
Subchronic Exposure to Arsenic Represses the TH/TRβ1-CaMK IV Signaling Pathway in Mouse Cerebellum

Abstract
:1. Introduction
2. Results
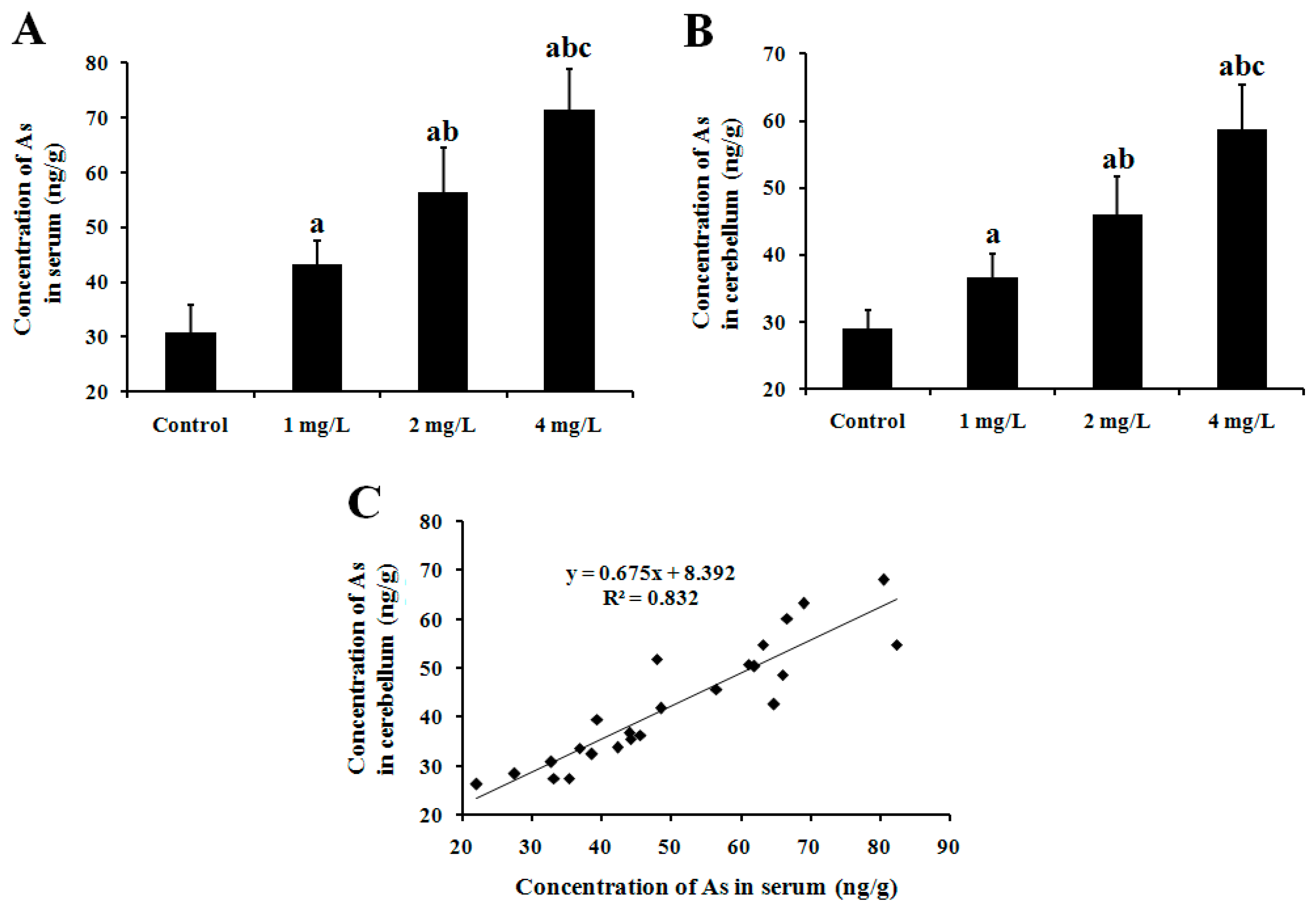
2.1. As Concentration in Mouse Serum and Cerebellum

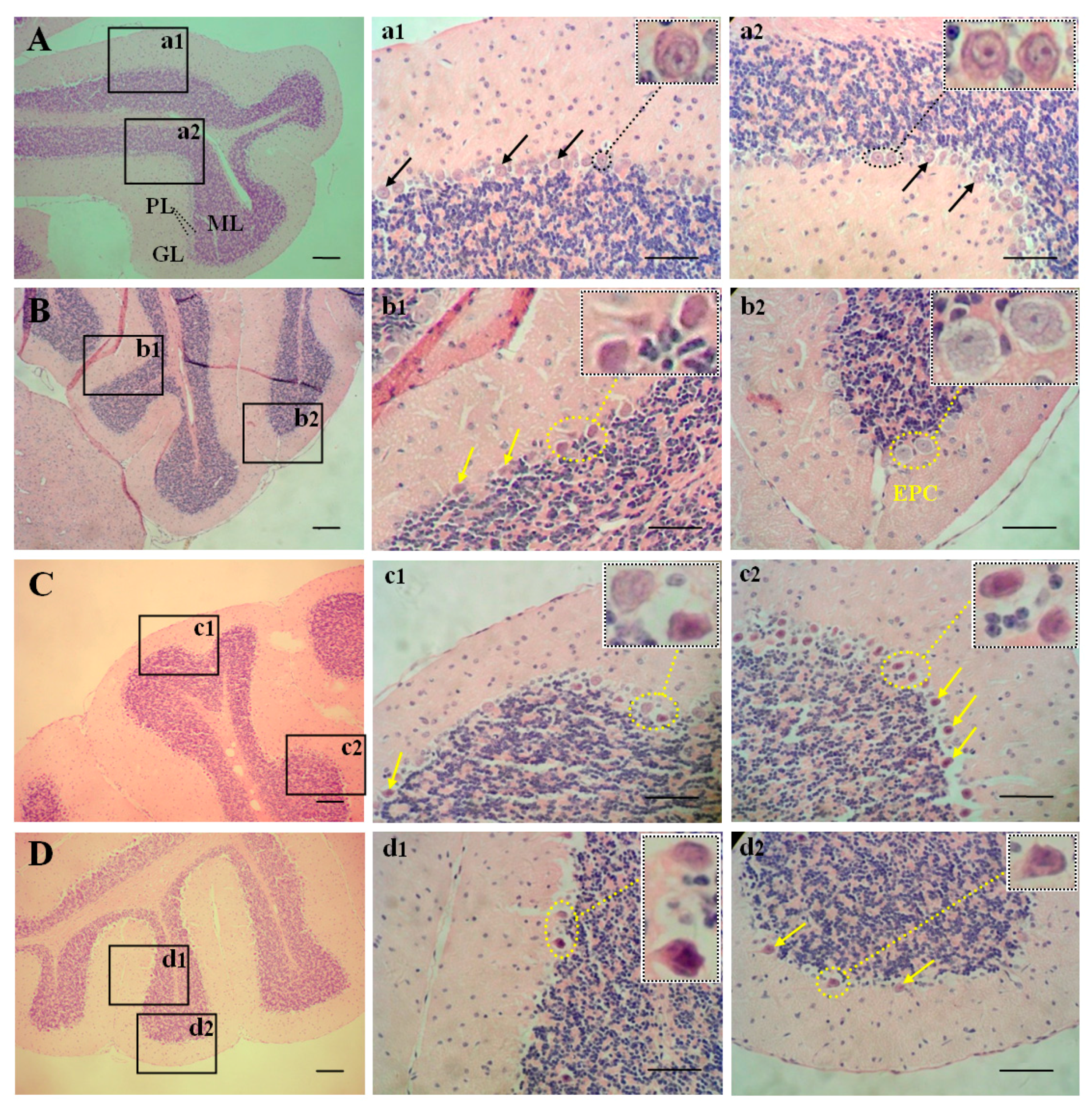
2.2. Morphology of the Cerebellum

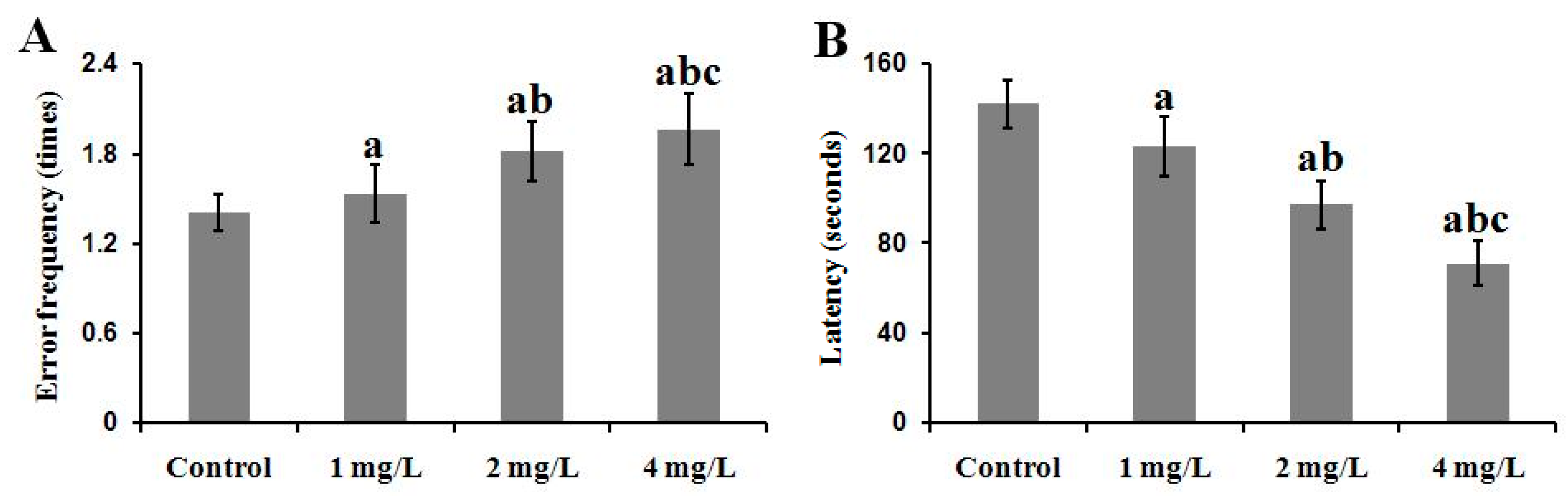
2.3. Performance of the Behavioral Tests
2.3.1. The Step-Down Passive Avoidance Task

2.3.2. The MWM Tests

2.4. Protein Expression of CaMK IV in Mouse Cerebellum
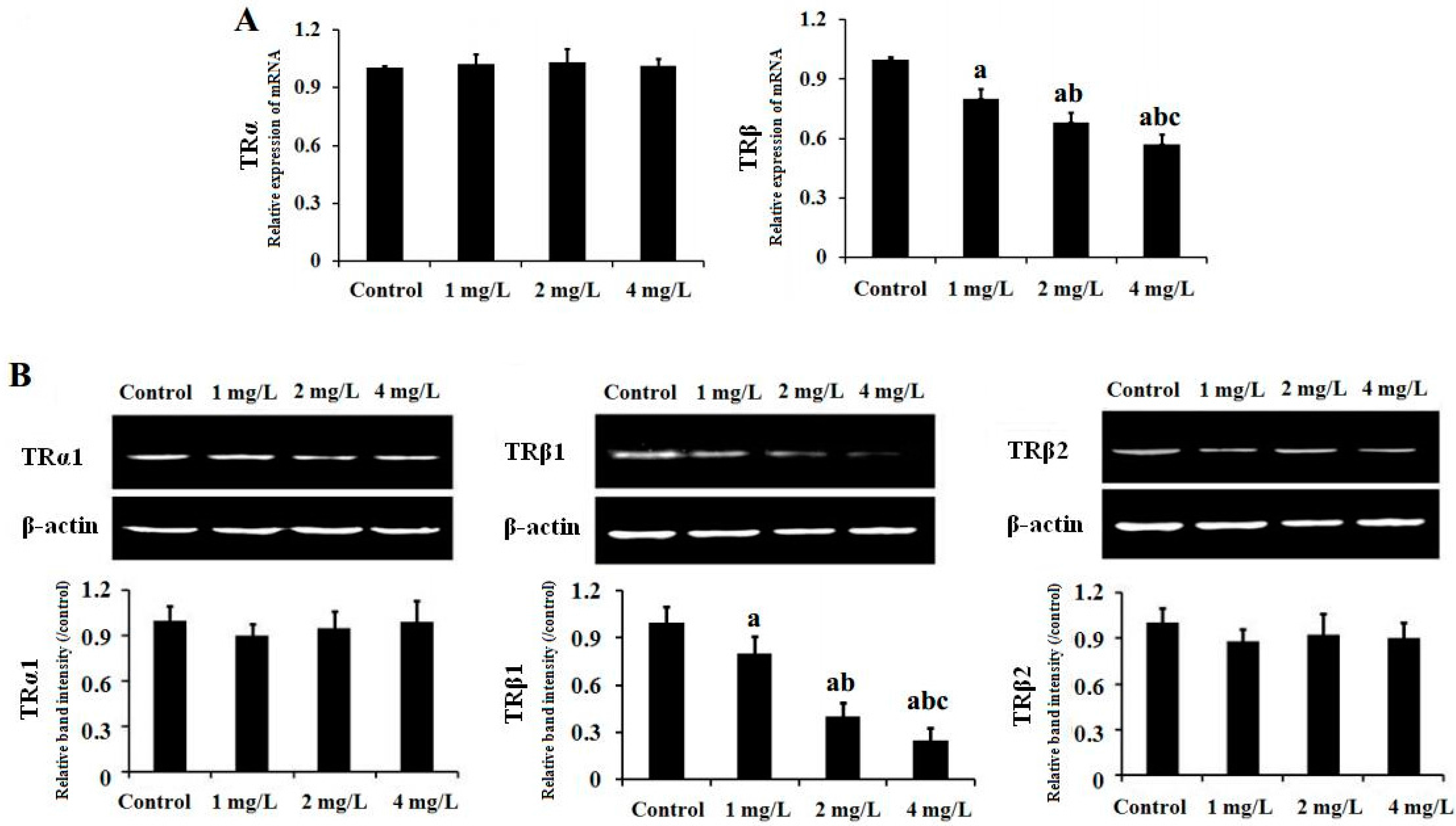
2.5. Gene and Protein Expression of TR in Mouse Cerebellum


2.6. Gene and Protein Expression of RXR in Mouse Cerebellum

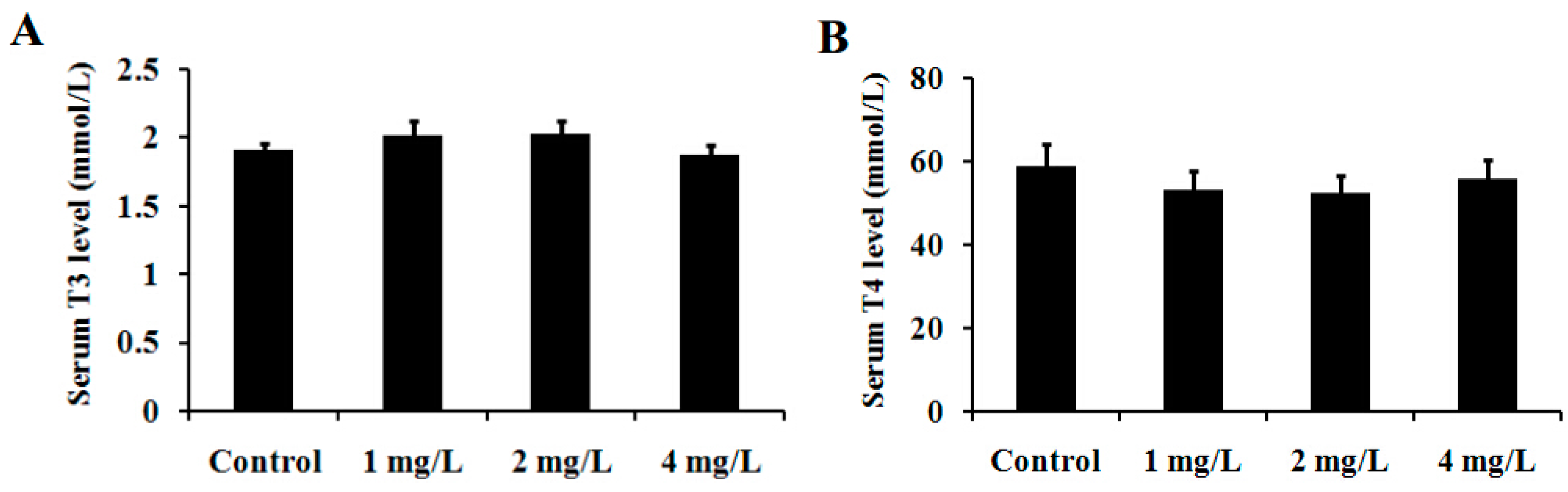
2.7. T3 and T4 Levels in Serum

3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals and Treatment
4.3. Detection of As Concentration in Mouse Serum and Cerebellum Tissue
4.4. Behavioral Testing
4.4.1. Step-down Passive Avoidance Task
4.4.2. MWM Tests
4.5. Hematoxylin-Eosin (HE) Staining
4.6. Real Time RT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| RXRα | TGAGACATACGTGGAGGCAAACA GAAGAGCCCTTATGATCCCAAAC | GGCCCACTCCACAAGAGTGAA AGATAGCGAAGTCCCGTCCC |
| RXRβ | GCAAACGGCTCTGTGCAATC ATGGCATTCCGGGCGATTT | GCTGGCGCTTGTCCACTGTA GGTTTTGGTTTCTCGAAGCTCA |
| TRα | GACAAGGCCACCGGTTATCACTAC GACAAATGTGGCGGGACCATA | CAGCAGCTGTCATACTTGCAG GA TGGATTAGCCATTCACACTTCTC |
| TRβ | AGCCAGAACCCACGGATGAG AGGACACGAAGTGAGAAGCC | CGATGGGTGCTTGTCCAATG GTGAGGGTTGAAGTTGAGAACA |
| β-actin GAPDH | CATCCGTAAAGACCTCTATGCCAAC AGGTCGGTGTGAACGGATTTG | ATGGAGCCACCGATCCACA TGTAGACCATGTAGTTGAGGTCA |
4.7. Western Blot
4.8. RIA
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chen, K.P.; Wu, H.Y.; Wu, T.C. Epidemiologic studies on blackfoot disease in Taiwan. 3. Physicochemical characteristics of drinking water in endemic Blackfoot disease areas. Mem. Coll. Med. 1962, 8, 115–129. [Google Scholar]
- Chowdhury, U.K.; Biswas, B.K.; Chowdhury, T.R.; Samanta, G.; Mandal, B.K.; Basu, G.C.; Chanda, C.R.; Lodh, D.; Saha, K.C.; Mukherjee, S.K.; et al. Groundwater arsenic contamination in Bangladesh and West Bengal, India. Environ. Health Perspect. 2000, 108, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Tondel, M.; Ahmad, S.A.; Axelson, O. Diabetes mellitus associated with arsenic exposure in Bangladesh. Am. J. Epidemiol. 1998, 148, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Bian, J.M.; Li, Z.Y.; Wang, N.; Zhang, N.; Zhang, H. Relationship between hydrochemical enviornment and arsenism in areas with arsenic poisoning drinking water in China. Asian J. Ecotoxicol. 2013, 8, 8. [Google Scholar]
- Hsieh, R.L.; Huang, Y.L.; Shiue, H.S.; Huang, S.R.; Lin, M.I.; Mu, S.C.; Chung, C.J.; Hsueh, Y.M. Arsenic methylation capacity and developmental delay in preschool children in Taiwan. Int. J. Hyg. Environ. Health 2014, 217, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Nahar, M.N.; Inaoka, T.; Fujimura, M. A consecutive study on arsenic exposure and intelligence quotient (IQ) of children in Bangladesh. Environ. Health Prev. Med. 2014, 19, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Baldissarelli, L.A.; Capiotti, K.M.; Bogo, M.R.; Ghisleni, G.; Bonan, C.D. Arsenic alters behavioral parameters and brain ectonucleotidases activities in zebrafish (Danio rerio). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2012, 155, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.; Zheng, G.; Liu, M.; Shen, X.; Zhao, F.; Wang, J.; Zhang, J.; Huang, G.; Dai, P.; Chen, Y.; et al. Changes in the synaptic structure of hippocampal neurons and impairment of spatial memory in a rat model caused by chronic arsenite exposure. Neurotoxicology 2012, 33, 1230–1238. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, S.; Piao, F.; Hong, Y.; Liu, P.; Zhao, Y. Arsenic down-regulates the expression of CaMK IV, an important gene related to cerebellar LTD in mice. Neurotoxicol. Teratol. 2009, 31, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Su, Q.; Jiang, M.; Xie, H.; Cong, J.; Wang, L. Arsenic affects on cerebellar development of mice. Toxicol. Mech. Methods 2013, 23, 672–677. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gao, Y.; Yao, H.; Zhou, L.; Sun, D.; Wang, J. Neuroglobin involvement in the course of arsenic toxicity in rat cerebellar granule neurons. Biol. Trace Elem. Res. 2013, 155, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Piao, F.; Ma, N.; Hiraku, Y.; Murata, M.; Oikawa, S.; Cheng, F.; Zhong, L.; Yamauchi, T.; Kawanishi, S.; Yokoyama, K. Oxidative DNA damage in relation to neurotoxicity in the brain of mice exposed to arsenic at environmentally relevant levels. J. Occup. Health 2005, 47, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Mizoi, M.; An, Y.; Nakano, M.; Wanibuchi, H.; Endo, G.; Endo, Y.; Hoshino, M.; Okada, S.; Yamanaka, K. Oral administration of diphenylarsinic acid, a degradation product of chemical warfare agents, induces oxidative and nitrosative stress in cerebellar Purkinje cells. Life Sci. 2007, 81, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Negishi, T.; Takahashi, M.; Matsunaga, Y.; Hirano, S.; Tashiro, T. Diphenylarsinic acid increased the synthesis and release of neuroactive and vasoactive peptides in rat cerebellar astrocytes. J. Neuropathol. Exp. Neurol. 2012, 71, 468–479. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Funabiki, K.; Nakanishi, S. Role of granule-cell transmission in memory trace of cerebellum-dependent optokinetic motor learning. Proc. Natl. Acad. Sci. USA 2014, 111, 5373–5378. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Ginty, D.D.; Linden, D.J. A late phase of cerebellar long-term depression requires activation of CaMK IV and CREB. Neuron 1999, 23, 559–568. [Google Scholar] [CrossRef]
- Casu, M.A.; Pisu, C.; Sanna, A.; Tambaro, S.; Spada, G.P.; Mongeau, R.; Pani, L. Effect of Δ9-tetrahydrocannabinol on phosphorylated CREB in rat cerellum: An imminohistochemical study. Brain Res. 2005, 1048, 41–47. [Google Scholar] [CrossRef] [PubMed]
- El Hajj Chehadeh, S.; Dreumont, N.; Willekens, J.; Canabady-Rochelle, L.; Jeannesson, E.; Alberto, J.M.; Daval, J.L.; Guéant, J.L.; Leininger-Muller, B. Early methyl donor deficiency alters cAMP signaling pathway and neurosteroidogenesis in the cerebellum of female rat pups. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E1009–E1019. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Chatila, T.A.; Ram, R.A.; Thompson, R.F. Impaired memory of eyeblink conditioning in CaMKIVKO mice. Behav. Neurosci. 2009, 123, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Boyden, E.S.; Katoh, A.; Pyle, J.L.; Chatila, T.A.; Tsien, R.W.; Raymond, J.L. Selective engagement of plasticity mechanisms for motor memorystorage. Neuron 2006, 51, 823–834. [Google Scholar] [CrossRef] [PubMed]
- Ho, N.; Liauw, J.A.; Blaeser, F.; Wei, F.; Hanissian, S.; Muglia, L.M.; Wozniak, D.F.; Nardi, A.; Arvin, K.L.; Holtzman, D.M.; et al. Impaired synaptic plasticity and cAMP response element-binding protein activation in Ca2+/calmodulin-dependent protein kinase type IV/Gr-deficient mice. J. Neurosci. 2000, 20, 6459–6572. [Google Scholar] [PubMed]
- Li, D.; Yamada, T.; Wang, F.; Vulin, A.I.; Samuels, H.H. Novel roles of retinoid X receptor (RXR) and RXR ligand in dynamicallymodulating the activity of the thyroid hormone receptor/RXR heterodimer. J. Biol. Chem. 2004, 279, 7427–7437. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Brent, G.A. Thyroidhormone-dependent gene expression in differentiated embryonic stem cells and embryonal carcinoma cells: Identificationof novel thyroid hormone target genes by deoxyribonucleic acid microarray analysis. Endocrinology 2005, 146, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Morte, B.; Díez, D.; Ausó, E.; Belinchón, M.M.; Gil-Ibáñez, P.; Grijota-Martínez, C.; Navarro, D.; de Escobar, G.M.; Berbel, P.; Bernal, J. Thyroid hormone regulation of gene expression in the developing rat fetal cerebral cortex: Prominent role of the Ca2+/calmodulin-dependent protein kinase IV pathway. Endocrinology 2010, 151, 810–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, J.C.; Nomikos, A.P.; Wungjiranirun, M.; Sherman, J.R.; Ingram, L.; Batki, C.; Lariviere, J.P.; Hamilton, J.W. Arsenic as an endocrine disruptor: Arsenic disrupts retinoic acid receptor-and thyroid hormone receptor-mediated gene regulation and thyroid hormone-mediated amphibian tail metamorphosis. Environ. Health Perspect. 2008, 116, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, V.M.; Jiménez-Capdeville, M.E.; Giordano, M. The effects of arsenic exposure on the nervous system. Toxicol. Lett. 2003, 145, 1–18. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Chou, H.Y.; The, H.W.; Chen, C.M.; Chen, C.J. The effects of chronic arsenic exposure from drinking water on the neurobehavioral development in adolescence. Neurotoxicology 2003, 24, 747–753. [Google Scholar] [CrossRef]
- Nagaraja, T.N.; Desiraju, T. Effects on operant learning and brain acetylcholine esterase activity in rats following chronic inorganic arsenic intake. Hum. Exp. Toxicol. 1994, 13, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, V.M.; Carrizales, L.; Mendoza, M.S.; Fajardo, O.R.; Giordano, M. Effects of sodium arsenite exposure on development and behavior in the rat. Neurotoxicol. Teratol. 2002, 24, 743–750. [Google Scholar] [CrossRef]
- Rodríguez, V.M.; Carrizales, L.; Jiménez-Capdeville, M.E.; Dufour, L.; Giordano, M. The effects of sodium arsenite exposure on behavioral parameters in the rat. Brain Res. Bull. 2001, 55, 301–308. [Google Scholar] [CrossRef]
- Schmahmann, J.D. The role of the cerebellum in cognition and emotion: Personal reflections since 1982 on the dysmetria of thought hypothesis, and its historical evolution from theory to therapy. Neuropsychol. Rev. 2010, 20, 236–260. [Google Scholar] [CrossRef] [PubMed]
- Bastian, A.J. Moving, sensing and learning with cerebellar damage. Curr. Opin. Neurobiol. 2011, 21, 596–601. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, E.; Casali, S. Seeking a unified framework for cerebellar function and dysfunction: From circuit operations to cognition. Front. Neural Circ. 2012, 6, 116. [Google Scholar] [CrossRef] [PubMed]
- Krebs, J.; Honegger, P. Calmodulin kinase IV: Expression and function during rat brain development. Biochim. Biophys. Acta 1996, 1313, 217–222. [Google Scholar] [CrossRef]
- Krebs, J.; Means, R.L.; Honegger, P. Induction of calmodulin kinase IV by the thyroid hormone during the development of rat brain. J. Biol. Chem. 1996, 271, 11055–11105. [Google Scholar] [PubMed]
- Mark, M.; Ghyselinck, N.B.; Chambon, P. Function of retinoid nuclear receptors: Lessons from genetic and pharmacological dissections of the retinoic acid signaling pathway during mouse embryogenesis. Annu. Rev. Pharmacol. Toxicol. 2006, 46, 451–480. [Google Scholar] [CrossRef] [PubMed]
- Suen, C.S.; Yen, P.M.; Chin, W.W. In vitro transcriptional studies of the roles of the thyroidhormone (T3) response elements and minimal promoters in T3-stimulated gene transcription. J. Biol. Chem. 1994, 269, 1314–1322. [Google Scholar] [PubMed]
- Mohanta, R.K.; Garg, A.K.; Dass, R.S.; Behera, S.K. Blood biochemistry, thyroid hormones, and oxidant/antioxidant status of guinea pigs challenged with sodium arsenite or arsenic trioxide. Biol. Trace Elem. Res. 2014, 160, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.J.; Li, H.B.; Xiang, P.; Zhang, X.; Ma, L.Q. Short-term exposure of arsenite disrupted thyroid endocrine system and altered gene transcription in the HPT axis in zebrafish. Environ. Pollut. 2015, 205, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Van Beeren, H.C.; Kwakkel, J.; Ackermans, M.T.; Wiersinga, W.M.; Fliers, E.; Boelen, A. Action of specific thyroidhormone receptor α(1) and β(1) antagonists in the central and peripheral regulation of thyroidhormone metabolism in the rat. Thyroid 2012, 22, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Husson, M.; Enderlin, V.; Alfos, S.; Féart, C.; Higueret, P.; Pallet, V. Triiodothyronine administration reverses vitamin A deficiency-related hypo-expression of retinoic acid and triiodothyronine nuclear receptors and of neurogranin in rat brain. Br. J. Nutr. 2003, 90, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Husson, M.; Enderlin, V.; Alfos, S.; Boucheron, C.; Pallet, V.; Higueret, P. Expression of neurogranin and neuromodulin is affected in the striatum of vitamin A-deprived rats. Brain Res. Mol. Brain Res. 2004, 123, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, J.; Zhao, L.; Hu, S.; Piao, F. Effect of subchronic exposure to arsenic on levels of essential trace elements in mice brain and its gender difference. Biometals 2013, 26, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.H.; Zhang, J.; Wang, Y.M.; Ye, Y.P.; Luo, Q.Q. Perinatal exposure to bisphenol-A impairs learning-memory by concomitant down-regulation of N-methyl-d-aspartate receptors of hippocampus in male offspring mice. Horm. Behav. 2010, 58, 326–333. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, H.; Li, S.; Guo, Y.; Liu, X.; Yang, Y.; Guo, J.; Li, S.; Zhang, C.; Shang, L.; Piao, F. Subchronic Exposure to Arsenic Represses the TH/TRβ1-CaMK IV Signaling Pathway in Mouse Cerebellum. Int. J. Mol. Sci. 2016, 17, 157. https://doi.org/10.3390/ijms17020157
Guan H, Li S, Guo Y, Liu X, Yang Y, Guo J, Li S, Zhang C, Shang L, Piao F. Subchronic Exposure to Arsenic Represses the TH/TRβ1-CaMK IV Signaling Pathway in Mouse Cerebellum. International Journal of Molecular Sciences. 2016; 17(2):157. https://doi.org/10.3390/ijms17020157
Chicago/Turabian StyleGuan, Huai, Shuangyue Li, Yanjie Guo, Xiaofeng Liu, Yi Yang, Jinqiu Guo, Sheng Li, Cong Zhang, Lixin Shang, and Fengyuan Piao. 2016. "Subchronic Exposure to Arsenic Represses the TH/TRβ1-CaMK IV Signaling Pathway in Mouse Cerebellum" International Journal of Molecular Sciences 17, no. 2: 157. https://doi.org/10.3390/ijms17020157
APA StyleGuan, H., Li, S., Guo, Y., Liu, X., Yang, Y., Guo, J., Li, S., Zhang, C., Shang, L., & Piao, F. (2016). Subchronic Exposure to Arsenic Represses the TH/TRβ1-CaMK IV Signaling Pathway in Mouse Cerebellum. International Journal of Molecular Sciences, 17(2), 157. https://doi.org/10.3390/ijms17020157