
Evaluation and Selection of Appropriate Reference Genes for Real-Time Quantitative PCR Analysis of Gene Expression in Nile Tilapia (Oreochromis niloticus) during Vaccination and Infection


Abstract
:1. Introduction
2. Results
2.1. qPCR Amplification of Candidate Reference Genes and Target Gene


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5'→3') | Amplification Size (bp) | PCR Efficiency (%) | Correlation Coefficients |
|---|---|---|---|---|
| 18S rRNA-F | GGACACGGAAAGGATTGACAG | 111 | 101.1 | 0.997 |
| 18S rRNA-R | GTTCGTTATCGGAATTAACCAGAC | |||
| ACTB-F | GACCCACACAGTGCCCATCT | 140 | 97.7 | 0.998 |
| ACTB-R | TCTCGGCTGTGGTGGTGAA | |||
| TUBA-F | AGCCAGACGGACAGATGCC | 153 | 92.4 | 0.992 |
| TUBA-R | TTCCTGCACGCACCTCATC | |||
| UBCE-F | GCGGACAGCTTTGGAGATGA | 108 | 93.6 | 0.994 |
| UBCE-R | CGGCAGAGAGTTAGACAAAATCG | |||
| GAPDH-F | GATAATGGCAAACTTGTCGTCG | 205 | 92.8 | 0.993 |
| GAPDH-R | ACATTGGAGCATCGGGTGAG | |||
| EF1A-F | GCACGCTCTGCTGGCCTTT | 250 | 94.2 | 0.994 |
| EF1A-R | GCGCTCAATCTTCCATCCC | |||
| IgM-F | GGGAAGATGAGGAAGGAAATGA | 120 | 91.7 | 0.997 |
| IgM-R | GTTTTACCCCCCTGGTCCAT |
2.2. Expression Levels of Reference Genes in Different Periods of Vaccination and Infection

| Group | Time | Cq Values (Mean ± SD) | ||||||
|---|---|---|---|---|---|---|---|---|
| 18S rRNA | ACTB | TUBA | UBCE | GAPDH | EF1A | IgM | ||
| rSip | 0 a | 11.20 ± 0.08 | 26.45 ± 0.18 | 30.53 ± 1.52 | 31.31 ± 0.77 | 18.95 ± 1.30 | 19.51 ± 0.74 | 29.82 ± 0.71 |
| 1w b | 10.48 ± 0.22 * | 23.46 ± 0.15 * | 28.12 ± 0.38 | 30.22 ± 0.26 | 19.38 ± 1.06 | 17.86 ± 0.81 * | 26.29 ± 0.07 * | |
| 2w b | 11.45 ± 0.15 * | 25.18 ± 0.14 * | 28.50 ± 0.45 | 30.00 ± 0.28 * | 20.71 ± 0.90 * | 18.92 ± 0.70 * | 27.62 ± 0.21 * | |
| 3w b | 10.48 ± 0.10 * | 23.61 ± 0.17 * | 28.13 ± 0.36 | 29.53 ± 0.35 * | 22.26 ± 0.33 * | 18.79 ± 0.61 * | 28.47 ± 0.18 * | |
| 4w b | 10.70 ± 0.10 * | 27.32 ± 0.18 * | 30.90 ± 1.24 | 31.31 ± 0.38 | 21.53 ± 0.85 * | 20.36 ± 0.35 * | 29.40 ± 0.31 | |
| C24h c | 12.20 ± 0.11 * | 27.41 ± 0.33 * | 32.59 ± 0.55 | 29.60 ± 0.21 * | 22.75 ± 1.54 * | 21.32 ± 0.68 * | 28.40 ± 0.43 * | |
| C2w c | 11.31 ± 0.07 | 25.87 ± 0.13 * | 30.87 ± 0.45 | 29.91 ± 1.06 | 21.38 ± 1.04 * | 20.65 ± 1.14 * | 29.05 ± 1.29 | |
| PBS | 0 a | 11.20 ± 0.08 | 26.45 ± 0.18 | 30.53 ± 1.52 | 31.31 ± 0.77 | 18.95 ± 1.30 | 19.51 ± 0.74 | 29.82 ± 0.71 |
| 1w b | 10.71 ± 0.08 * | 25.34 ± 0.15 * | 30.56 ± 2.08 | 31.10 ± 0.32 | 18.48 ± 0.75 | 18.62 ± 1.28 * | 28.00 ± 0.12 * | |
| 2w b | 10.54 ± 0.06 * | 24.05 ± 0.04 * | 29.66 ± 0.70 | 30.00 ± 0.23 * | 19.62 ± 1.12 * | 18.35 ± 0.72 * | 28.30 ± 0.17 * | |
| 3w b | 11.13 ± 0.11 | 26.41 ± 0.13 | 30.64 ± 1.07 | 30.96 ± 0.37 | 21.41 ± 0.94 * | 19.43 ± 0.93 | 28.65 ± 0.19 * | |
| 4w b | 10.80 ± 0.45 | 25.63 ± 0.52 * | 29.16 ± 0.83 | 31.30 ± 0.74 | 20.93 ± 1.47 * | 20.53 ± 0.66 * | 29.42 ± 0.61 | |
| C24h c | 12.75 ± 0.27 * | 27.33 ± 0.46 * | 33.92 ± 1.75 * | 31.80 ± 0.81 | 22.68 ± 0.63 * | 21.36 ± 1.04 * | 29.08 ± 0.70 | |
| C2w c | 12.36 ± 0.82 | 27.06 ± 0.67 | 31.99 ± 1.39 | 32.40 ± 1.52 | 21.36 ± 0.76 * | 20.12 ± 0.49 * | 29.64 ± 1.08 | |
2.3. Determining the Expression Stability of Reference Genes during Vaccination and Infection
| Group | Rank | Method | ||||
|---|---|---|---|---|---|---|
| Comparative ΔCt (Average of SD) | BestKeeper (SD (±CP)) | NormFinder (Stability Value) | geNorm (Stability Value M) | RefFinder (Overall Final Ranking) | ||
| All groups | 1 | EF1A (0.98) | 18S rRNA (0.56) | EF1A (0.34) | EF1A/18S rRNA (0.77) | EF1A (1.32) |
| 2 | 18S rRNA (1.01) | UBCE (0.76) | 18S rRNA (0.43) | - | 18S rRNA (1.41) | |
| 3 | ACTB (1.07) | EF1A (0.91) | ACTB (0.64) | ACTB (0.84) | ACTB (3.22) | |
| 4 | TUBA (1.26) | ACTB (1.10) | TUBA (0.99) | TUBA (0.95) | UBCE (3.98) | |
| 5 | UBCE (1.29) | GAPDH (1.20) | UBCE (1.04) | UBCE (1.04) | TUBA (4.43) | |
| 6 | GAPDH (1.47) | TUBA (1.23) | GAPDH (1.30) | GAPDH (1.18) | GAPDH (5.73) | |
| rSip group | 1 | EF1A (1.01) | 18S rRNA (0.48) | EF1A (0.35) | ACTB/TUBA (0.70) | EF1A (1.73) |
| 2 | 18S rRNA (1.20) | UBCE (0.60) | 18S rRNA (0.66) | - | 18S rRNA (2.00) | |
| 3 | ACTB (1.23) | EF1A (0.98) | ACTB (0.85) | EF1A (0.74) | ACTB (2.59) | |
| 4 | TUBA (1.26) | GAPDH (1.13) | TUBA (0.93) | 18S rRNA (0.97) | TUBA (3.13) | |
| 5 | UBCE (1.54) | ACTB (1.31) | UBCE (1.29) | UBCE (1.15) | UBCE (3.98) | |
| 6 | GAPDH (1.63) | TUBA (1.46) | GAPDH (1.42) | GAPDH (1.31) | GAPDH (5.42) | |
| PBS group | 1 | 18S rRNA (0.74) | UBCE (0.50) | 18S rRNA (0.08) | 18S rRNA/UBCE (0.51) | 18S rRNA (1.19) |
| 2 | ACTB (0.84) | 18S rRNA (0.69) | ACTB (0.47) | - | UBCE (2.00) | |
| 3 | EF1A (0.87) | EF1A (0.83) | EF1A (0.52) | ACTB (0.57) | ACTB (2.63) | |
| 4 | UBCE (0.89) | ACTB (0.88) | UBCE (0.64) | EF1A (0.65) | EF1A (3.22) | |
| 5 | TUBA (1.15) | TUBA (1.16) | TUBA (0.99) | TUBA (0.83) | TUBA (5.00) | |
| 6 | GAPDH (1.17) | GAPDH (1.26) | GAPDH (1.01) | GAPDH (0.94) | GAPDH (6.00) | |
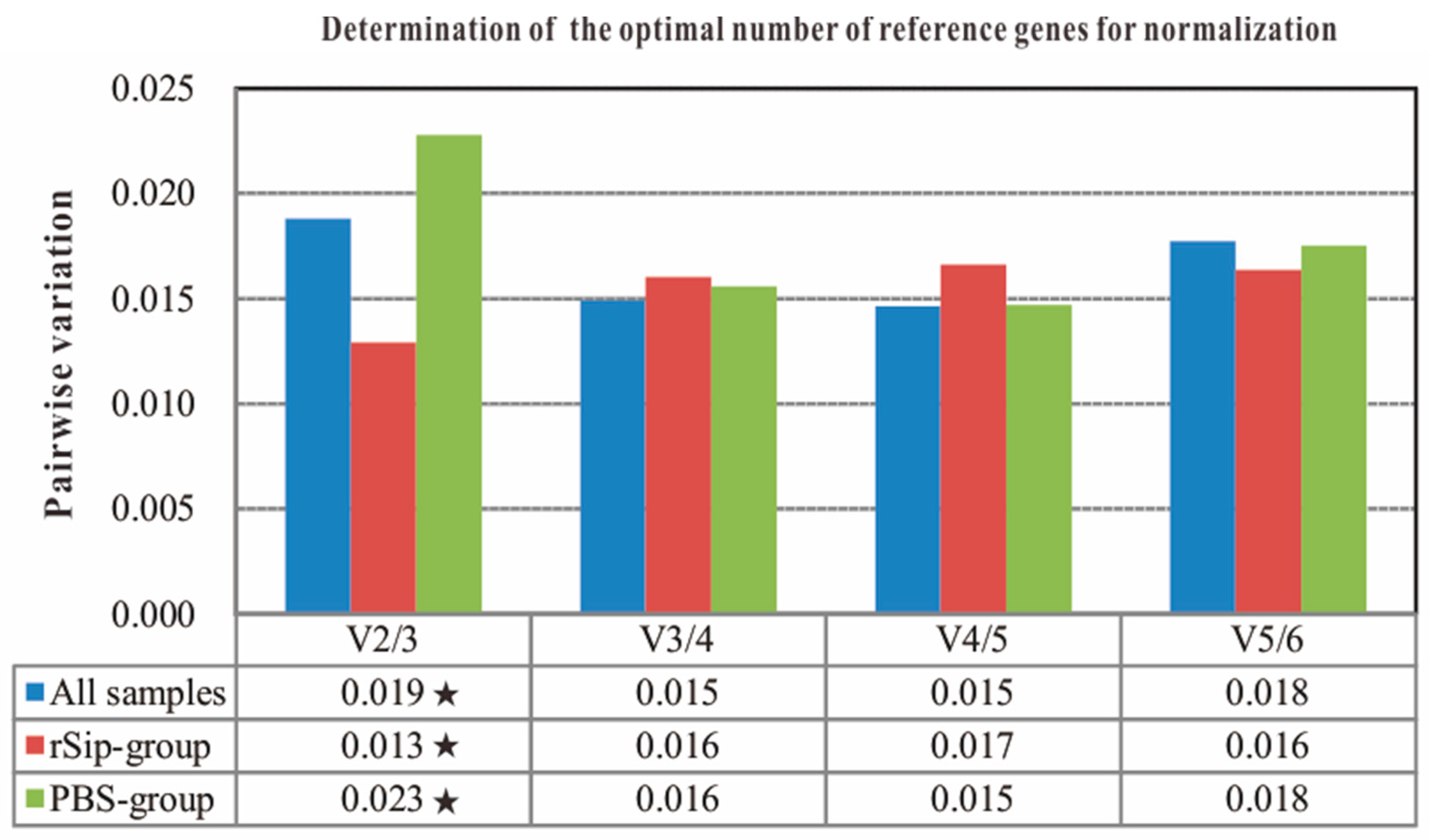
2.4. The Number of Reference Genes Required for Accurate Normalization

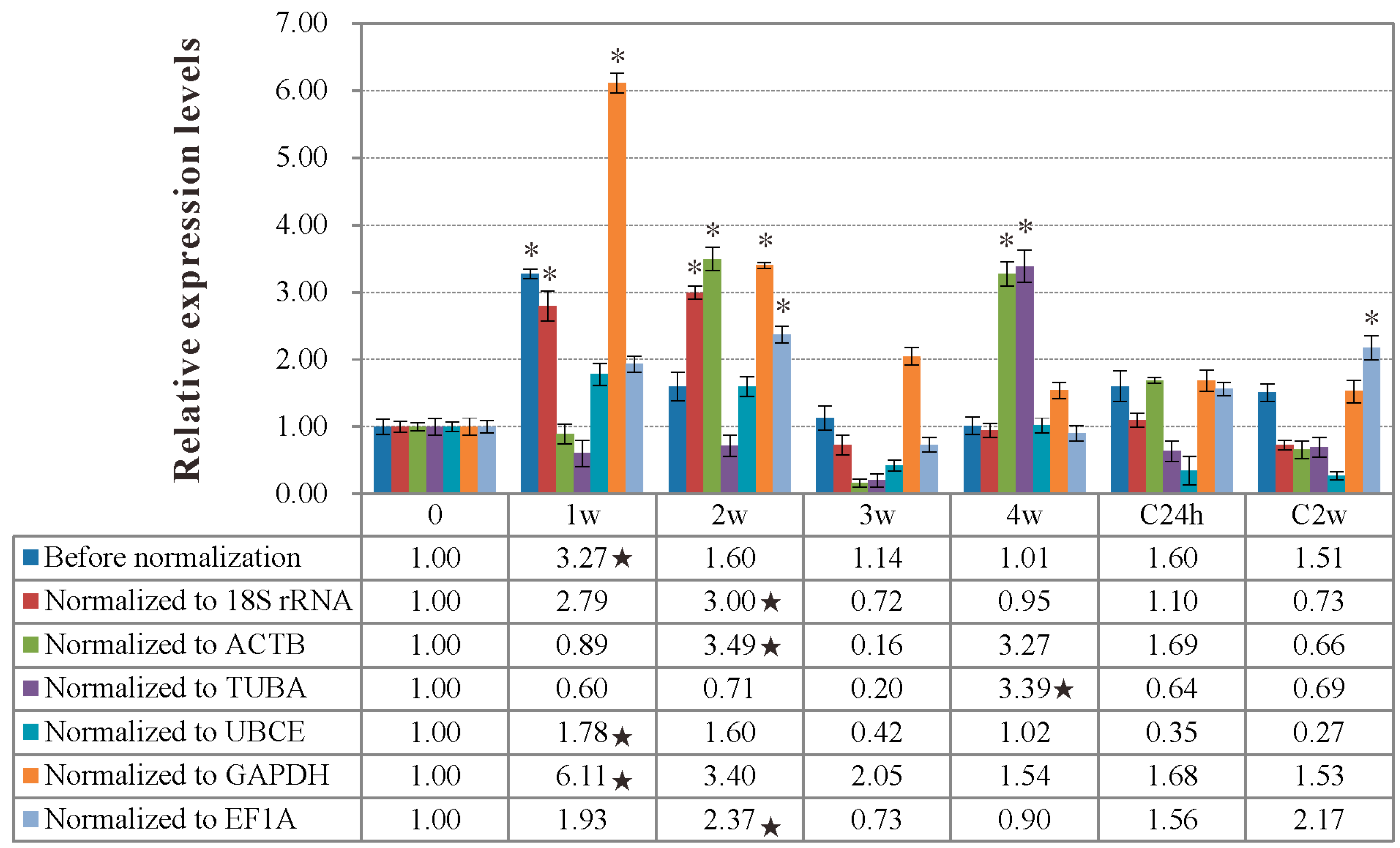
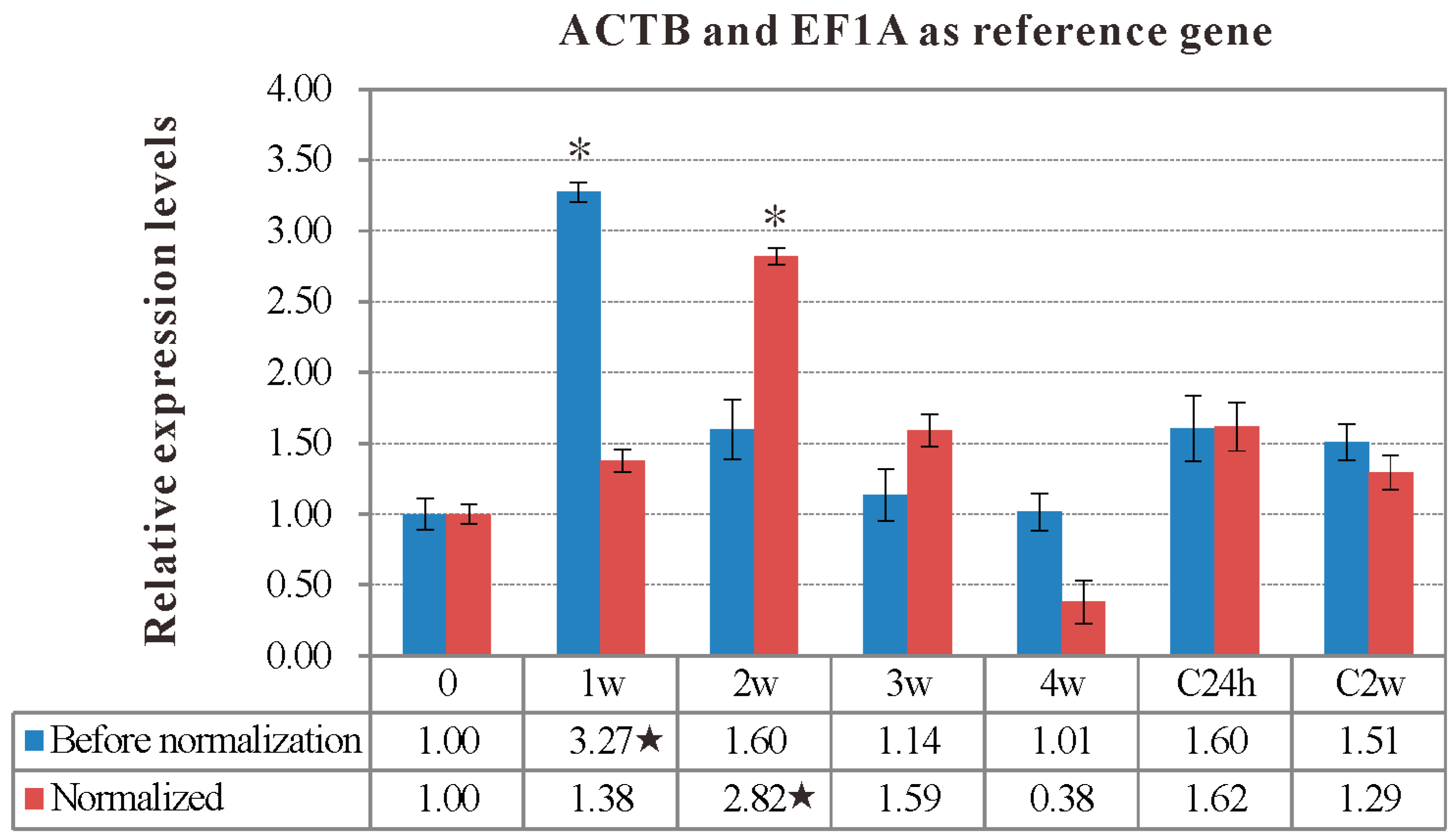
2.5. Validation of Reference Gene Selection


| Group | Time | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 1w | 2w | 3w | 4w | C24h | C2w | |
| rSip group | - | 1:16 | 1:64 | 1:32 | 1:8 | 1:32 | 1:8 |
| PBS group | - | - | - | - | - | 1:16 | - |
3. Discussion
4. Experimental Section
4.1. Animals and Sample Preparation
4.2. RNA Extraction and cDNA Synthesis
4.3. Primer Design
| Gene Symbol | Gene Name | Function | Accession Number/Reference |
|---|---|---|---|
| 18S rRNA | 18S ribosomal RNA | Ribosomal subunit | JF698683 |
| ACTB | β-actin | Cytoskeletal protein | XM_003443127 |
| TUBA | α-tubulin | Cytoskeletal protein | XM_003445344 |
| UBCE | Ubiquitin-conjugating enzyme | Protein degradation | XM_003460024 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | Glycolytic enzyme | XM_003460024 |
| EF1A | Elongation factor 1 α | Protein synthesis | AB075952 |
| IgM | Immunoglobulin M | Immunoglobulin | [44] |
4.4. Quantitative RT-PCR with SYBR Green
4.5. Validation of Reference Gene Selection
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wong, M.L.; Medrano, J.F. Real-time PCR for mRNA quantitation. Biotechniques 2005, 39, 75. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.; Hands, R.E.; Bustin, S.A. Quantification of mRNA using real-time RT-PCR. Nat. Protoc. 2006, 1, 1559–1582. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Dodd, A.; Lai, D.; McNabb, W.C.; Love, D.R. Validation of zebrafish (Danio rerio) reference genes for quantitative real-time RT-PCR normalization. Acta Biochim. Biophys. Sin. 2007, 39, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Ginzinger, D.G. Gene quantification using real-time quantitative PCR: An emerging technology hits the mainstream. Exp. Hematol. 2002, 30, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Gadaleta, A.; Giancaspro, A.; Cardone, M.F.; Blanco, A. Real-time PCR for the detection of precise transgene copy number in durum wheat. Cell. Mol. Biol. Lett. 2011, 16, 652–668. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.F.; Espy, M.J.; Binnicker, M.J. Detection of herpes simplex virus in cerebrospinal fluid using real-time PCR. In Molecular Diagnostics; Springer: New York, NY, USA, 2014; pp. 107–119. [Google Scholar]
- Grimholt, R.M.; Urdal, P.; Klingenberg, O.; Piehler, A.P. Rapid and reliable detection of α-globin copy number variations by quantitative real-time PCR. BMC Hematol. 2014, 14, 4. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Li, D.; Wang, W.; Su, M.; Lin, J.; Chen, H.; Jiang, L.; Chen, J.; Yang, B.; Ou, Q. Establishment of real time allele specific locked nucleic acid quantitative PCR for detection of HBV YIDD (ATT) mutation and evaluation of its application. PLoS ONE 2014, 9, e90029. [Google Scholar] [CrossRef] [PubMed]
- Collard, J.; Wang, X.; Mahamane, A.E.; Idi, I.; Issaka, B.; Ousseni, M.; Mayer, L.W.; Nato, F.; Moulia-Pelat, J. A five-year field assessment of rapid diagnostic tests for meningococcal meningitis in Niger by using the combination of conventional and real-time PCR assays as a gold standard. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, T.L.; McQualter, R.B. The quantitative real-time polymerase chain reaction for the analysis of plant gene expression. In Cereal Genomics; Springer: New York, NY, USA, 2014; pp. 97–115. [Google Scholar]
- Udvardi, M.K.; Czechowski, T.; Scheible, W. Eleven golden rules of quantitative RT-PCR. Plant Cell Online 2008, 20, 1736–1737. [Google Scholar] [CrossRef]
- Ye, X.; Zhang, L.; Dong, H.; Tian, Y.; Lao, H.; Bai, J.; Yu, L. Validation of reference genes of grass carp Ctenopharyngodon idellus for the normalization of quantitative real-time PCR. Biotechnol. Lett. 2010, 32, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Radonić, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yuan, M.; Shakeel, M.; Zhang, Y.; Wang, S.; Wang, X.; Zhan, S.; Kang, T.; Li, J. Selection and evaluation of reference genes for expression analysis using qRT-PCR in the beet armyworm Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae). PLoS ONE 2014, 9, e84730. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.G.; Wang, X.L.; Tian, J.; Liu, W.; Wu, F.; Jiang, M.; Wen, H. Evaluation of reference genes for quantitative real-time RT-PCR analysis of gene expression in Nile tilapia (Oreochromis niloticus). Gene 2013, 527, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Giricz, O.; Lauer-Fields, J.L.; Fields, G.B. The normalization of gene expression data in melanoma: Investigating the use of glyceraldehyde 3-phosphate dehydrogenase and 18S ribosomal RNA as internal reference genes for quantitative real-time PCR. Anal. Biochem. 2008, 380, 137–139. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S. EF1α is a useful internal reference for studies of gene expression regulation in amphioxus Branchiostoma japonicum. Fish Shellfish Immun. 2012, 32, 1068–1073. [Google Scholar] [CrossRef]
- Imai, T.; Ubi, B.E.; Saito, T.; Moriguchi, T. Evaluation of reference genes for accurate normalization of gene expression for real time-quantitative PCR in Pyrus pyrifolia using different tissue samples and seasonal conditions. PLoS ONE 2014, 9, e86492. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Yuan, F.; Long, G.; Qin, L.; Deng, Z. Selection of reference genes for quantitative real-time RT-PCR analysis in citrus. Mol. Biol. Rep. 2012, 39, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, L.; Mauriat, M.; Guénin, S.; Pelloux, J.; Lefebvre, J.F.; Louvet, R.; Rusterucci, C.; Moritz, T.; Guerineau, F.; Bellini, C. The lack of a systematic validation of reference genes: A serious pitfall undervalued in reverse transcription-polymerase chain reaction (RT-PCR) analysis in plants. Plant Biotechnol. J. 2008, 6, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Sun, L. Evaluation of housekeeping genes as references for quantitative real time RT-PCR analysis of gene expression in Japanese flounder (Paralichthys olivaceus). Fish Shellfish Immun. 2011, 30, 638–645. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, h34. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Sun, G.; Stiller, J.W.; Zhang, B. Genome-wide functional analysis of the cotton transcriptome by creating an integrated EST database. PLoS ONE 2011, 6, e26980. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.J.; Klesius, P.H.; Pasnik, D.J.; Bohnsack, J.F. Human Streptococcus agalactiae isolate in Nile tilapia (Oreochromis niloticus). Emerg. Infect. Dis. 2009, 15, 774. [Google Scholar] [CrossRef] [PubMed]
- Rioux, S.; Martin, D.; Ackermann, H.; Dumont, J.; Hamel, J.; Brodeur, B.R. Localization of surface immunogenic protein on group B streptococcus. Infect. Immun. 2001, 69, 5162–5165. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.; Yu, L.; Li, S.; Shen, X. Intranasal immunization with GBS surface protein Sip and ScpB induces specific mucosal and systemic immune responses in mice. FEMS Immunol. Med. Microbiol. 2010, 58, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.; Rioux, S.; Gagnon, E.; Boyer, M.; Hamel, J.; Charland, N.; Brodeur, B.R. Protection from group B streptococcal infection in neonatal mice by maternal immunization with recombinant Sip protein. Infect. Immun. 2002, 70, 4897–4901. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.Y.; Wang, K.Y.; Xiao, D.; Chen, D.F.; Geng, Y.; Wang, J.; He, Y.; Wang, E.L.; Huang, J.L.; Xiao, G.Y. Safety and immunogenicity of an oral DNA vaccine encoding Sip of Streptococcus agalactiae from Nile tilapia Oreochromis niloticus delivered by live attenuated Salmonella typhimurium. Fish Shellfish Immun. 2014, 38, 34–41. [Google Scholar] [CrossRef]
- He, Y.; Wang, K.Y.; Xiao, D.; Chen, D.F.; Huang, L.Y.; Liu, T.Q.; Wang, J.; Geng, Y.; Wang, E.L.; Yang, Q. A recombinant truncated surface immunogenic protein (tSip) plus adjuvant FIA confers active protection against Group B streptococcus infection in tilapia. Vaccine 2014, 32, 7025–7032. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, M.; Liu, C.; Qiu, R.; Sun, L. A divalent DNA vaccine based on Sia10 and OmpU induces cross protection against Streptococcus iniae and Vibrio anguillarum in Japanese flounder. Fish Shellfish Immun. 2012, 32, 1216–1222. [Google Scholar] [CrossRef]
- Hu, Y.; Sun, L. A bivalent Vibrio harveyi DNA vaccine induces strong protection in Japanese flounder (Paralichthys olivaceus). Vaccine 2011, 29, 4328–4333. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, C.; Sun, L. Comparative study of the immune effect of an Edwardsiella tarda antigen in two forms: Subunit vaccine vs. DNA vaccine. Vaccine 2011, 29, 2051–2057. [Google Scholar] [CrossRef] [PubMed]
- Racine, R.; Winslow, G.M. IgM in microbial infections: Taken for granted? Immunol. Lett. 2009, 125, 79–85. [Google Scholar] [CrossRef]
- Boes, M. Role of natural and immune IgM antibodies in immune responses. Mol. Immunol. 2000, 37, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Hu, Y.; Liu, C.; Sun, L. Construction and comparative study of monovalent and multivalent DNA vaccines against Streptococcus iniae. Fish Shellfish Immun. 2012, 33, 1303–1310. [Google Scholar] [CrossRef]
- Cruz, F.; Kalaoun, S.; Nobile, P.; Colombo, C.; Almeida, J.; Barros, L.M.; Romano, E.; Grossi-de-Sá, M.F.; Vaslin, M.; Alves-Ferreira, M. Evaluation of coffee reference genes for relative expression studies by quantitative real-time RT-PCR. Mol. Breed. 2009, 23, 607–616. [Google Scholar] [CrossRef]
- Filby, A.L.; Tyler, C.R. Appropriate housekeeping genes for use in expression profiling the effects of environmental estrogens in fish. BMC Mol. Biol. 2007, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Huggett, J.F.; Foy, C.A.; Benes, V.; Emslie, K.; Garson, J.A.; Haynes, R.; Hellemans, J.; Kubista, M.; Mueller, R.D.; Nolan, T. The digital MIQE guidelines: Minimum information for publication of quantitative digital PCR experiments. Clin. Chem. 2013, 59, 892–902. [Google Scholar] [CrossRef] [PubMed]
- Huggett, J.; Nolan, T.; Bustin, S.A. MIQE: Guidelines for the design and publication of a reliable real-time PCR assay. In Real-Time PCR: Advanced Technologies and Applications; Horizon Scientific Press: Wymondham, UK, 2013; pp. 247–258. [Google Scholar]
- Sun, Y.; Liu, C.; Sun, L. Construction and analysis of the immune effect of an Edwardsiella tarda DNA vaccine encoding a D15-like surface antigen. Fish Shellfish Immun. 2011, 30, 273–279. [Google Scholar] [CrossRef]
- Phuyindee, C.; Unajak, S.; Srisapoome, P. Molecular characterization and expression analysis of a cDNA encoding immunoglobulin (Ig) M heavy chain secreted form of Nile tilapia (Oreochromis niloticus). In Proceedings of the 49th Kasetsart University Annual Conference, Kasetsart University, Thailand, 1–4 February 2011; Volume 3, pp. 200–211.
- Kubista, M.; Andrade, J.M.; Bengtsson, M.; Forootan, A.; Jonák, J.; Lind, K.; Sindelka, R.; Sjöback, R.; Sjögreen, B.; Strömbom, L. The real-time polymerase chain reaction. Mol. Asp. Med. 2006, 27, 95–125. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, E.; Wang, K.; Chen, D.; Wang, J.; He, Y.; Long, B.; Yang, L.; Yang, Q.; Geng, Y.; Huang, X.; et al. Evaluation and Selection of Appropriate Reference Genes for Real-Time Quantitative PCR Analysis of Gene Expression in Nile Tilapia (Oreochromis niloticus) during Vaccination and Infection. Int. J. Mol. Sci. 2015, 16, 9998-10015. https://doi.org/10.3390/ijms16059998
Wang E, Wang K, Chen D, Wang J, He Y, Long B, Yang L, Yang Q, Geng Y, Huang X, et al. Evaluation and Selection of Appropriate Reference Genes for Real-Time Quantitative PCR Analysis of Gene Expression in Nile Tilapia (Oreochromis niloticus) during Vaccination and Infection. International Journal of Molecular Sciences. 2015; 16(5):9998-10015. https://doi.org/10.3390/ijms16059998
Chicago/Turabian StyleWang, Erlong, Kaiyu Wang, Defang Chen, Jun Wang, Yang He, Bo Long, Lei Yang, Qian Yang, Yi Geng, Xiaoli Huang, and et al. 2015. "Evaluation and Selection of Appropriate Reference Genes for Real-Time Quantitative PCR Analysis of Gene Expression in Nile Tilapia (Oreochromis niloticus) during Vaccination and Infection" International Journal of Molecular Sciences 16, no. 5: 9998-10015. https://doi.org/10.3390/ijms16059998
APA StyleWang, E., Wang, K., Chen, D., Wang, J., He, Y., Long, B., Yang, L., Yang, Q., Geng, Y., Huang, X., Ouyang, P., & Lai, W. (2015). Evaluation and Selection of Appropriate Reference Genes for Real-Time Quantitative PCR Analysis of Gene Expression in Nile Tilapia (Oreochromis niloticus) during Vaccination and Infection. International Journal of Molecular Sciences, 16(5), 9998-10015. https://doi.org/10.3390/ijms16059998