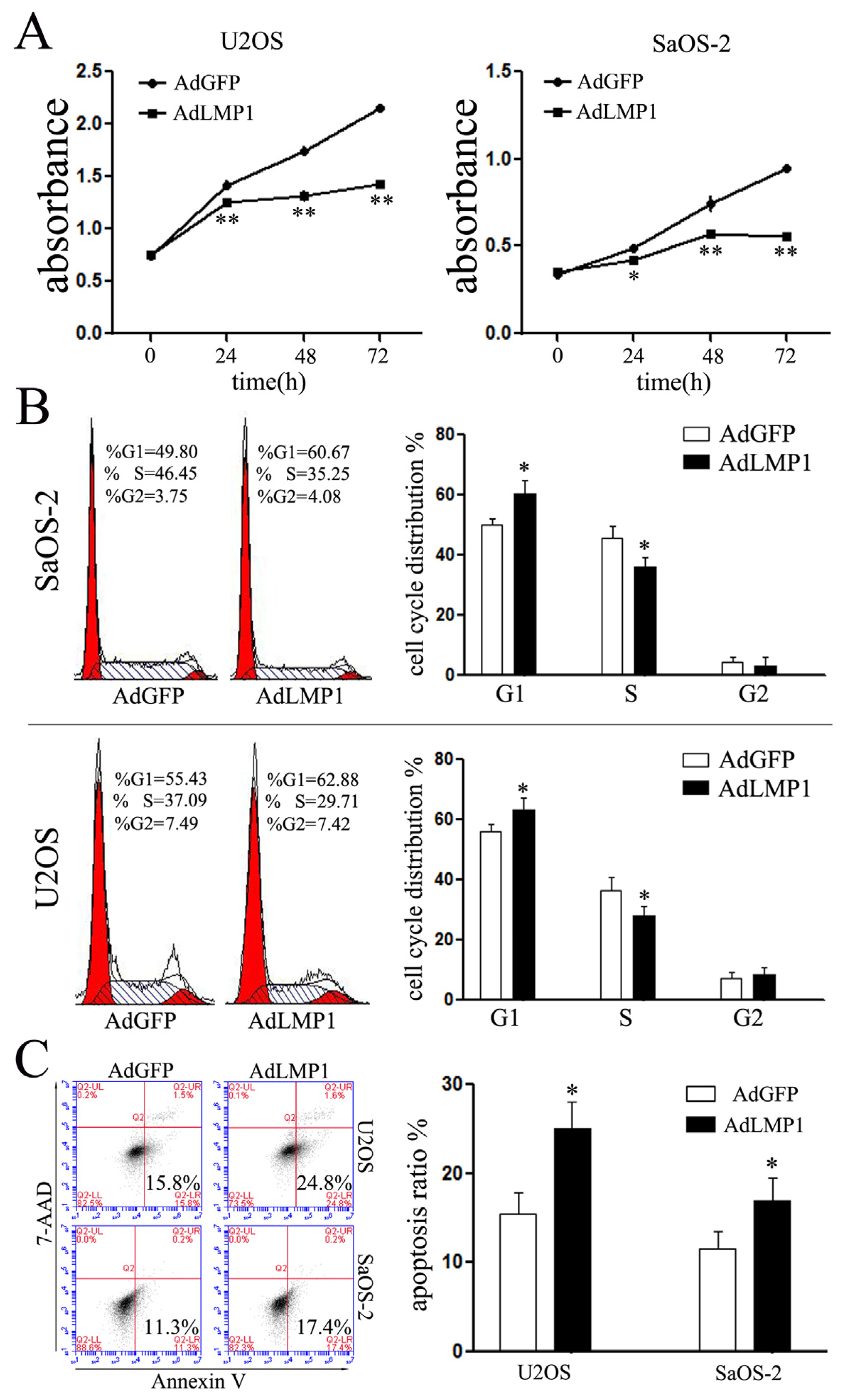
2.3. Increased LMP-1 Inhibits Cell Proliferation and Cell Cycle Progression of OS Cells
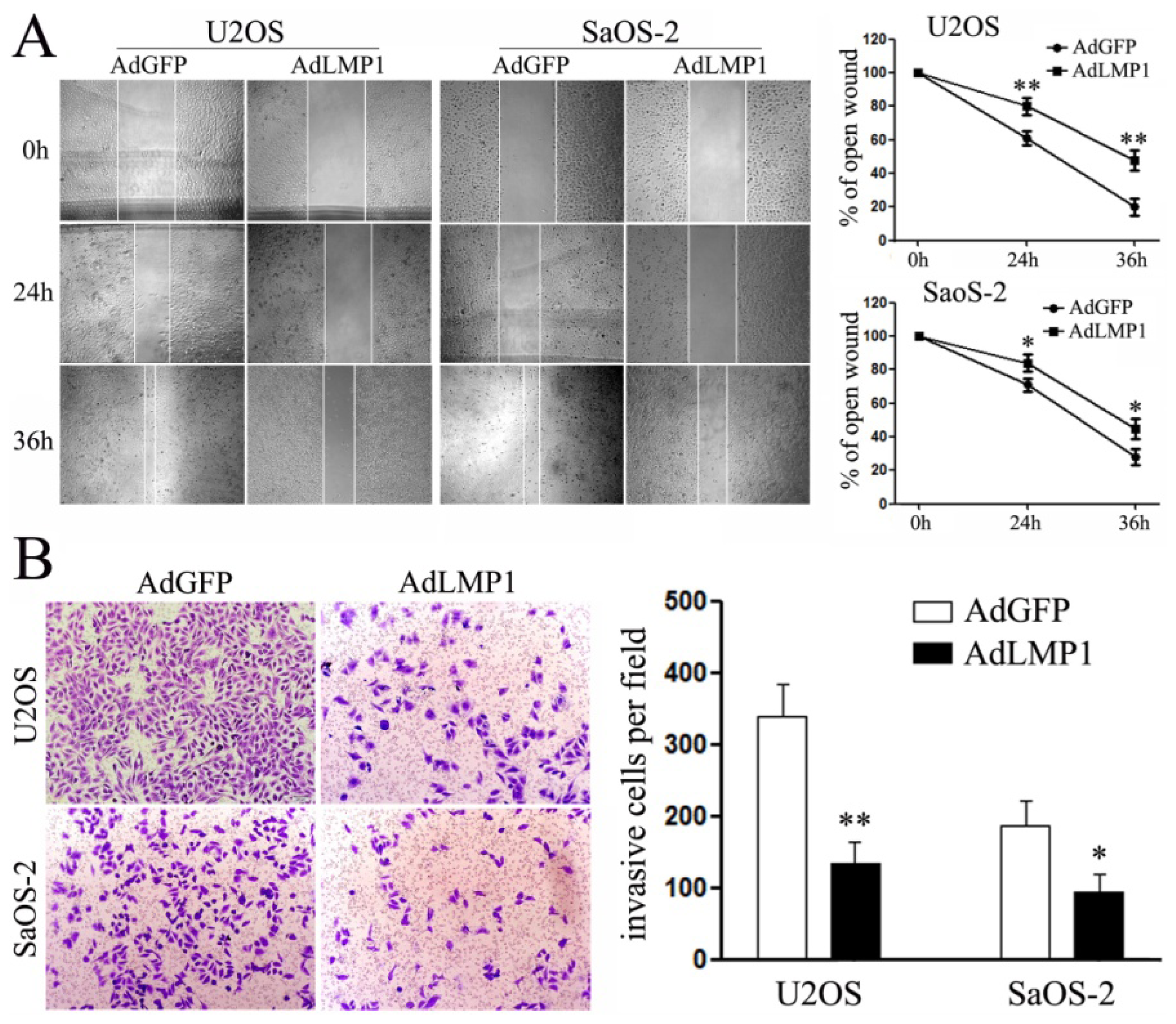
2.4. Ectopic Expression of LMP-1 Inhibits Cell Migration and Invasion of OS Cells
3. Discussion
4. Material and Methods
4.1. Patients and Clinical Samples
4.2. Immunohistochemical Analysis
4.3. Cell Line and Cell Culture
4.4. Recombinant Adenovirus Vector Construction and Infection
4.5. RNA Isolation and Quantitative Real-Time-PCR
4.6. Proliferation Assay
4.7. Cell Cycle Analysis and Apoptosis Assay
4.8. Wound Migration Assay
4.9. Transwell Assays
4.10. Western Blot
4.11. Statistic Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Mirabello, L.; Troisi, R.J.; Savage, S.A. International osteosarcoma incidence patterns in children and adolescents, middle ages and elderly persons. Int. J. Cancer 2009, 125, 229–234. [Google Scholar]
- Gorlick, R.; Khanna, C. Osteosarcoma. J. Bone Miner. Res 2010, 25, 683–691. [Google Scholar]
- Bacci, G.; Longhi, A.; Versari, M.; Mercuri, M.; Briccoli, A.; Picci, P. Prognostic factors for osteosarcoma of the extremity treated with neoadjuvant chemotherapy: 15-Year experience in 789 patients treated at a single institution. Cancer 2006, 106, 1154–1161. [Google Scholar]
- Bacci, G.; Mercuri, M.; Briccoli, A.; Ferrari, S.; Bertoni, F.; Donati, D.; Monti, C.; Zanoni, A.; Forni, C.; Manfrini, M. Osteogenic sarcoma of the extremity with detectable lung metastases at presentation. Results of treatment of 23 patients with chemotherapy followed by simultaneous resection of primary and metastatic lesions. Cancer 1997, 79, 245–254. [Google Scholar]
- Wagner, E.R.; Luther, G.; Zhu, G.; Luo, Q.; Shi, Q.; Kim, S.H.; Gao, J.L.; Huang, E.; Gao, Y.; Yang, K.; et al. Defective osteogenic differentiation in the development of osteosarcoma. Sarcoma 2011, 2011, 325238. [Google Scholar]
- Haydon, R.C.; Luu, H.H.; He, T.C. Osteosarcoma and osteoblastic differentiation: A new perspective on oncogenesis. Clin. Orthop. Relat. Res 2007, 454, 237–246. [Google Scholar]
- Luo, X.; Chen, J.; Song, W.X.; Tang, N.; Luo, J.; Deng, Z.L.; Sharff, K.A.; He, G.; Bi, Y.; He, B.C.; et al. Osteogenic BMPs promote tumor growth of human osteosarcomas that harbor differentiation defects. Lab. Investig 2008, 88, 1264–1277. [Google Scholar]
- Boden, S.D.; Liu, Y.; Hair, G.A.; Helms, J.A.; Hu, D.; Racine, M.; Nanes, M.S.; Titus, L. LMP-1, a LIM-domain protein, mediates BMP-6 effects on bone formation. Endocrinology 1998, 139, 5125–5134. [Google Scholar]
- Fei, Q.; Boden, S.D.; Sangadala, S.; Viggeswarapu, M.; Liu, Y.; Titus, L. Truncated human LMP-1 triggers differentiation of C2C12 cells to an osteoblastic phenotype in vitro. Acta Biochim. Biophys. Sin. 2007, 39, 693–700. [Google Scholar]
- Viggeswarapu, M.; Boden, S.D.; Liu, Y.; Hair, G.A.; Louis-Ugbo, J.; Murakami, H.; Kim, H.S.; Mayr, M.T.; Hutton, W.C.; Titus, L. Adenoviral delivery of LIM mineralization protein-1 induces new-bone formation in vitro and in vivo. J. Bone Jt. Surg. Am. Vol. 2001, 83-A, 364–376. [Google Scholar]
- Strohbach, C.; Kleinman, S.; Linkhart, T.; Amaar, Y.; Chen, S.T.; Mohan, S.; Strong, D. Potential involvement of the interaction between insulin-like growth factor binding protein (IGFBP)-6 and LIM mineralization protein (LMP)-1 in regulating osteoblast differentiation. J. Cell. Biochem 2008, 104, 1890–1905. [Google Scholar]
- Wang, X.; Cui, F.; Madhu, V.; Dighe, A.S.; Balian, G.; Cui, Q. Combined VEGF and LMP-1 delivery enhances osteoprogenitor cell differentiation and ectopic bone formation. Growth Factors 2011, 29, 36–48. [Google Scholar]
- Strohbach, C.A.; Rundle, C.H.; Wergedal, J.E.; Chen, S.T.; Linkhart, T.A.; Lau, K.H.; Strong, D.D. LMP-1 retroviral gene therapy influences osteoblast differentiation and fracture repair: A preliminary study. Calcif. Tissue Int 2008, 83, 202–211. [Google Scholar]
- Marina, N.; Gebhardt, M.; Teot, L.; Gorlick, R. Biology and therapeutic advances for pediatric osteosarcoma. Oncology 2004, 9, 422–441. [Google Scholar]
- Liu, H.; Bargouti, M.; Zughaier, S.; Zheng, Z.; Liu, Y.; Sangadala, S.; Boden, S.D.; Titus, L. Osteoinductive LIM mineralization protein-1 suppresses activation of NF-κB and selectively regulates MAPK pathways in pre-osteoclasts. Bone 2010, 46, 1328–1335. [Google Scholar]
- Yoon, S.T.; Park, J.S.; Kim, K.S.; Li, J.; Attallah-Wasif, E.S.; Hutton, W.C.; Boden, S.D. ISSLS prize winner: LMP-1 upregulates intervertebral disc cell production of proteoglycans and BMPs in vitro and in vivo. Spine 2004, 29, 2603–2611. [Google Scholar]
- Deng, Z.L.; Sharff, K.A.; Tang, N.; Song, W.X.; Luo, J.; Luo, X.; Chen, J.; Bennett, E.; Reid, R.; Manning, D.; et al. Regulation of osteogenic differentiation during skeletal development. Front. Biosci 2008, 13, 2001–2021. [Google Scholar]
- James, A.W. Review of signaling pathways governing MSC osteogenic and adipogenic differentiation. Scientifica 2013, 2013, 684736. [Google Scholar]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.H.; Inada, M.; et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 1997, 89, 755–764. [Google Scholar]
- Wagner, D.O.; Sieber, C.; Bhushan, R.; Borgermann, J.H.; Graf, D.; Knaus, P. BMPs: From bone to body morphogenetic proteins. Sci. Signal 2010, 3, mr1. [Google Scholar] [CrossRef]
- Sieber, C.; Kopf, J.; Hiepen, C.; Knaus, P. Recent advances in BMP receptor signaling. Cytokine Growth Factor Rev 2009, 20, 343–355. [Google Scholar]
- Tim Yoon, S.; Kim, K.S.; Li, J.; Park, J.S.; Akamaru, T.; Elmer, W.A.; Hutton, W.C. The effect of bone morphogenetic protein-2 on rat intervertebral disc cellsin vitro. Spine 2003, 28, 1773–1780. [Google Scholar]
- Liu, H.W.; Huang, L.; Han, Z.M.; Luo, J.P.; Zhan, P.; Dai, M.; Cao, K. Construction of recombinant adenoviral vector encoding LMP-1 and its expression in osteosarcoma cells. J. Chognqing Med. Univ 2012, 37, 589–594. [Google Scholar]
- Kozak, M. An analysis of 5′-noncoding sequences from 699 vertebrate messenger RNAs. Nucleic Acids Res 1987, 15, 8125–8148. [Google Scholar]
- Kang, Q.; Song, W.X.; Luo, Q.; Tang, N.; Luo, J.; Luo, X.; Chen, J.; Bi, Y.; He, B.C.; Park, J.K.; et al. A comprehensive analysis of the dual roles of BMPs in regulating adipogenic and osteogenic differentiation of mesenchymal progenitor cells. Stem Cells Dev 2009, 18, 545–559. [Google Scholar]




© 2014 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, H.; Huang, L.; Zhang, Z.; Zhang, Z.; Yu, Z.; Chen, X.; Chen, Z.; Zen, Y.; Yang, D.; Han, Z.; et al. LIM Mineralization Protein-1 Inhibits the Malignant Phenotypes of Human Osteosarcoma Cells. Int. J. Mol. Sci. 2014, 15, 7037-7048. https://doi.org/10.3390/ijms15047037
Liu H, Huang L, Zhang Z, Zhang Z, Yu Z, Chen X, Chen Z, Zen Y, Yang D, Han Z, et al. LIM Mineralization Protein-1 Inhibits the Malignant Phenotypes of Human Osteosarcoma Cells. International Journal of Molecular Sciences. 2014; 15(4):7037-7048. https://doi.org/10.3390/ijms15047037
Chicago/Turabian StyleLiu, Huiwen, Lu Huang, Zhongzu Zhang, Zhanming Zhang, Zhiming Yu, Xiang Chen, Zhuo Chen, Yongping Zen, Dong Yang, Zhimin Han, and et al. 2014. "LIM Mineralization Protein-1 Inhibits the Malignant Phenotypes of Human Osteosarcoma Cells" International Journal of Molecular Sciences 15, no. 4: 7037-7048. https://doi.org/10.3390/ijms15047037
APA StyleLiu, H., Huang, L., Zhang, Z., Zhang, Z., Yu, Z., Chen, X., Chen, Z., Zen, Y., Yang, D., Han, Z., Shu, Y., Dai, M., & Cao, K. (2014). LIM Mineralization Protein-1 Inhibits the Malignant Phenotypes of Human Osteosarcoma Cells. International Journal of Molecular Sciences, 15(4), 7037-7048. https://doi.org/10.3390/ijms15047037